Самый обширный отдел водорослей, включающий от 13 000 до 20 000 видов. Обитают в основном в пресных водоемах, имеют зеленую окраску вследствие преобладания хлорофилла a и b по количеству над другими пигментами (каротиноидами, ксантофиллами). Этот отдел включает в себя одноклеточные, многоклеточные и колониальные формы. Большинство из них растет на глубине 20-40 метров.
Клеточная стенка зеленых водорослей образована целлюлозой, запасное питательное вещество — крахмал. У многих представителей в жизненном цикле наблюдается чередование полового поколения (гаметофита) и бесполого (спорофита).
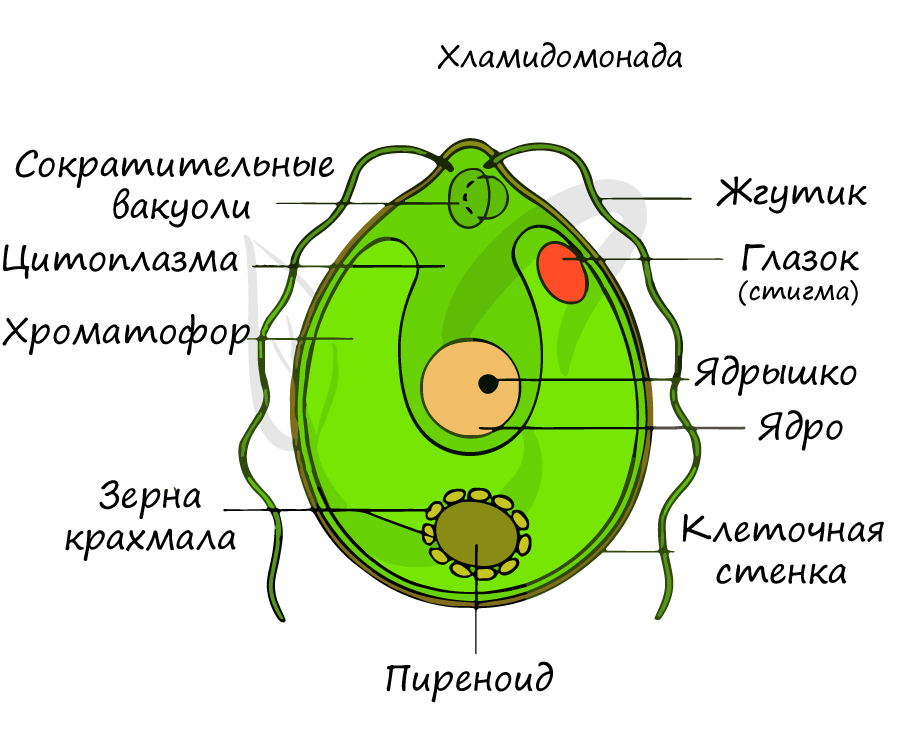
Хламидомонада
Хламидомонада — одноклеточная двужгутиковая зеленая водоросль, обитающая в лужах, пресных водоемах, прудах. Форма клетки грушевидная. На переднем конце тела имеет два жгутика, за счет которых активно движется.
Светочувствительный глазок (стигма) помогает хламидомонаде занять наиболее освещенное место для активного процесса фотосинтеза, который идет в хроматофоре. Сократительные (пульсирующие) вакуоли клетки удаляют избыток постоянно поступающей внутрь воды, таким образом, они поддерживают осмотическое давление на уровне, необходимом для жизни.
Хламидомонада имеет чашевидный хроматофор с пиреноидом — округлой белковой гранулой, содержащей фермент, который участвует в синтезе сахаров. Вокруг пиреноида запасается крахмал.
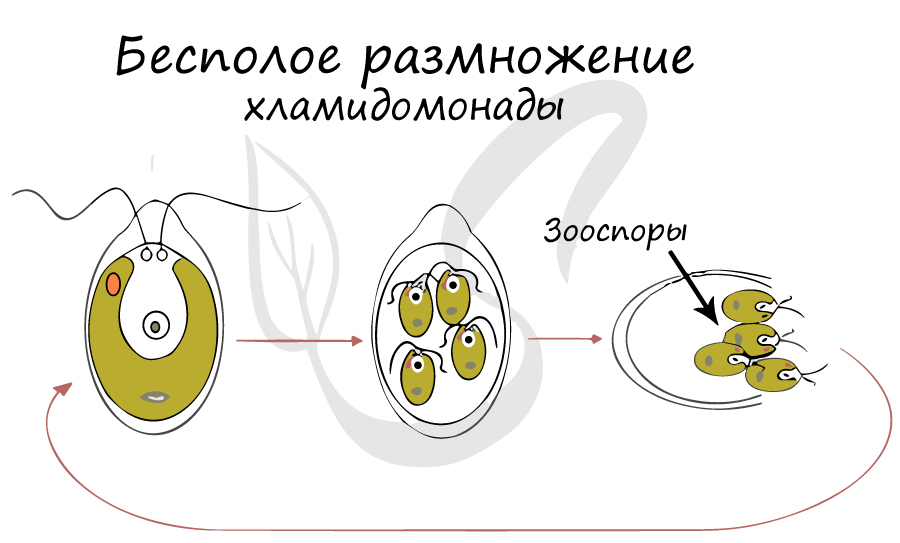
Размножение хламидомонады
Размножается преимущественно бесполым путем. Половой процесс — конъюгация.
При благоприятных условиях (летом) размножается бесполым путем с помощью зооспор. Хламидомонада (n) дважды делится митотически без разрыва материнской оболочки, в результате образуются 4 клетки (n). Они растут, у каждой из них развивается жгутик, появляется глазок и клеточная стенка. С течением времени материнская оболочка, окружающая клетки, разрывается, и зооспоры выходят во внешнюю среду. Из каждой зооспоры развивается взрослая клетка.
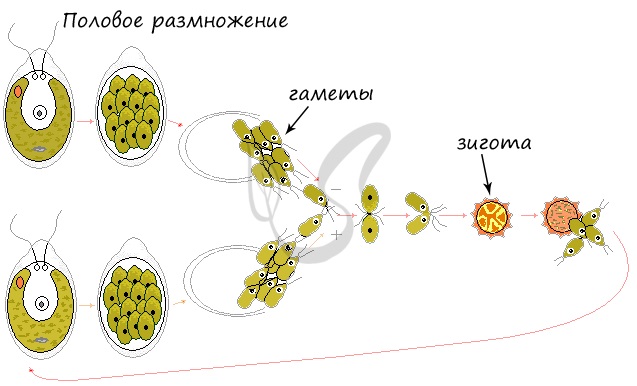
Рассмотрим изогамный половой процесс, при котором гаметы не отличаются по строению, внешнему виду, одинаково подвижны.
