
У человека гаструляция осуществляется следующим способом выберите один или несколько ответов
У человека имплантация погружная, или интерстициальная. За одни сутки бластоциста погружается в эндометрий почти наполовину, а через 40 часов — полностью. Дефект, образовавшийся в слизистой оболочке матки, восстанавливается в течение 5 суток. Условно имплантация состоит из двух фаз — прилипания (адгезии) бластоцисты к слизистой оболочке матки и погружения (инвазии) бластоцисты в глубину слизистой оболочки.
На 6-е сутки эмбриогенеза бластоциста прикрепляется к эпителию эндометрия (обычно, эмбриональным полюсом в области задней или вентральной стенки у маточного угла). В адгезии бластоцисты важную роль играют вещества группы интегринов, вырабатываемые эпителиоцитами слизистой оболочки матки. В норме у женщин между 19-24 сутками менструального цикла — т. е. в самые оптимальные для взаимодействия с бластоцистой сроки, наблюдается активная экспрессия гена интегрина в эпителиоцитах слизистой оболочки матки.
Гаструляция
У зародыша человека гаструляция инициируется в конце 1-й недели развития, сразу по окончании дробления и сброса прозрачной зоны, когда зародыш начинает погружаться в стенку матки.
1-я фаза гаструляция продолжается всю 2-ю неделю развития. Материал внутренней клеточной массы расщепляется способом деламинации на два листка — эпибласт (верхнюю часть) и гипобласт (нижнюю часть). Клетки гипобласта — мелкие кубические, их цитоплазма имеет пенистый вид, они формируют тонкий слой под эпибластом и граничат с полостью бластоцисты. Клетки эпибласта более высокие и имеют вид псевдомногослойного призматического эпителия.
Клетки эпибласта разобщаются посредством небольших полостей, при слиянии которых в дальнейшем формируется амниотическая полость. В нижней части амниотической полости остается небольшая группа клеток эпибласта — материал будущего зародыша (эмбриобласт) и внезародышевых органов.
2-я фаза гаструляции происходит на 3-й неделе развития, осуществляется способом иммиграции и завершается формированием трех зародышевых листков — эктодермы, мезодермы, энтодермы. В будущем из материала этих листков возникнут ткани эмбриона и внезародышевых органов. Происходящие во 2-й фазе гаструляции события напоминают таковые у куриного зародыша и представителей млекопитающих.
Инициация иммиграции связана с активной и неравномерной в разных участках эпибласта пролиферацией клеток, которая приводит к формированию клеточных потоков и образованию на поверхности эпибласта структур, известных как первичная полоска и гензеновский узелок. Округлый и плоский зародышевый диск во 2-ю фазу гаструляции превращается в вытянутый с расширенным краниальным и более узким каудальным концами.
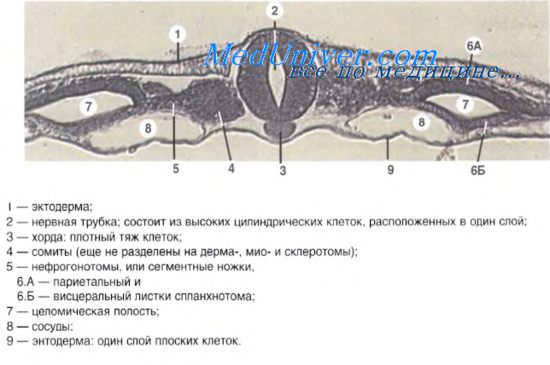
Особое морфогенетическое значение в раннем эмбриогенезе принадлежит головному отростку (нотохорду). Головной отросток растет в краниальном направлении между эпи- и гипобластом, пока не достигнет прехордальной пластинки, которая является «индикатором» будущего рта зародыша. Головной отросток дает начало развитию хорды зародыша — своеобразного клеточного стержня, который определяет первичную ось эмбриона и придает ему «жесткость».
Хорда формирует ось скелета зародыша человека и является основой развития костей осевого скелета (позвоночника, ребер, грудины, черепа). Вокруг хорды в будущем сформируется позвоночный столб. Нотохорд оказывает важное индуцирующее влияние на дифференцировку прилежащего к нему сверху участка эктодермы в нервную пластинку и далее — в нервную трубку, из которой будут развиваться головной и спинной мозг. К концу 3-й недели эмбриогенеза хорда почти полностью сформирована и простирается от орофарингеальной мембраны до каудального конца зародыша.
Источник
Гаструляция книдарий: ключ к пониманию филогенеза или хаос вторичных модификаций?

