
- Жгутики бактерий. Строение, химический состав, расположение. Методы выявления. Фимбрии и F – пили.
- Жгутики строение расположение функция способы выявления
- Жгутики — это поверхностные локомоторные структуры. Строение жгутиков прокариот и эукариот.
- Общая характеристика жгутиков
- Особенности жгутиков прокариот и эукариот
- Жгутики архей и бактерий
- Строение и функционирование бактериального жгутика
- Структура базального мотора
- Жгутик ядерной клетки
Жгутики бактерий. Строение, химический состав, расположение. Методы выявления. Фимбрии и F – пили.
Жгутик — спирально изогнутая полая нить, образованная субъединицами флагеллина, поверхностная структура, присутствующая у многих прокариотических и эукариотических клеток и служащая для их движения в жидкой среде или по поверхности твёрдых сред. бактериальный жгутик имеет толщину 10—20 нм и длину 3—15 мкм, он пассивно вращается расположенным в мембране мотором
Жгутики бактерий состоят из трёх субструктур:
• Филамент (фибрилла, пропеллер) — полая белковая нить толщиной 10—20 нм и длиной 3—15 мкм, состоящая из флагеллина, субъединицы которого уложены по спирали. Полость внутри используется при синтезе жгутика — он происходит в направлении от цитоплазматической мембраны. По полости к собираемому в настоящий момент участку переносятся субъединицы флагеллина.
• Крюк — более толстое, чем филамент (20—45 нм), белковое (не флагеллиновое) образование.
• Базальное тело (трансмембранный мотор)
Расположение жгутиков — характерный признак, имеющий таксономическое значение. Варианты расположения жгутиков приведены на рис. 4-1. У некоторых бактерий жгутики расположены по всей поверхности клеточной стенки (например, у бактерий рода Proteus), такие бактерии известны как перитрихи . Некоторые бактерии снабжены только одним толстым жгутиком (например, представители рода Vibrio), они известны как монотрихи. Политрихи — бактерии, имеющие одиночный по виду жгутик, образованный пучком из 2-50 жгутиков. Полярные жгутики прикреплены к одному или обоим концам бактерии. Монополярно-политрихиальное расположение жгутиков имеют лофотрихи , к ним, например, относят представителей рода Pseudomonas. Биполярно-политрихиальное жгутикование имеют амфитрихи (например, бактерии рода Spirillum).
Окраска жгутиков методом Леффлера.
В основе выявления жгутиков лежит осаждение на них красителя, чем достигается увеличение толщины жгутиков и уменьшение их прозрачности.
• Препарат готовят из 16-18 часовой культуры, которую вносят в 1-2 мл стерильной водопроводной воды до получения тонкой опалесцирующей взвеси.
• Через 20 мин капля суспензии наносят на поверхность чистого обезжиренного стекла и высушивают на воздухе.
• Обрабатывают в течение 15 мин протравой следующего состава: 1 мл насыщенного спиртового раствора основного фуксина, 10 мл 25% водного раствора таннина, 5 мл насыщенного водного раствора сернокислого железа.
• Препарат промывают водой.
• Окрашивают карболовым фуксином Циля, разведенным водой в соотношении 1:1, в течение 5 мин при легком подогревании.
• Промывают водой, высушивают.
При микроскопии готового препарата жгутики видны как тонкие нитевидные структуры.
Окраска по Романовскому — Гимзе
цитологический метод окраски простейших, бактерий, клеточных структур и тканей различных видов (в том числе крови) при световой микроскопии. Предложена в 1904 году Густавом Гимзой. В авторской версии название красителя — «Giemsasche Lözung für die Romanowsky färbung» (Раствор Гимзы для окраски по Романовскому) Окрашивает ацидофильные образования в различные оттенки красного цвета, базофильные — в цвета от пурпурного до синего.
Методика окраски. Мазки, фиксированные в метиловом спирте, окрашивают раствором (1 мл готовой жидкой краски + 2 мл основного буферного раствора + 47 мл дистиллированной воды) в течение 40—120 мин (продолжительность окрашивания подбирают эмпирически). Пользуются фосфатным буфером, но рН буфера зависит от вида мазка: для мазка костного мозга — 5,8 — 6,0, для мазка крови — 6,4 — 6,5, для выявления простейших — 6,8, малярийного плазмодия — 7,0 — 7,2. Ополаскивают в дистиллированной воде, высушивают и исследуют при иммерсии. Бактерии окрашиваются в фиолетово-красный цвет, цитоплазма клеток — в голубой, ядра — в красный. При окрашивании простейших их цитоплазма приобретает голубой цвет, а ядра — красно-фиолетовый.
Фимбрии (от лат. firnbriac – бахрома), длинные, тонкие, прямые выросты, состоящие из гидрофобного белка и находящиеся в большом количестве (иногда до нескольких тысяч) на поверхности клеток грамотрицательных бактерий. Длина Ф. – до 12 мкм, толщина – не более 100 Å. Они значительно тоньше и короче жгутиков. «Мужские» клетки бактерий (доноры) могут иметь 1–3 половые Ф. (пили), образующие между ними и «женскими» клетками (реципиентами) полые мостики, через которые при конъюгации бактерий передаётся ДНК. Ф. могут быть как у подвижных, так и у неподвижных бактерий; возникают обычно из базального тельца, находящегося в цитоплазматической мембране, и проходят через клеточную стенку наружу. Ф. придают бактериальной клетке способность неспецифически «прилипать» к плотной поверхности клеток, тканей и т.п.
F-пили бактерий , или «секс-пили», — жёсткие цилиндрические образования, участвующие в конъюгации бактерий. Пили впервые обнаружены у Escherichia coli K12, то есть у штаммов, содержащих F-фактор ( (англ. fertility плодовитость; син.: половой фактор бактерий, секс-фактор) — плазмида, определяющая конъюгационные свойства мужских штаммов бактерий.). Обычно клетка снабжена 1-2 пилями, имеющими вид полых белковых трубочек длиной 0,5-10 мкм; нередко они имеют шаровидное утолщение на конце. Большинство F-пилей образует специфический белок — пилин. Образование пилей кодируют плазмиды. Их идентифицируют с помощью донорспецифических бактериофагов, адсорбирующихся на пилях и лизирующих клетки.
Поможем написать любую работу на аналогичную тему
Жгутики бактерий. Строение, химический состав, расположение. Методы выявления. Фимбрии и F – пили.
Жгутики бактерий. Строение, химический состав, расположение. Методы выявления. Фимбрии и F – пили.
Жгутики бактерий. Строение, химический состав, расположение. Методы выявления. Фимбрии и F – пили.
Источник
Жгутики строение расположение функция способы выявления
Схема строения жгутика бактерий.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от ЦПМ, имеющие большую длину, чем сама клетка. Толщины жгутиков 12-20 нм, длина 3-12 мкм.
Жгутики состоят из сократительного белка типа миозина – флагеллина (от. лат. flagellum – жгутик), обладающего антигенной специфичностью. Субъединицы флагеллина закручены в виде спирали.