Половое размножение активируется при наступлении неблагоприятных условий (пересыхание водоема, понижение температуры внешней среды). Внутри хламидомонады (n) путем митоза образуются половые клетки — гаметы (n). Запомните, что в половом размножении всегда участвуют половые клетки 😉
Гаметы (n) разных хламидомонад попарно сливаются, в результате чего образуется зигота (2n), которая покрывается плотной защитной оболочкой — цистой. При благоприятных условиях зигота (2n) делится мейозом, по итогам которого образуются 4 хламидомонады (n).
Красный снег
Красный снег — явление, характерное для приполярных областей Земли, также встречается на высоких горах. Снег приобретает нехарактерную красную окраску, связанную с массовым размножением Хламидомонады снежной, клетки которой содержат красный каротиноид — астаксантин. Для особей этого вида благоприятными являются низкие температуры, при температуре выше +4 °С они погибают.
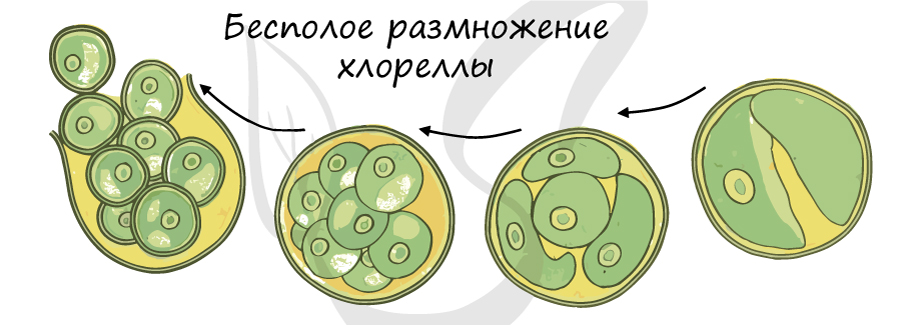
Хлорелла
Хлорелла — одноклеточная зеленая водоросль без жгутиков, обитающая в самых разных средах: на сырой почве, на стволах деревьев, скалах, в соленой и пресной воде. Ее скопления хорошо заметны в виде налета зеленого цвета.
Клетка содержит чашевидный хроматофор (имеет вид сильно вырезанной чаши), запасающий крахмал. Хлорелла отличается быстрым темпом деления клеток, в связи с этим ее используют для получения кормов. Фотосинтез у нее также идет очень интенсивно. Эта водоросль одной из первых побывала в космосе, ее используют на космических кораблях для получения кислорода.
Размножение осуществляется только бесполым путем, содержимое материнской клетки делится митотически на 4 или 8 дочерних клеток, после чего оболочка материнской клетки рвется, и дочерние клетки выходят наружу, развиваются во взрослых особей, после чего снова делятся.
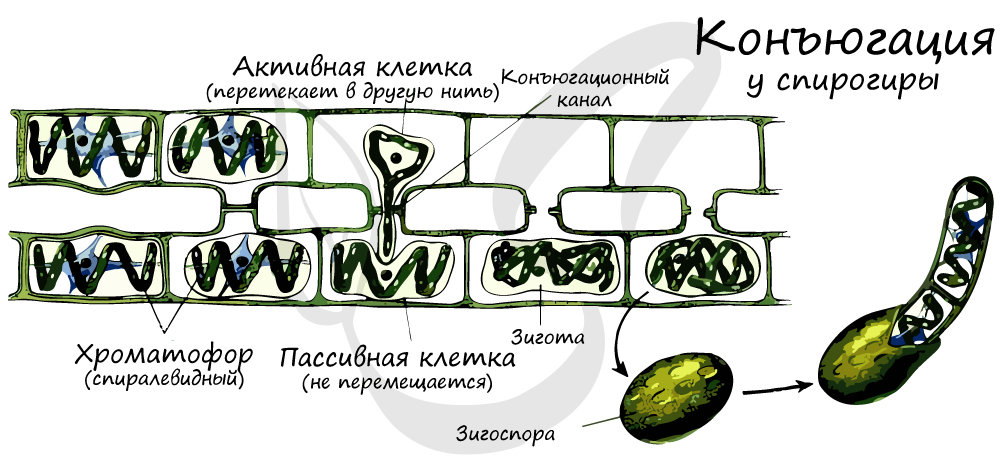
Спирогира
Спирогира — многоклеточная нитчатая зеленая водоросль. Скопления нитей спирогиры на поверхности рек и прудов образуют тину.
Хроматофор у спирогиры спиралевидный, представлен в виде одной или нескольких лент, опоясывающих клетку в пристенном слое цитоплазмы. В клетке содержится крупное ядро, расположенное в центре и подвешенное на тяжах цитоплазмы.
Размножается бесполым и половым путями.
Бесполое
Бесполое (вегетативное) размножение может осуществляться частями таллома: нить водоросли разрывается на отдельные участки, или даже клетки, которые дают начало новому организму.
Половой процесс — конъюгация. Две нити водоросли располагаются параллельно, клетки сближаются, у них образуются боковые выросты. При соприкосновении боковых выростов между клетками разных нитей водорослей образуется копуляционный канал, по которому происходит перемещение содержимого одной клетки (n) в другую (n), после чего сливаются цитоплазмы и ядра, образуя зигоспору (2n).
После периода покоя зигоспора (2n) делится мейозом, образуются четыре клетки (n), из которых только одна прорастает в новую особь, а три остальных — погибают.
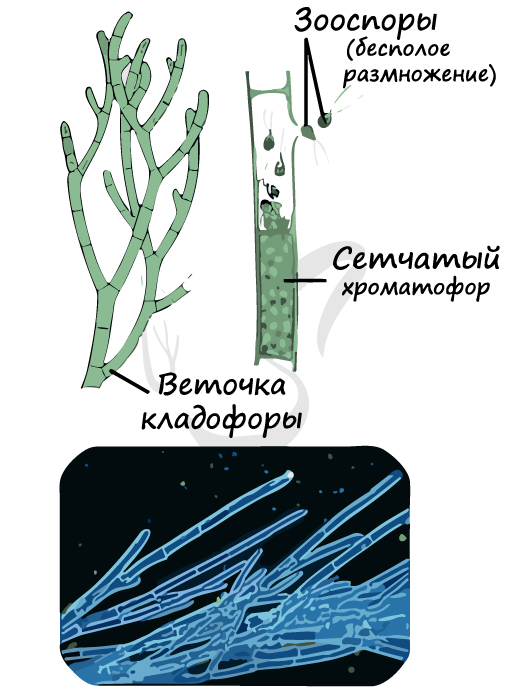
Кладофора
Кладофора — многоклеточная нитчатая зеленая водоросль. Ее ветвящиеся нити непрочно прикреплены к субстрату, от которого часто отрываются. Хроматофор имеет вид сеточки (сетчатый). Бесполое размножение осуществляется с помощью зооспор, половое размножение в форме изогамии.
