
Локальный: паракринный, аутокринный, юкстакринный




![]()
![]()
Гормоны осуществляют свое действие на клетки-мишени на разных дистанциях (расстояниях) от места образования и выделения. В отличие от них действие аутакоидной системы является локальным.
 Система аутакоидоввключает химические сигнальные вещества, которые образуются практически во всех клетках организма и выделяются в межклеточную жидкость. Аутакоидная система относится, вместе с эндокринной (рис.3), к системе гуморальной регуляции. В отличии от эндокринной (системной) регуляции, аутакоиды действуют локально на специфические мембранные рецепторы клеток по механизму аутокринной, паракринной или юкстакринной регуляции. Аутокринное — воздействие клетки на саму себя (рис.4).
Система аутакоидоввключает химические сигнальные вещества, которые образуются практически во всех клетках организма и выделяются в межклеточную жидкость. Аутакоидная система относится, вместе с эндокринной (рис.3), к системе гуморальной регуляции. В отличии от эндокринной (системной) регуляции, аутакоиды действуют локально на специфические мембранные рецепторы клеток по механизму аутокринной, паракринной или юкстакринной регуляции. Аутокринное — воздействие клетки на саму себя (рис.4).
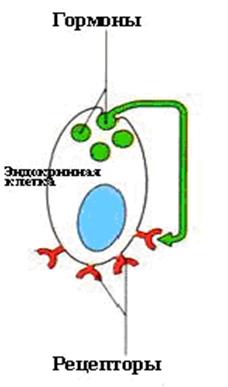
Рис.4 Аутокринное воздействие гормона.
Паракринным воздействием регулируется деятельность соседних клеток (рис.5).
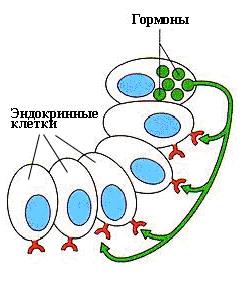
Рис.5 Паракринное воздействие гормона.
Юкстакринная – воздействие — разновидность паракринного действия, когда гормон не попадает в межклеточную жидкость, а сигнал передается через плазматическую мембрану рядом расположенной другой клетки.
 Слово аутакоиды греческого происхождения и означает auto — сам и acos – вещество. Чаще всего аутакоиды изменяют силу и характер ответа клеток на поступающие к мембране клетки электрические и химические сигналы, то есть выступают в роли модулятора. Регуляторными агентами в аутакоидной системе являются химические передатчики (сигналы): эйкозаноиды (простагландины, тромбоксаны, лейкотриены), цитокины, ангиотензин, нейротензин, оксид азота, кинины (кининоген, брадикинин, каллидин, тахикинин). Они не являются истинными гормонами, так как, во-первых, действуют исключительно местно, вблизи места синтеза, а во-вторых, возможность их взаимодействия со специфическими рецепторами существенно зависит от уровня местного метаболизма. Аутакоиды могут вырабатываться любыми, даже неспециализированными клетками организма. Аутакоиды выделяются в интерстициальную жидкость или кровь путем экзоцитоза. Перемещаясь в этих жидкостях, аутокринные агенты достигают рецепторов на поверхности своей или соседней клетки. Образующийся комплекс действует также как гормон-рецепторный комплекс эндокринной системы. Аутакоиды, несмотря на локальный характер действия играют важную роль в поддержании гомеостаза всего организма в целом.
Слово аутакоиды греческого происхождения и означает auto — сам и acos – вещество. Чаще всего аутакоиды изменяют силу и характер ответа клеток на поступающие к мембране клетки электрические и химические сигналы, то есть выступают в роли модулятора. Регуляторными агентами в аутакоидной системе являются химические передатчики (сигналы): эйкозаноиды (простагландины, тромбоксаны, лейкотриены), цитокины, ангиотензин, нейротензин, оксид азота, кинины (кининоген, брадикинин, каллидин, тахикинин). Они не являются истинными гормонами, так как, во-первых, действуют исключительно местно, вблизи места синтеза, а во-вторых, возможность их взаимодействия со специфическими рецепторами существенно зависит от уровня местного метаболизма. Аутакоиды могут вырабатываться любыми, даже неспециализированными клетками организма. Аутакоиды выделяются в интерстициальную жидкость или кровь путем экзоцитоза. Перемещаясь в этих жидкостях, аутокринные агенты достигают рецепторов на поверхности своей или соседней клетки. Образующийся комплекс действует также как гормон-рецепторный комплекс эндокринной системы. Аутакоиды, несмотря на локальный характер действия играют важную роль в поддержании гомеостаза всего организма в целом.