Жгутик состоит из трех компонентов – спиральной жгутиковой нити (филамента) постоянной толщины, крючка (колена) и базального тельца. Крючок, к которому присоединена жгутиковая нить, имеет длину 30-45 нм и состоит из отличающегося от флагеллина белка. Он соединен с базальным тельцем, которое располагается в оболочке (в клеточной стенке и ЦПМ). Нити жгутиков приводит в движение мембранный шарнироподобный базальный крюк. Базальное тельце состоит из центрального стержня, заключенного в систему особых колец. Кольца выполняют роль «приводного диска» и «подшипника» на внутренней поверхности пептидогликанового слоя. Вся конструкция выполняет функцию хемомеханического преобразователя (флагеллиновый мотор).
У грамотрицательных бактерий две пары колец: внешняя (кольца L и Р) и внутренняя (кольца S и М). Кольца L и Р расположены внутри клеточной стенки (кольцо L в ЛПС, а кольцо Р – в слое пептидогликана). Они выполняют, очевидно, роль втулки для стержня. Внутренняя пара (кольца S и М) фиксирована на ЦПМ, причем кольцо S располагается в периплазматическом пространстве, а кольцо М – на ЦПМ или в ней. Жгутики грамположительных бактерий, имеющих более толстую гомогенную клеточную стенку, содержат только одну пару колец – S и М.
Вращение жгутика в клеточной стенке происходит из-за вращательного движения колец S и М относительно друг друга и обеспечивается за счет энергии трансмембранного градиента ионов водорода или натрия. Благодаря такому вращению происходит движение бактерий в наиболее благоприятном для них направлении. Жгутиковый аппарат обладает особым бинарным переключателем, который позволяет менять направление вращения жгутиков против часовой стрелки на противоположное. Таким образом, бактерии, получив химический сигнал из окружающей среды, изменяют направление движения и выбирают оптимальные условия обитания.
Модель движения жгутика.

Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном конце клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Расположение жгутиков у бактерий.

Жгутики выявляют при помощи электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки специальными методами, основанными на протравливании и адсорбции различных веществ, приводящих к увеличению толщины жгутиков (например, после серебрения). Подвижность бактерий определяют при помощи фазово-контрастной или световой микроскопии «раздавленной» или «висячей» капли).

Vibrio cholerae. Фрагменты клеток с полярно расположенным жгутиком (Ж). Электронная микроскопия, х130000.
Источник
Жгутики — это поверхностные локомоторные структуры. Строение жгутиков прокариот и эукариот.

Жгутики – это длинные нитевидные образования на поверхности клетки, обеспечивающие ее активное пространственное перемещение. Несмотря на многообразие организмов, эти структуры внутри каждого надцарства (прокариот либо эукариот) характеризуются общей схемой строения.
Общая характеристика жгутиков
У доядерных организмов (бактерий и архей) жгутики – это основной способ передвижения. Среди эукариот эти локомоторные структуры в основном присутствуют у одноклеточных организмов, – простейших, но также характерны для гамет растений и животных. У некоторых многоклеточных беспозвоночных, – например, губок, – жгутики выполняют функцию перемещения жидкого субстрата относительно неподвижного клеточного пласта.

Морфологически жгутик состоит из закрепленного в толще клетки основания и длинной наружной нити, совершающей вращательные движения по спиральной траектории. Строение и механизм работы этих частей у прокариот и эукариот сильно отличаются, в связи с чем выделяют два соответствующих класса жгутиков.
Особенности жгутиков прокариот и эукариот
Наружную нить жгутика называют филаментом. У прокариот она состоит из белка флагеллина и пассивно движется за счет вращения базального мотора. Филамент ядерных клеток устроен значительно сложнее и благодаря взаимодействию белков тубулина и динеина способен изгибаться самостоятельно.
| у прокариот | у эукариот | |
| размеры органеллы (толщина, нм; длина, мкм) | 10-30 нм, 6–15 мкм | 200 нм, 100 мкм |
| белки жгутиковой нити | флагеллин | тубулин и динеин |
| мембрана вокруг филамента | отсутствует | присутствует |
| степень вращения | 360 ° | 180 ° |
| источник энергии | трансмембранный потенциал (у архей возможно АТФ) | АТФ |
| движение нити | пассивное | активное |
| субструктуры | филамент, крюк, базальное тельце | филамент, базальное тело (кинетосома) |
| строение филамента | сплошной (у архей) или полый белковый цилиндр | дублеты микротрубочек |
| структура базальной части | стержень, закрепленный в сложной системе колец или мембраноподобные органеллы (у архей) | триплеты микротрубочек |
Такое количество отличий свидетельствует об отсутствии гомологии между этими органоидами, то есть они не одинаковы по происхождению и строению, хоть и выполняют сходные функции.
Надцарство прокариот включает в себя царства архей и бактерий. Локомоторные структуры этих таксонов тоже не гомологичны друг другу, однако очень близки по строению. Жгутики архей изучены гораздо хуже.
Жгутики архей и бактерий
По способу перемещения подвижные бактерии подразделяются на плавающие и скользящие. Жгутики – это локомоторный орган плавающих микроорганизмов, позволяющий им развивать скорость от 20 до 200 мкм/сек.
Движение может быть спонтанным (если физико-химические характеристики среды одинаковы во всех направлениях) либо целенаправленным, когда бактерия стремится попасть в наиболее выгодные для нее условия. При адаптивном перемещении вращение базального мотора контролируется сенсорными системами.
По количеству и расположению жгутиков на клетке бактерии выделяют четыре морфологических типа микроорганизмов:
- монотрихи – имеют единственный жгутик;
- лофотрихи – характеризуются жгутиковым пучком на одном из клеточных полюсов;
- амфитрихи – имеют один или несколько жгутиков на обоих концах клетки;
- перитрихи – покрыты множеством жгутиков со всех сторон.