Улотрикс
Улотрикс — многоклеточная нитчатая зеленая водоросль. Обитает в пресной и морской воде, образует на подводных объектах зеленый налет — тину. Хроматофор в виде незамкнутого кольца (пояска), содержит пиреноид. Преимущественно размножается бесполым путем, с помощью четырехжгутиковых зооспор. Есть возможность полового размножения по типу изогамии.
В цикле развития улотрикса преобладает гаметофит (n) — вегетативное гаплоидное поколение. Также заметьте, что гаметы улотрикса (n) образуются из клеток слоевища (n) путем митоза.
Плеврококк
Скорее всего, любой гетеротроф сделает ошибку, первый раз встретив это название 🙂 Уж слишком сильно оно смахивает на название бактерий, таких как стафилококки, стрептококки. Запомните и не ошибайтесь: плеврококк — зеленая водоросль. Плеврококк имеет клетки шаровидной формы, они могут быть одиночные или соединенные в группы. Видимые вакуоли в клетке отсутствуют, хроматофор в виде пластинки, не содержит пиреноидов.
Плеврококк распространен повсеместно, способен вынести полное пересыхание. Образует зеленый налет на стволах деревьев, поверхности скал и почве.
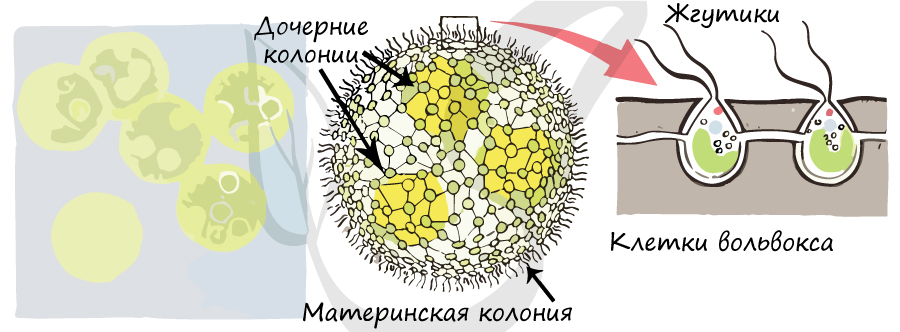
Вольвокс
«Вольвокс» означает «катящийся». Представляет собой зеленую подвижную колониальную водоросль, имеющую шаровидную форму. Одна колония вольвокса может достигать 3мм, а по количеству клеток — 200 до 10 тысяч.
Клетки расположены на периферии, соединены между собой тяжами цитоплазмы — протоплазматическими нитями, обеспечивают движение колонии и питание. В центре колонии имеется полость, занятая слизью. Каждая из клеток на периферии имеет два жгутика, обращенных во внешнюю среду, клетки напоминают хламидомонаду.
Вольвокс играет очень важное эволюционное значение, и помогает сделать вывод о том, что развитие живых организмов от одноклеточных форм к многоклеточным происходило через колониальные формы.
Большая часть клеток в колонии вольвокса вегетативные. Вегетативное размножение вольвокса происходит с помощью дочерних колоний внутри материнской, особыми клетками — партеногонидиями. Эти клетки делятся митозом перпендикулярно поверхности шара. В результате образуется пластинка, которая выворачивается и образует дочерний шар. Дочерние шары разрастаются, при этом происходит разрыв материнского организма (шара).
Половой процесс происходит в специализированных местах — антеридиях, где развиваются сперматозоиды (n), и оогониях, где созревают яйцеклетки (n). Сперматозоид проникает в оогоний, образуется зигота, или ооспора (2n). При благоприятных условиях зигота делится мейозом, образуются клетки вольвокса (n), которые затем делятся множеством митотических делений.
Таким образом, основная форма существования клеток в колонии вольвокса — гаплоидна (n), диплоидна в жизненном цикле только зигота (2n).
Сине-зеленые водоросли
Спешу предупредить об очень частом заблуждении! Сине-зеленые водоросли — это вовсе не водоросли, их по-другому называют цианобактерии. Они представляют собой отдел крупных грамотрицательных бактерий, которые способны выделять кислород в процессе фотосинтеза.
Эволюционно сине-зеленые водоросли — очень древние микроорганизмы, которые возникли в архее. Им отведена крайне важна роль: они являются первыми фотосинтезирующими организмами. Благодаря им 2 млрд. лет назад в атмосфере Земли впервые появился кислород.
У них отсутствуют жгутики, они могут иметь нитчатую или колониальную форму, или же быть одноклеточными. Относительно крупные размеры цианобактерий и сходство в строении с водорослями было изначальной причиной их рассмотрения в составе растений. На настоящее время доказано сходство цианобактерий с остальными бактериями.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
1) Указание типа питания животного: смешанный тип питания — и автотрофный (на свету) и гетеротрофный (в темноте или когда много органических веществ); или ответ — миксотрофный тип питания.
2) Обоснование, например: На свету фотосинтезирует — значит, автотрофный тип питания. В темноте или когда много органических веществ — питается готовыми органическими веществами — всасывая их всей поверхностью, значит, гетеротрофный тип питания. Такой тип питания называется — миксотрофный.
Какой тип питания характерен для хламидомонады, изображённой на рисунке? Обоснуйте свой ответ.
Определите тип питания организмов, приведённых в списке.
Запишите цифры, под которыми указаны организмы в списке, в соответствующую ячейку таблицы.
Ответ в каждую ячейку запишите в виде последовательности цифр, в порядке их возрастания.
1) Указание типа питания животного: смешанный тип питания — и автотрофный (на свету) и гетеротрофный (в темноте или когда много органических веществ); или ответ — миксотрофный тип питания.
2) Обоснование, например: на свету фотосинтезирует — значит, автотрофный тип питания. В темноте или когда много органических веществ — питается готовыми органическими веществами — всасывая их всей поверхностью, значит, гетеротрофный тип питания. Такой тип питания называется — миксотрофный.
Какой тип питания характерен для папоротника, изображённого на рисунке? Обоснуйте свой ответ.
Определите тип питания организмов, приведённых в списке.
2) холерный вибрион
5) дождевой червь
Запишите цифры, под которыми указаны организмы в списке, в соответствующую ячейку таблицы.
Ответ в каждую ячейку запишите в виде последовательности цифр, в порядке их возрастания.
2) Обоснование, например: на свету фотосинтезирует — значит, автотрофный тип питания. Образует из неорганических веществ органические на свету — значит, автотрофный тип питания.
Какой тип питания характерен для кукушкиного льна, изображённого на рисунке? Обоснуйте свой ответ.
Определите тип питания организмов, приведённых в списке.