 Назовите способы передачи гормональных сигналов (см.рис.6).
Назовите способы передачи гормональных сигналов (см.рис.6).
Источник
Физиология эндокринной системы
Системы межклеточного управления
Уровни управления системами организма можно разделить на:
- внутриклеточные (управление на уровне клетки);
- межклеточные (согласовывают работу различных систем и органов целого организма).
В каждом случае системы управления могут быть неспециализироваными и специализированными. В неспециализированных системах управления соединения передают информацию вторично, основная их функция – источник пластического или энергетического материала. В специализированных системах главная функция соединений – передача информации. Поэтому такие вещества называют сигнальными. Внутриклеточная обработка информации осуществляется низкомолекулярными (вторичные посредники) и высокомолекулярными (белково-пептидные посредники) посредниками сигнальных соединений.
Три главные системы межклеточной передачи информации – нервная, эндокринная и иммунная, которые сейчас часто объединяют в нейроиммуноэндокринную систему. Каждая из них дистантно управляет жизнедеятельностью, но осуществляет это разными способами, подключая местное самоуправление. В зависимости от дистантности действия сигнального соединения различают местное (региональное) и системное управление (рис. 1 и 2).
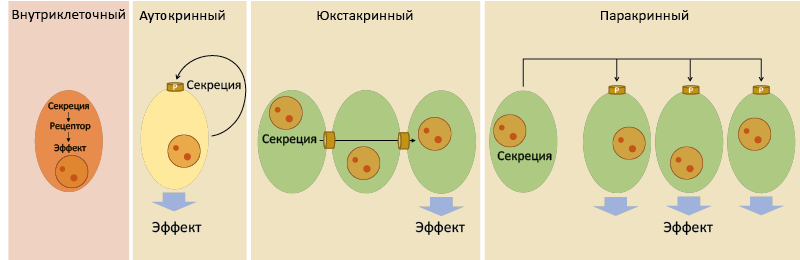
Рис. 1. Типы местного контроля
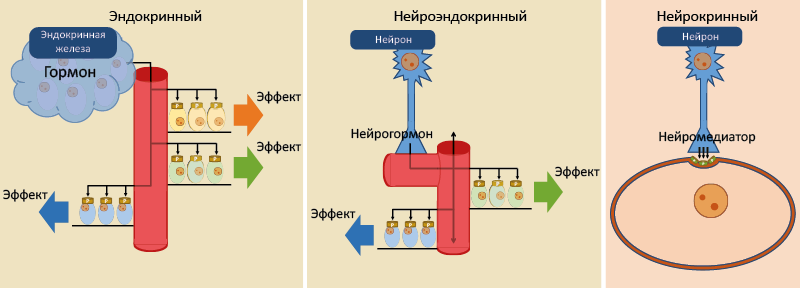
Рис. 2. Типы системного контроля
Местный контроль включает:
- Внутриклеточный;
- Аутокринный;
- Юкстакринный;
- Паракринный.
Системный контроль включает:
- Эндокринный;
- Нейрокринный;
- Нейроэндокринный.
Сигнальное соединение синтезируется в эффекторной клетке, где связывается со своим рецептором и вызывает характерные изменения. Чаще всего встречается в самих эндокринных клетках. Это наименее распространенный тип местного контроля.
Также предназначен для саморегуляции клетки, но сопровождается секрецией сигнального соединения. После выделения сигнальное соединение связывается с рецептором на секретирующей клетке и оказывает своё действие.
Сигнальное соединение не секретируется, а транспортируясь через поры щелевых контактов, достигает соседних клеток и индуцирует в них эффект.
Сигнальное соединение выделяется клетками определенного органа или ткани и действует на соседние клетки того же органа. Это самый распространенный тип местного контроля. Сигнальные вещества с преобладанием паракринных эффектов часто называют паракринными факторами или гистогормонами. Передача информации нейротрансмиттерами – один из примеров специализированной паракринной передачи.
Эндокринный (гормональный) контроль
Гормон выделяется эндокринной железой, эндокринной или эффекторной клеткой и поступает в системный кровоток, действуя на все структуры организма с соответствующим рецептором. Эффекты гормона зависят от типа рецептора и р еагирующей ткани.
Нейрогормон секретируется терминалями аксонов и через капиллярное сплетение поступает в системный кровоток.Дальнейшие события аналогичны эндокринному способу регуляции.