Тип жгутикования может быть как видовым признаком, так и результатом изменения условий культивирования или стадии жизненного цикла бактерии.
Жгутик архей во многом похож на бактериальный, однако имеет ряд отличий в ультраструктуре и механизме движения. Так, филамент у архей тоньше, построен из другого типа флагеллина, полый каналец в нити отсутствует. Длина крюка непостоянна, базальное тело имеет совсем другое строение и функционирует, скорее всего, на основе энергии АТФ. Археи движутся значительно медленнее бактерий.
Строение и функционирование бактериального жгутика
Жгутик бактерий образован тремя субструктурами: наружной нитью (филаментом), гибким сочленением (крюком) и базальным тельцем, заякоренным в клеточной оболочке. Синтез и сборку этих элементов кодируют около 50 fla-генов. За работу мотора отвечают mot-гены, а за адаптивные реакции – che-гены.
Филамент жгутика – это относительно жесткая белковая спираль, закрученная против часовой стрелки с образованием центрального полого канала диаметром до 3 нм. Такая конструкция способствует формированию спиральной траектории движения нити. По каналу филамента транспортируются молекулы флагеллина (FliC).
Крюк соединяет нить с базальным телом жгутика и состоит из двух типов белка: FlgE и FlgKl. Длина сочленения постоянна и составляет около 50 нм. Из-за изогнутой формы крюка при вращении мотора основание фибриллы описывает круг, благодаря чему возможно спиралевидное движение жгутика.
Базальное тельце закреплено в клеточной стенке и цитоплазматической мембране бактерий. Эта субструктура выполняет не только фиксирующую функцию, но и является мотором жгутика. Строение и локализация базального тельца зависят от типа клеточной стенки микроорганизма. У грамотрицательных бактерий оно состоит из двух внутренних (M и S) и двух внешних (P и L) колец, нанизанных на соединенный с крюком стержень. В состав базального тела также входит экспортная система, транспортирующая белковые элементы для сборки жгутика.
Структура базального мотора
В состав M-S-комплекса входят движущиеся структуры, называемые ротором, и переключатель направления вращения, который на более подробных схемах строения обозначают как С-кольцо. Вокруг ротора сосредоточены образованные белками MotAB ионные каналы – статоры. Мотор работает за счет энергии протонного (H + ) или натриевого (Na + ) градиента.
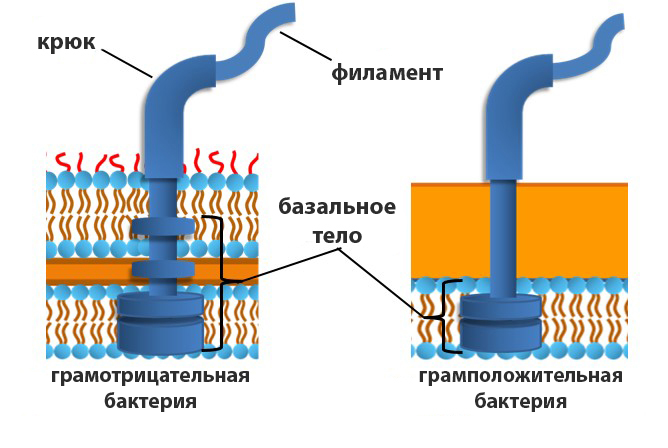
Расположение кольцевых субъединиц в клеточной оболочке соответствует следующей схеме:
- «М» – цитоплазматическая мембрана;
- «S» – периплазматическое пространство или клеточная стенка у Г + -бактерий;
- «P» – пептидогликановый слой;
- «L» – наружняя мембрана.
Внешние кольца P и L неподвижны и выполняют поддерживающую функцию. У грамположительных бактерий они отсутствуют.
Жгутик ядерной клетки
Эукариотический жгутик представляет собой цитоплазматический вырост клетки, состоящий из окруженной мембраной внешней части (ундулиподия) и погруженного в цитоплазму базального тела (кинетосомы).

Структурной основой ундулиподия является аксонема, состоящая из системы связанных друг с другом белковых цилиндров – микротрубочек. Их расположение соответствует формуле 9×2+2, – то есть девять периферических дублетов и две одиночные трубочки в центре (синглеты).
Дублеты образованы спаренными цилиндрами А и В, построенными из субъединиц белка тубулина. От каждой А-трубочки в сторону соседней пары отходит динеиновые рукоятки, которые преобразуют энергию АТФ в механическое движение. Дублеты соединены с синглетами радиальными спицами, а друг с другом – нексиновыми связками. Пространство между структурными элементами ундулиподия заполнено цитоплазмой.

Структура кинетосомы представлена девятью триплетами микротрубочек (формула 9+0), которые заякоривают жгутик в эукариотической клетке. Синглеты в базальном теле отсутствуют.
Источник