4) печеночный сосальщик
6) кишечная палочка
Запишите цифры, под которыми указаны организмы в списке, в соответствующую ячейку таблицы.
Ответ в каждую ячейку запишите в виде последовательности цифр, в порядке их возрастания.
19.2.5. Порядок 5. Улотриксовые — Ulotrichales (Н. А. Мошкова)
Порядок Ulotrichales объединяет около 460 видов водорослей, относящихся к 70 родам. В подавляющем большинстве это многоклеточные индивиды нитчатой, разнонитчатой или пластинчатой * структуры, реже малоклеточные до одноклеточных или колониальные, микроскопические или различимые невооруженным глазом, нередко макроскопические, достигающие в длину до нескольких дециметров, окрашены обычно в зеленый цвет разных оттенков, немногие желтовато-оранжевые или оранжево-красные (виды рода Trentepohlia Mart.). У некоторых видов оранжево-красна я окраска наблюдается лишь на определенных стадиях развития.
* ( Строение тела особи дано по М. М. Голлербаху [78].)
Одноклеточные формы обычно образуют временные короткие нити из небольшого числа клеток или плотные скопления. Нитчатые талломы простые или разветвленные,однорядные, реже многорядные, часто покрыты слизью разной толщины и структуры, при определенных условиях легко распадаются на малоклеточные фрагменты и даже отдельные клетки. У форм, живущих в свободном состоянии, отсутствует морфологически уловимое различие между вершиной и основанием нити (Binuclearia Wittr., Geminella Тигр., Gloeotila Kütz., Radiofilum Schmidle, Chlorhormidium Fott и др.), у прикрепленных организмов (виды родов Ulothrix Kütz., Uronema Lagerh. и др.) базальная и апикальная клетки отличаются от остальных клеток таллома. Апикальные клетки полушаровидные, тупые или заостренные, прямые, реже изогнутые, базальные — обычно имеют форму довольно вытянутого конуса, суженной частью обращенного к субстрату. На нижнем конце базальных клеток могут возникать пектозные или целлюлозные образования в виде полушаровидных подушечек или дисковидных стоп с ровными или изрезанными краями (дермоиды), иногда базальная клетка вытягивается в ризоид.
Разнонитчатые талломы представляют собой дальнейшее морфологическое усложнение нитчатого тела и характерны для большого числа родов улотриксовых. Разнонитчатая структура представлена не только разветвленными в плоскости талломами, но и восходящими нитями, у просто организованных улотриксовых — простыми, у более сложно организованных — разнообразно разветвленными. Усиление или ослабление развития стелющейся или вертикальной частей таллома способствовало возникновению различных модификаций разнонитчатой структуры.
При более или менее полном исчезновении вертикальных нитей таллом приобретает вид разнообразных однослойных пластинок (Protoderma viride Kütz., Epibolium dermaticola Printz) или дисков (Coleochaete scutata Breb., C. orbicularis Pringsh.).
При недоразвитии или выпадении горизонтальной части интенсивно развитыми оказываются вертикальные нити (виды родов Draparnaldia Bory, Draparnaldiella Meyer et Skabitsch. и др.). Разветвленные формы обычно кустиковидные, при этом у одних представителей нет резкого различия между главными нитями и боковыми ветвями (Stigeoclonium Kütz., Microthamnion Näg.), у других эти различия выражены ярко (Cloniophora Tiffany, Draparnaldiopsis Sm. et Klyver и др.).
Для некоторых разветвленных водорослей характерно образование большого количества слизи объединяющей нити в полушаровидные и лопастные колонии (Chaetophora Schrank). Слизь образуется в результате видоизменения пектиновых слоев оболочки клеток или обверток нитей.
Двуслойные пластинчатые талломы улотриксовых обычно плоские, листовидно расширенные или лентовидные. Если между слоями таллома образуется полость — тогда пузыревидные, мешковидные (Monostroma Thur.) или трубчатые с однослойными стенками, простые или разветвленные (Enteromorpha Link).
На поверхности многих клеток стелющихся нитей и однослойных пластинчатых талломов образуются преимущественно одноклеточные волоски., иногда луковицеобразно вздутые у основания, или щетинки. Многие апикальные клетки кустиковидных талломов нередко вытягиваются в одноклеточный, заостренный, чаще многоклеточный, тупой волосок.
Клетки цилиндрические, боченковидные, эллипсоидные, шаровидные, яйцевидные, веретеновидные, иголковидные, прямые или изогнутые, на концах тупые или заостренные, преимущественно со сплошной двуслойной оболочкой, обычно бесцветной, реже окрашенной соединениями железа в бурый цвет. Внутренний слой оболочки тонкий, целлюлозный, наружный — более толстый, с примесью пектиновых веществ, гомогенный или слоистый, с продольным или косым расположением слоев, легко ослизняющийся; оболочки из двух равных частей, не имеющие формы буквы Н, встречаются только у видов рода Radiofilum. Все клетки в нитях одеты снаружи еще своеобразной обверткой (влагалищем) из пектиновых веществ, обычно тонкой, хорошо заметной лишь у поперечных перегородок, реже довольно толстой, гомогенной или отчетливо слоистой (фиброзной), нередко легко ослизняющейся. Цитоплазма прилегает к клеточным стенкам; середину клетки занимает центральная вакуоль с клеточным соком. Ядро преимущественно одно, шаровидное или слегка эллипсоидное, незначительных размеров, мало заметное, обычно размещается в пристенном слое цитоплазмы, реже в особом слизистом тяже, проходящем посередине клетки или смещенном к одной из ее сторон. Хлоропласт обычно один, пристенный, у одних видов дисковидный, пластинчатый, прямой или согнутый в форме полуцилиндра с ровными или лопастными краями, реже лентовидно вытянутый и слегка спирально изогнутый, у других — в форме широкого или узкого цилиндра (кольца), цельного или продырявленного нередко настолько сильно, что принимает вид сеточки, выстилающей продольные стенки клеток. У водорослей семейства Prasiolaceae хлоропласт центральный, звездчатый. Чашевидный хлоропласт встречается преимущественно у репродуктивных клеток. Хлоропласты улотриксовых содержат хлорофиллы а и b и каротиноиды (преимущественно β-каротин и ксантофилл).
Внутри хлоропластов, реже на их поверхности располагаются пиреноиды, преимущественно скорлупчатые, окруженные покровом из ассимилятов; реже пиреноиды отсутствуют.
Продукт ассимиляции — крахмал или маслоподобное вещество, бесцветное (у видов рода Microthamnion Näg.) или окрашенное (у видов рода Trentepohlia).