Нервную систему можно рассматривать как систему с паракринным способом регуляции, поскольку нейромедиаторы действуют на коротком расстоянии на близлежащие элементы через специализированные рецепторы. Дистантность действия достигается за счёт длины аксона и синаптических переключений.
Многообразие сигнальных функций гормонов достигается благодаря сочетанию дистантного и локального действия (рис. 3), но спектр эффектов на системном и местном уровнях может существенно отличаться (рис. 4). Стоит отметить, что сигнальное соединение причисляется к гормонам при наличии у него не только паракринных, но и выраженных системных эффектов.
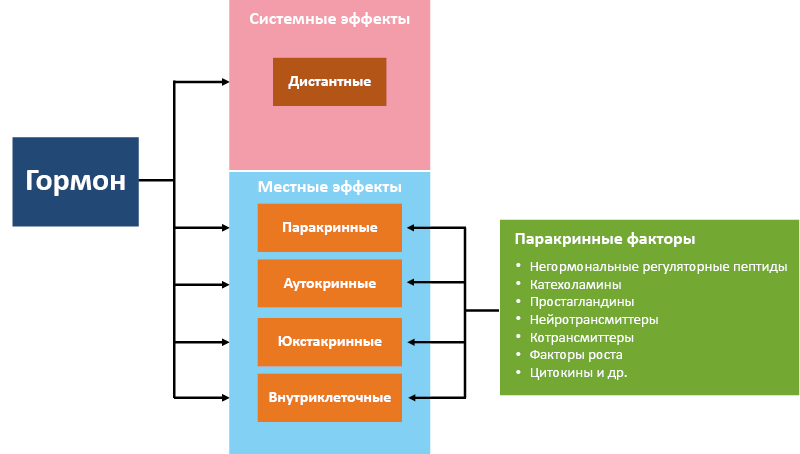
Рис. 3. Дистантность действия гормонов

Рис. 4. Отличия системных и местных эффектов гормонов
Системные эффекты предназначены для управления процессами жизнедеятельности.Гормоны определяют процессы роста и развития, половой дифференцировки, размножения, обмена веществ, участвуют в неспецифической адаптации и взаимной регуляции гормональной сети. Системные эффекты гормонов подразделяют на:
- необратимые программирующие (детерминирующие);
- обратимые регуляторные.
Обычно соединение оказывает детерминирующие эффекты при первичном воздействии на экспрессию генов в ограниченные критические периоды онтогенеза (чаще в эмбриональном и неонатальном периодах). При этом необратимо изменившийся уровень экспрессии данных генов меняет фенотип клетки, который сохраняется у её потомков даже в отсутствие гормона. Если затрагивается экспрессия только нескольких генов, то в клетке необратимо меняется активность метаболических систем, но фенотип остаётся прежним.
Паракринное действие гормонов призвано суммировать поступающую к ткани информацию, координировать её суммарный ответ на внешний сигнал, усиливать гормональный эффект. Многие локальные эффекты гормонов сходны с эффектами паракринных факторов:
- поддержание активности секретирующей ткани;
- регуляция про- и антипролиферативной активности клеток;
- регуляция ангиогенеза;
- регуляция воспалительных реакций.
Гормоны могут секретировать клетки разного уровня специализации (рис. 5):
- эндокринные;
- со смешанными функциями;
- эффекторные.
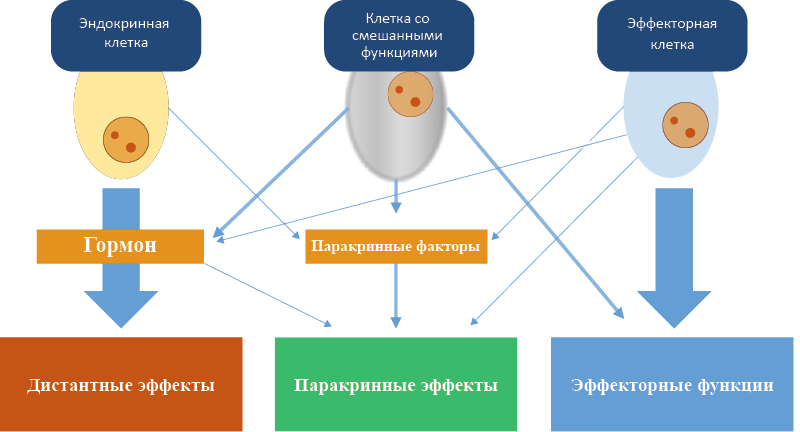
Рис. 5. Степень специализации гормонпродуцирующих клеток
Эндокринные железы, как правило, выделяют сразу несколько типов гормонов (см. Приложение 1). И важно отметить, что гормоны той же структуры, что продуцируются эндокринными железами, могут синтезироваться местно, в эффекторных клетках и тканях (см. Приложения 2 м 3).
Высоко специализирована на продукции гормонов, способна секретировать ряд паракринных факторов (напр., клетки эндокринных желез).
Клетка со смешанными функциями
В дополнение к продукции гормонов и паракринных факторов имеет дополнительные эффекторные функции (напр., клетки Сертоли продуцируют эстрогены, активин, ингибин и другие гормоны, одновременно поддерживая развитие сперматозоидов).
Эффекторная клетка (Приложение 4)
Специализирована на выполнении отдельной физиологической функции, но способна секретировать гормоны и паракринные факторы (напр., адипоциты специализированы на липидном обмене, секретируют гормоны лептин, адипонектин, резистин и другие системные регуляторы жирового обмена и пищевого поведения).
Эндокринные клетки могут группироваться различным образом:
- встраиваясь в неэндокринные органы отдельными клеточными элементами;
- входя в состав желез смешанной секреции группами клеточных элементов;
- формируя отдельный орган – эндокринную железу.
Эндокринные железы (рис. 6) содержат клеточные элементы разного эмбрионального происхождения, однако у низших позвоночных эндокринные клетки разных типов не всегда объединяются в единый орган (рис. 7, 8). Соединение нескольких эндокринных клеток в железу облегчает паракринный обмен информацией. Биологический смысл подобной интеграции эндокринных клеток – увеличение контроля за потоками информации и эффекторными функциями (рис. 6).
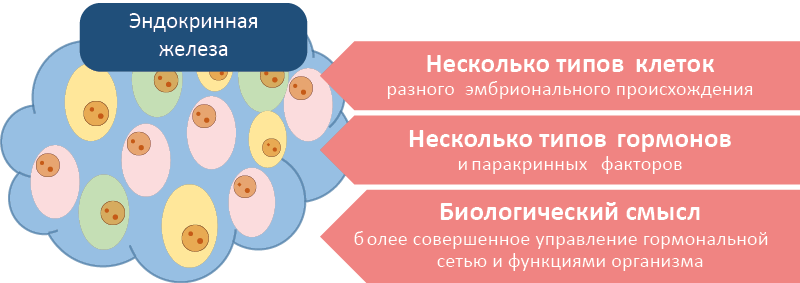
Рис. 6. Эндокринная железа
Эндокринные железы – эволюционное приобретение. Например, у акуловых интерреналовая ткань (гомолог коры надпочечников) в соответствии со своим названием располагается между почками, а хромафинная ткань (гомолог мозгового слоя) разбросана в виде узлов вдоль почек. В ходе эволюции у хвостатых амфибий объединились интерреналовая и хромафинная ткани. Впоследствии это объединение расширилось и п ривело к оформлению отдельных надпочечниковых желез высших позвоночных (рис. 7).
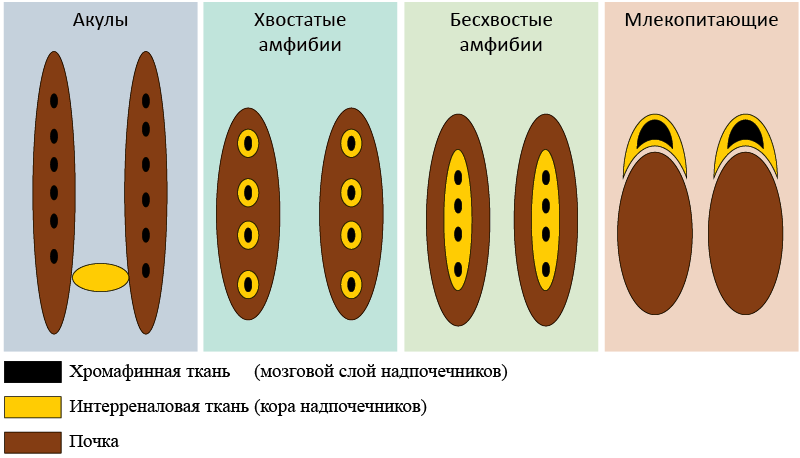
Рис. 7. Эволюция надпочечников позвоночных
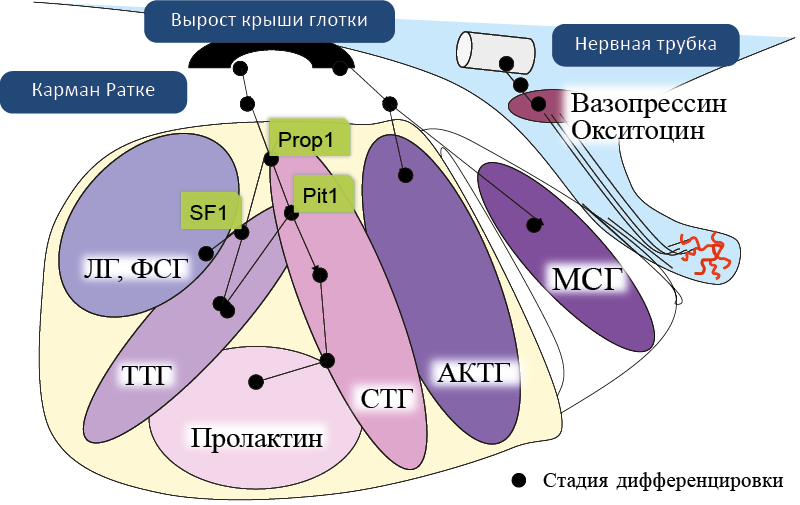
Рис. 8. Эмбриогенез гипофизамлекопитающих, топография гормонпродуцирующих клеток
Передняя и задняя доли гипофиза развиваются из разных эмбриональных зачатков – эктодермы глотки (кармана Ратке) и нейроэктодермы нервной трубки, соответственно. Соматорофы, лактотрофы, тиреотрофы и гонадотрофы передней доли гипофиза развиваются из дорзо-вентральной части кармана Ратке, а кортикотрофы передней доли и клетки средней доли гипофиза – из его дистальной части, тесно контактирующей и срастающейся с нейрогипофизом. Последовательное действие транскрипционных факторов prop-1 и pit-1 индуцирует развитие соматотрофов и лактотрофов переднего гипофиза, а также влияет на дифференцировку тиреотрофов. В дифференцировке гонадотрофов участвуют факторы транскрипции prop-1 и SF-1.