Размножаются улотриксовые водоросли как бесполым, так и половым путем. Вегетативное размножение происходит путем фрагментации тела водорослей на отдельные многоклеточные или одноклеточные участки или отдельными клетками с утолщенной оболочкой и большим количеством запасных питательных веществ — акинетами размножения. Подобные акинеты весьма часто встречаются у представителей родов Ulothrix, Binuclearia Gloeotila, Chlorhormidium, Cylindrocapsa Reinsch., a также многих хетофоровых водорослей. Собственно бесполое размножение осуществляется большей частью подвижными четырехжгутиковыми или двужгутиковыми зооспорами, реже неподвижными апланоспорами и гипноспорами. Половое размножение — посредством гамет, возникающих в неизмененных, слегка измененных или значительно преобразованных клетках — гаметангиях. Гаметы обычного монадного типа, двужгутиковые. У улотриксовых известны все три формы полового процесса — изо-, гетеро- и оогамия. При изогамии, если обе копулирующие гаметы развиваются на одной особи, водоросли называются гомоталличными, если копуляция возможна только между гаметами, образовавшимися на разных особях, — гетероталличными. При этом гаметы морфологически совершенно подобны друг другу и различия между ними являются чисто физиологическими. При гетерогамии обе гаметы также подвижны, но различаются между собой по величине. Более крупные и менее подвижные гаметы (макрогаметы) считаются женскими, меньшие по величине и более подвижные (микрогаметы) — мужскими. У форм с гетерогамным половым процессом наблюдается разная степень дифференциации мужских и женских гамет. В одних случаях различия невелики, в других весьма значительны. У некоторых улотриксовых водорослей с гетерогамным половым процессом наблюдается своеобразный переход от гетерогамии к оогамии (виды родов Aphanochaete А. Br. и Chaetonema Nowak.). Настоящая оогамия встречается в семействах Cylindrocapsaceae и Coleochaetaceae.
У некоторых улотриксовых водорослей одна и та же особь может давать начало как зооспорам, так и гаметам, у других — зооспоры и гаметы образуются на разных особях, т. е. цикл развития водорослей включает в себя как половую (гаметофит), так и бесполую (спорофит) форму развития. У многих улотриксовых водорослей, размножающихся половым путем, жизненный цикл проходит в основном в гаплоидной фазе, диплоидной оказывается только зигота, в которой, при ее прорастании и происходит мейоз (зиготическая редукция). Такой гаплофазный цикл развития свойствен многим видам водорослей, принадлежащим к подпорядкам Ulotrichineae и Chaetophorimae. В тех случаях, когда редукционное деление ядра происходит в спорангии, при образовании спор (спорическая редукция), часть жизненного цикла водоросли от зиготы до образования спор проходит в диплоидной фазе, а часть от споры до образования гамет — в гаплоидной фазе. Такой диплогаплофазный цикл развития наблюдается у видов подпорядка Ulvinea. В морфологическом отношении здесь наблюдается смена форм развития двух типов: изоморфная, когда обе формы бесполая (спорофит) и половая (гаметофит) морфологически подобны друг другу (известна у родов Ulva и Enteromorpha), и гетероморфная, когда они морфологически различны (встречается у рода Monostroma). Гаметофит моностромы многоклеточный, пластинчатый, крупный, спорофит — одноклеточный, мелкий.
У видов рода Prasiola Ag. (Р. stipitata Suhr) обнаружена соматическая редукция. Мейоз происходит в вегетативных клетках верхней части диплоидного гаметофита, где возникают участки гаплоидных клеток, в которых образуются гаплоидные гаметы. Таким образом, при соматической редукции вегетативные клетки из разных участков одной и той же особи могут отличаться по плоидности их ядер.
Единого мнения об объеме порядка Ulotrichales и его месте в системе нитчатых зеленых водорослей до сего времени не существует. Одними исследователями улотриксовые рассматриваются в широком объеме в ранге порядка класса Chlorophyceae, другими — в том же объеме, но в ранге класса Ulotrichophyceae. Некоторые принимают порядок Ulotrichales в узком объеме, как один из порядков класса Ulotrichophyceae, но и в этом случае неоднозначно [65, 221, 222, 359, 414, 416, 417, 423, 495, 580].
Мы рассматриваем порядок Ulotrichales в классе собственно зеленых водорослей. Он представляет собой весьма четко очерченную систематическую группу, которая по своим морфологическим особенностям хорошо отличается от других равноценных по рангу систематических групп класса Chlorophyceae. Порядок Ulotrichales в нашем представлении объединяет нитчатые, разнонитчатые, пластинчатые и трубчатые структуры, которые могут быть правильно поняты только как производные нити.
Вопрос о происхождении Ulotrichales решается по-разному. Существует точка зрения, что возникновение нитчатого таллома зеленых водорослей было связано с непосредственным переходом древних примитивных монадных или коккоидных форм к прикрепленным условиям существования [305]. У прикрепленной клетки постепенно возникла и закрепилась способность к неограниченному росту ее апикальной части с последующей сегментацией протопласта и образованием поперечных перегородок. Высказывается также мнение о возможности другого пути возникновения порядка Ulotrichales из хлорсарциновых водорослей, которые, будучи тесно связанными с хлорококковыми, отличаются вегетативным делением клеток, сопровождающимся образованием клеточных пакетов.
Эволюция улотриксовых шла в направлении от простейшей формы многоклеточного таллома — нити — к сложной разнонитчатой структуре. В пределах нитчатого типа строения водорослей различают однорядную и многорядную нити, последняя возникает в результате определенного усложнения однорядной нити. Производными многорядной нити являются, с одной стороны, однослойная и двуслойная пластинчатая форма тела, а с другой — трубчато-пузыревидная. Разнонитчатая структура таллома обусловлена совмещением двух форм вегетативного размножения клетки — деления и почкования, а также функциональным расчленением таллома в пределах однорядной нитчатой структуры. Однослойная пластинчатая форма тела могла возникнуть также в результате редукции прямостоячей части разнонитчатого таллома и усиленного развития ползучей по субстрату горизонтальной части или в результате срастания обильно разветвленных в плоскости однорядных нитей.
Ulotrichales широко распространены в природе. Преобладающее большинство обитает в пресных проточных и стоячих водах, значительно меньше видов встречается в щелочных, известковых и кислых водах, а также в солоноватых водоемах и морях. Улотриксовые водоросли преимущественно прикрепленные формы, изредка пассивно плавающие. Подавляющее большинство улотриксовых эпифиты, некоторые — эндофиты (Endoclonium chroolepiforme Szym., Entocladia gracilis Hansg., E. viridis Reinke, Coleochaete nitellarum Jost и др.), сохраняющие самостоятельное автотрофное питание.
Своеобразную экологическую группу представляют собой эндолитофитные организмы, связанные с известковым субстратом. С одной стороны — это сверлящие водоросли. В пресных водах СССР найдены Gomontia codiolifera (Chod.) Wille, внедряющаяся в известковые отложения, G. holdenii Collins, живущая на раковинах перловицы (Unio), G. perforans (Chod.) Acton, сверлящая раковины живых беззубок (Anadonta). Внедряясь в известковый субстрат, эти водоросли делают его рыхлым, легко поддающимся воздействию физических и химических факторов.
Более многочисленна группа туфообразующих водорослей, переводящих растворенные в воде соли кальция в нерастворимые и отлагающих их на поверхности своих талломов, благодаря чему на субстрате возникают мелкие твердые известковые полушаровидные подушечки, которые сливаются в известковые корочки с бугристой или зернистой поверхностью (Fridea torrenticola Schmidle, Chlorotyllium cataractarum Kütz., Ch. mammiforme (Balbis) Kütz., Gongrosira incrustans (Reinsch) Schmidle, G. schmidle P. Rich., G. depauperata Woronich.).
Улотриксовые водоросли развиваются преимущественно в теплый период года, некоторые вегетируют при низкой температуре. Криофильные виды обитают на поверхности слежавшегося снега или в его толще, а также на поверхности льда (виды родов Koliella Hind., Raphidonema Lagerh. em. Hindak).
Незначительная часть улотриксовых водорослей приспособилась к вневодным условиям существования на влажных кирпичных и деревянных стенах, оконных стеклах теплиц, коре хвойных и широколиственных деревьев, полусгнившей древесине, влажных скалах, камнях, железобетонных блоках, а также на влажной почве и непосредственно в почве (Chlorhormidium pseudostichococcus (Heering) Fott, Ch. mucosum (Boye Pet.) Starmach, Hormidiopsis crenulatum (Kütz.) Heering.).
Хозяйственное значение улотриксовых водорослей многообразно и еще не полностью выяснено. Наравне со многими другими группами водорослей они играют определенную роль в процессах естественного самоочищения вод. Некоторые виды, весьма чутко реагирующие на гидрохимические изменения среды, используют как биологические индикаторы для быстрого определения степени загрязненности воды [311]. Четко выраженными мезосапробами являются при высокой их продуктивности Stigeoclonium tenue (Ag.) Kütz., Enteromorpha mtestinalis (L.) Link; к менее выраженным мезосапробам относятся Chlorhormidium subtile (Kütz.) Starmach, Microthamnion kuetzingianum Näg., Chaetophora elegans (Roth) Ag., к олигосапробам — Ulothrix zonata (Web. et Mohr) Kütz,, Draparnaldia glomerate (Vauch.) Ag., D. plumosa (Vauch.) Ag., Coleochaete pulvinata A. Br.
Некоторые улотриксовые водоросли являются постоянными компонентами обрастаний судов и различных гидротехнических сооружений, погруженных в воду. Развиваясь в большом количестве, они способствуют коррозии металлических и бетонированных конструкций, гниению деревянных строений. В почвах улотриксовые участвуют в процессах накопления органических веществ, влияя тем самым на повышение плодородия. В тропических и субтропических областях виды рода Cephaleuros Kunze (Trentepohliaceae) вызывают заболевания культурных цветковых растений. Некоторые виды улотриксовых входят в состав лишайников как автотрофный компонент.
В соответствии с классификационной схемой [221] порядок Ulotrichales включает 4 подпорядка: Ulotrichineae, Ulvineae, Chaetophorineae, Prasiolineae.
Подпорядок 1. Улотриксовые — Ulotrichineae
Талломы нитчатые, простые, легко распадающиеся на фрагменты или отдельные клетки, реже многорядные, не распадающиеся, микроскопические или макроскопические, одноклеточные формы без слизи или объединенные слизью в колонии различной формы. К порядку относится 3 семейства.
Семейство Ulotrichaceae. Нити однорядные, простые, прикрепленные или пассивно плавающие, одноклеточные формы свободноживущие или объединены общей слизью, иногда образуют временные, легко распадающиеся короткие ниточки.
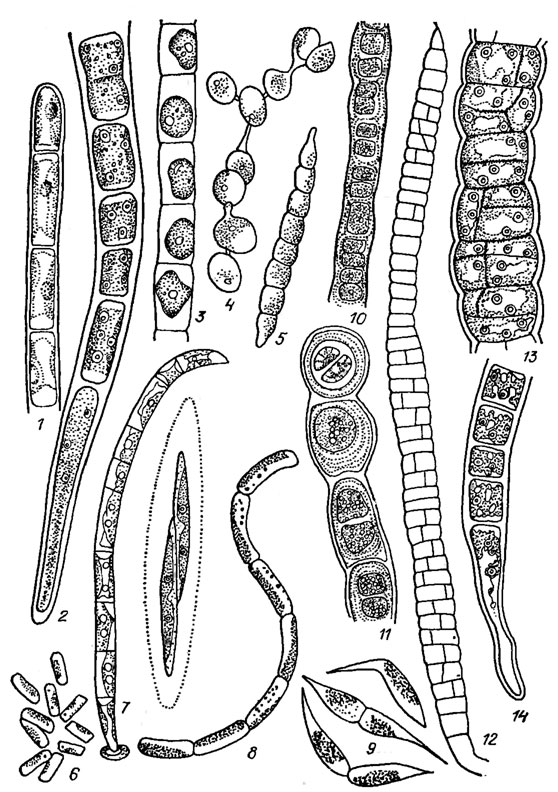
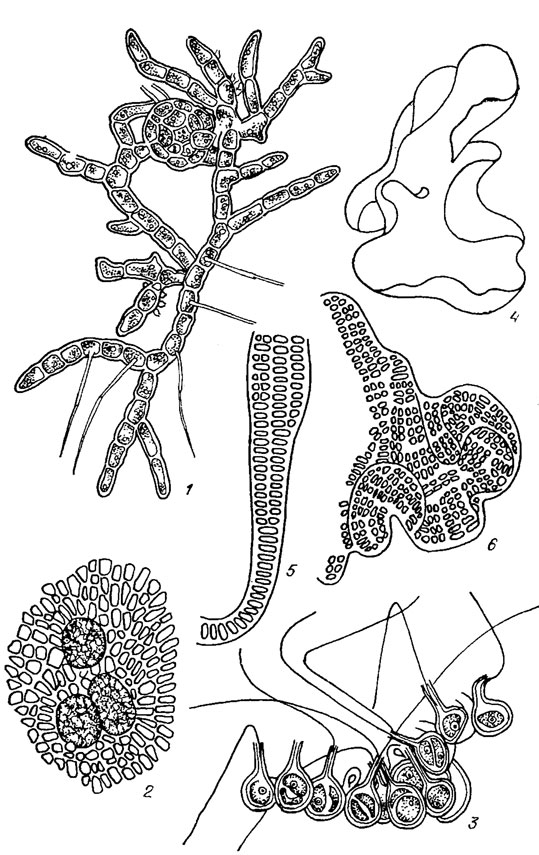
Наиболее распространенные виды представлены на рис. 19.19, 1-9.