- Prop-1 – фактор транскрипции, определяющий продукцию pit-1;
- pit-1 – транскрипционный фактор гипофиза;
- SF-1 – стероидогенный фактор 1.
Расшифровку сокращений гормонов см. в Приложении 1.
Функции гормонов можно условно разделить на две принципиально разные группы:
- эффекторные:
контроль внешних 1 структур, обеспечивающих жизнедеятельность организма; - сигнальные:
координация информационных связей внутри и вне эндокринной системы.
Сигнальную роль гормоны выполняют внутри соответствующей вертикальной/горизонтальной оси или между осями. В Приложении 8 представлены основные, но далеко не все, эффекторные и сигнальные функции наиболее изученных гормонов.
Эти функции связаны с действием гормонов на ткани-мишени, непосредственно участвующие в регуляции обмена веществ и поддержании физиологических процессов. Эффекторные функции гормонов могут быть необратимыми (в раннем онтогенезе – часто морфогенными) и обратимыми регуляторными. Примером подобных функций служит морфогенное и регуляторное действие андрогенов на развитие и функционирование простаты и других мужских половых органов; обратимая стимуляция глюкокортикоидами глюконеогенеза в печени и др.
Функции заключаются в поддержании/регуляции гормоном активности как своей гормональной оси, так и гормональной сети в целом. Подобные информационно-координирующие свойства гормонов также задействованы в координации других сигнальных систем: нервной и иммунной. Так, андрогены помимо указанных выше эффекторных функций входят в состав репродуктивной гормональной оси и регулируют ее активность. Они тормозят секрецию гонадотропинов гипофиза, гонадолиберина гипоталамуса и по механизму отрицательной обратной связи контролируют свой уровень в кровотоке. Но андрогены также влияют на продукцию гормонов других осей (напр., атриопептидов сердца, включаясь в регуляцию водно-солевого обмена) и дифференцировку мозга по мужскому типу, адаптируя активность ЦНС к потребностям мужских особей. Обычно сила регуляторного воздействия максимальна на собственную ось, и слабее – на другие оси. Соотношение эффекторных и информационных функций у разных групп гормонов может существенно различаться. У гормонов гипоталамо-гипофизарной системы чаще более выражены информационные функции, у гормонов периферических желез и клеток с эндокринной секрецией информационные и эффекторные функции сопоставимы.
Источник