Рис. 19.19. Ulotrichales: семейства Ulotrichaceae (1-9), Cylindrocapsaceae Wille (10, 11), Schizomeridaceae (12-14): 1, 2 Ulothrix zonata (Web. et Mohr) Kütz. (l — вершина нити, 2 — основание нити); 3 — Chlorhormidium subtile (Kütz.) Starmach; 4 — Radiofilum paradoxum (Chod. et Topali) Printz; 5 — Raphidonema brevirostre Scherff.; 6 — Stichococcus bacillaris Näg. s. str.; 7 — Uronema intermedia Bourr.; 8 — Gloeotila spiralis Chod.; 9 — Koliella helvetica (Kol) Hindak; 10, 11 — Cylindrocapsa conferta W. West (10 — вегетативная нить, 11 — образование оогониев); 12-14 — Schizomeris leibleinii Kütz. (12 — общий вид таллома, 13 — фрагмент взрослого таллома с утолщенной оболочкой, 14 — базальная часть нити)
Семейство Cylindrocapsaceae. Нити двурядные, прикрепляющиеся к субстрату целлюлозной подошвой; клетки шаровидные или эллипсоидные, с толстыми слоистыми оболочками, часто попарно сближенные; апикальная клетка округлая (см. рис. 19.19, 10, 11).
Семейство Schizomeridaceae. Нити многорядные, прикрепляющиеся к субстрату конусовидной, нередко очень вытянутой базальной клеткой, реже ризоидными выростами; клетки цилиндрические, с тонкими оболочками, плотно соединенные между собой; верхушечная клетка заостренная. Семейство представлено одним видом (см. рис. 19.19, 12-14).
Подпорядок 2. Ульвовые — Ulvineae
Талломы пузыревидные или трубчатые с однослойными стенками и полостью в середине или листовидно расширенные, двуслойные, без полости. К подпорядку относятся 2 семейства.
Семейство Monostromataceae. Таллом гаметофита макроскопический, трубчатый, мешковидный или пластинчатый, всегда однослойный, прикрепляющийся к субстрату подошвой из переплетенных ризоидов или базальным диском. Спорофит одноклеточный (рис. 19.20, 1-7).
Рис. 19.20. Ulotrichales: семейства Monostromataceae (1-7), Ulvaceae (8-14): 1-3 — Blidingia marginata (J. Ag) Dang. (1 — общий вид таллома; 2 — образование веточки; 3 — поперечный разрез таллома); 4, 5 — Monostroma bullosum (Roth) Wittr. (4 — общий вид таллома; 5 — поперечный разрез таллома); 6-7 — Monostroma wittrockii Bornt. (6 — общий вид таллома, 7 — поперечный разрез таллома); 8, 9 — Percursaria percursa (Ag.) Bory (8 — фрагмент двурядной нити, 9 — поперечный разрез двурядной нити); 10-12 — Enteromorpha Intestinaiis (L.) Link (10 — общий вид таллома, 11 — поперечный разрез таллома, 12 — образование веточки); 13, 14 — Ulva rigida Ag. (13 — общий вид таллома; 14 — поперечный разрез таллома)
Семейство Ulvaceae. Таллом макроскопический, нитевидный, простой, из 1-2 рядов клеток, или мешковидный, трубчатый, полый внутри, однослойный, простой или разветвленный, а также пластинчатый, двуслойный, без внутренней полости, на ранних стадиях развития прикрепляющийся ризоидами. Спорофит и гаметофит многоклеточные, макроскопические, сходного строения (см. рис. 19.20. 8-14).
Подпорядок 3. Хетофоровые — Chaetophorineae
Таллом нитчатый или распростерто-пластинчатый, микроскопический или довольно крупный. Нити ветвящиеся, однорядные, часто суженные на концах, обычно прикрепленные, прямостоячие или ползучие, полностью распростертые по субстрату или частично приподнимающиеся над ним. На многих клетках образуются одноклеточные или многоклеточные волоски или щетинки, или они отсутствуют. В составе подпорядка 6 семейств.
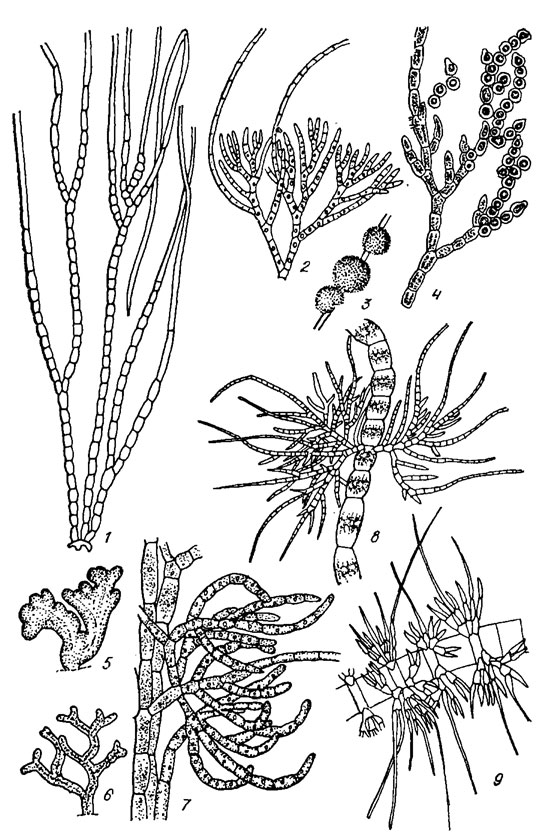
Семейство Chaetophoraceae. Таллом в виде разветвленно-нитевидных, вертикально растущих кустиков, обособленных или объединенных в дерновинки, полушаровидные подушечки и корочки, реже в виде пластинок из горизонтально распростертых нитей. В состав семейства входят 3 подсемейства: Chaetophoroideae (рис. 19.21), Ulvelloideae (рис. 19.22, 1, 2) и Leptosiroideae (см. рис. 19.22, 3-5).
Рис. 19.21. Ulotrichales: семейство Chaetophoraceae, подсемейство Chaetophoroideae: 1 — Stigeoclonium longipillum Kütz.; 2-4 — Chaetophora elegans (Roth) Ag. (2 — форма ветвления, 3 — общий вид таллома, 4 — образование акинет); 5-7 — Chaetophora incrassata (Huds.) Hazen (5, 6 — разные формы талломов. 7 — главные нити с боковой ветвью); 8 — Draparnaldia glomerata (Vauch.) Ag.; 9 — Draparnaldiopsis intermedia Obuch
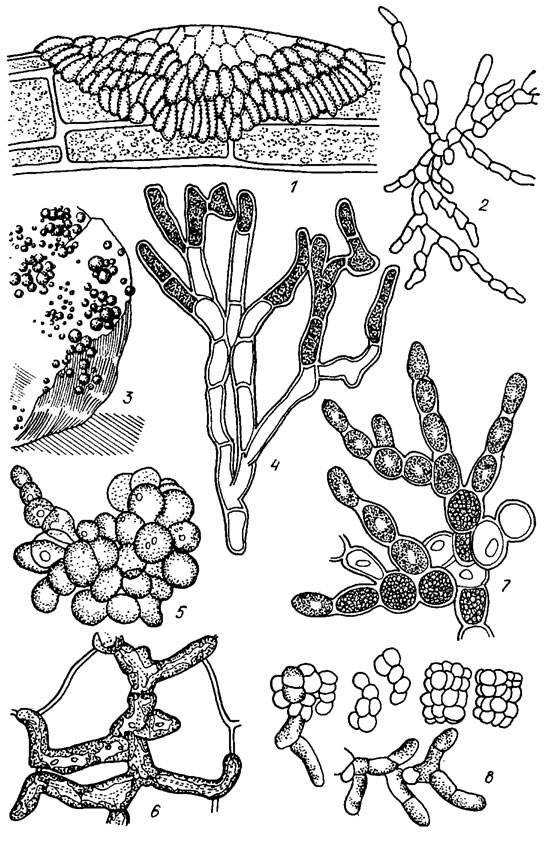
Рис. 19.22. Ulotrichales: семейство Chaetophoraceae, подсемейства Ulvelloideae (1, 2) и Leptosiroideae (3-8): 1 — Pringsheimiella scutata (Reinke) Schmidt; 2 — Entociadia viridis Reinke; 3, 4 — Gongrosira schmidlei P. Richter (3 — общий вид таллома, 4 — фрагмент таллома); 5 — Pleurastrum terrestre Fritsch et John; 6 — Epibolium polysporum During.; 7 — Leptosira mediciana Borzi; 8 — Desmococcus vulgaris (Näg.) Brand
Семейство Trentepohliaceae. Таллом в виде разветвленных, ползучих, приподнимающихся или прямостоячих обособленных нитей, разрастающихся на субстрате в порошковатые налеты или войлочные, подушковидные дерновинки; изредка образуются одноклеточные волосовидные выросты (рис. 19.23, 1-7).
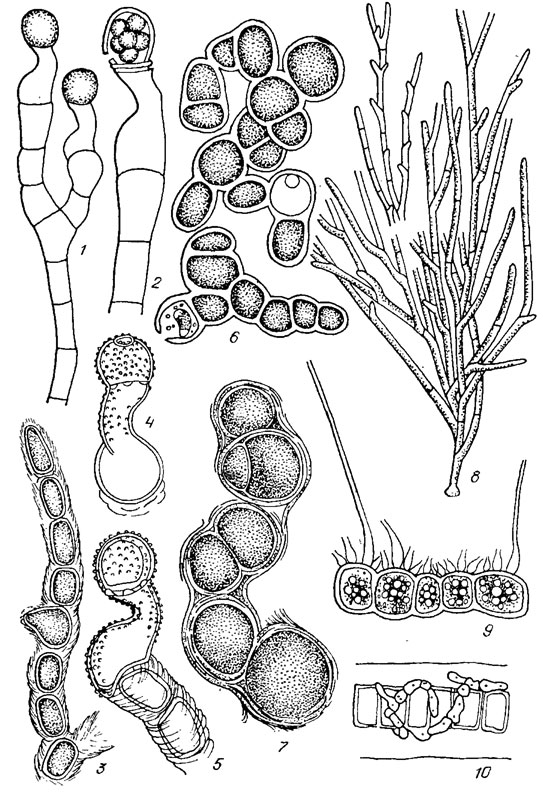
Рис. 19.23. Ulotrichales: семейства Trentepohliaceae (1-7), Microthamniaceae (8), Aphanochaetaceae (9, 10): 1, 2 — Tretitepohlia uncinata (Gobi) Hansg. (1 — фрагмент таллома с зооспор ангиями, 2 — зооспорангий с зооспорами); 3-5 — Trentepohlia jolithus (L.) Wallr. (3 — фрагмент таллома, 4, 5 — зооспорангий на разных стадиях развития); 6, 7 — Trentepohlia umbrina (Kütz.) Born. (6 — общий вид таллома, 7 — фрагмент нити при большом увеличении); 8 — Microthamnion strictissimum Rabench.; 9 — Aphanochaete polychaete (Hansg.) Fritsch; 10 — Aphanochaete vermiculofdes Wolle
Семейство Microthamniaceae. Таллом в виде плотно или рыхло разветвленных прямостоячих кустиков, без слизи, прикрепляющих си базальной клеткой. Волосовидные образования отсутствуют (см. рис. 19.23, 8).
Семейство Aphanochaetaceae. Таллом нитчатый, нити простые или ветвящиеся, обособленные, полностью распростертые по субстрату или частично приподнимающиеся, реже прямостоячие. Волоски одноклеточные, преимущественно вздутые у основания (см. рис. 19.23, 9, 10).
Семейство Coleochaetaceae. Таллом нитевидный, нити ползучие, ветвящиеся, обычно радиально расходящиеся от центральной группы клеток и тесно смыкающиеся боковыми сторонами в однослойную псевдопаренхимную пластинку округлых или неправильных очертаний, реже таллом подушковидный или представлен обособленными, беспорядочно ветвящимися нитями; клетки со щетинками, заключенными в трубчатое влагалище, отогнутое в форме воротничка (рис. 19.24, 1, 2).
Рис. 19.24. Ulotrichales: семейства Coleochaetaceae (1, 2), Chaetosphaeridiaceae (5), Prasiolaceae (4-6): 1 — Coleochaete divergens Pringsh.; 2 — Coleochaete orbicularis Pringsh.; 3 — Chaetosphaeridium pringsheimii Klebahn; 4-6 — Prasiola crispa (Lightf.) Menegh. (4 — общий вид таллома, 5, 6 — фрагменты таллома при большом увеличении)
Семейство Chaetosphaeridiaceae. Таллом многоклеточный, дисковидный, чаще в виде шаровидных или полушаровидных подушечек, прикрепленных к субстрату, реже одноклеточный. Щетинки без трубчатого влагалища либо оно слизистое, цилиндрическое или конусовидное, гомогенное или слоистое (см. рис. 19.24, 3).
Подпорядок 4. Празиоловые — Prasiolineae
Таллом однорядно- и многоряднонитчатый или однослойный, лентовидный или пластинчатый. Хлоропласт центральный, звездчатый.
К подпорядку относится 1 семейство Prasiolaceae, повторяющее признаки подпорядка (см. рис. 19.24, 4-6).