Данные по эмбриональному развитию книдарий, относящихся к низшим многоклеточным животным, часто используются для решения вопросов, связанных с происхождением и ранней эволюцией Metazoa, а также с основными закономерностями эволюции онтогенеза. Особое внимание уделяется гаструляции – морфогенетическому процессу, в ходе которого клетки раннего эмбриона дифференцируются на зародышевые листки и формируется первичный план строения. В статье проведен сравнительный анализ гаструляции различных книдарий. Мы показывали, что для гаструляционных морфогенезов книдарий характерна высокая степень межгрупповой, внутригрупповой и индивидуальной изменчивости. Мы считаем, что способ гаструляции у книдарий определяется не столько эволюционной историей рассматриваемого вида, сколько эволюционно пластичными адаптивными признаками, такими, как размер яйцеклетки и содержание в ней желтка, число клеток на стадии бластулы или морулы, наличие фототрофных симбионтов, экология личинки. Поскольку онтогенез книдарий обладает высокой эволюционной пластичностью, его изучение должно способствовать лучшему пониманию фундаментальных закономерностей эволюции процессов развития.
У многоклеточных животных (Metazoa) во время эмбрионального развития происходит дифференцировка зародышевых листков: экто- и эндодермы у «низших», «двухслойных» животных, или экто-, эндо- и мезодермы у «высших», трёхслойных животных. Зародышевые листки дифференцируются в ходе гаструляции — процесса, во время которого также закладывается первичный план строения организма. Считается, что изучение гаструляции «низших» Metazoa является ключом к пониманию происхождения и ранней эволюции животных, и особенно закономерностей эволюции их онтогенеза. Достаточно вспомнить теории гастреи Э. Геккеля и фагоцителлы И.И. Мечникова, в которых морфология эмбриона на стадии гаструлы рассматривается как рекапитуляция (воспроизведение) одного из самых ранних этапов эволюционного становления Metazoa. Данные по эмбриональному развитию «низших» Metazoa широко используются в настоящее время для подтверждения или опровержения филогенетических построений и макроэволюционных гипотез. Неформальная группа «низших Metazoa» объединяет типы Cnidaria (стрекающие или книдарии), Porifera (губки), Placozoa и Ctenophora (гребневики). Предполагается, что книдарии являются сестринской группой по отношению к билатерально-симетричным животным (Bilateria).
Как типичные представители низших Metazoa, книдарии обладают простым планом строения, который принято считать эволюционно примитивным (рис. 1). Они двухслойны: их тело состоит из экто- и эндодермы. Книдарии имеют единственную орально-аборальную ось тела, кишечную полость, открывающуюся наружу только одним (ротовым) отверстием. К числу признаков, характерных для типа Cnidaria, можно отнести также разнообразие способов бесполого размножения, высокую способность к регенерации, а также к тканевым и клеточным трансдифференцировкам (способность взрослой региональной стволовой клетки дифференцироваться в клетки другого органа и/или другого зародышевого листка). В настоящее время в типе Cnidaria выделяют пять классов: Anthozoa, Scyphozoa, Staurozoa, Hydrozoa и Cubozoa (рис. 2). Их филогенетические взаимоотношения остаются спорными.

Несмотря на прогресс в изучении развития книдарий современными методами (в том числе методами молекулярной генетики и биоинформатики), наши знания об их эмбриональном развитии, к сожалению, часто остаются на уровне конца XIX века. Могут ли имеющиеся у нас данные по раннему развитию книдарий прояснить спорные моменты их эволюции и филогении? В статье «Гаструляция книдарий: ключ к пониманию филогенеза или хаос вторичных модификаций?» авторы отвечают на этот вопрос, анализируют накопленные к настоящему моменту сведения об эмбриональном развитии, и в первую очередь, о гаструляции различных представителей типа Cnidaria.
У книдарий существует четкая преемственность между орально-аборальной полярностью взрослого животного и полярностью яйцеклетки. Оральный полюс полипа соответствует анимальному полюсу яйца, на котором происходит отделение полярных телец, расположен женский пронуклеус и закладывается первая борозда дробления. У видов, гаструляция которых морфологически поляризована, этот полюс также является полюсом формирования эндодермы. У активно плавающей личинки планулы этот полюс становится задним концом. Из заднего конца планулы в ходе метаморфоза формируется оральный полюс полипа.
Первичный план строения Cnidaria формируется при помощи широкого спектра различающихся путей развития. Эти пути эквифинальны, так как у абсолютного большинства книдарий они приводят к одному и тому же итогу: формированию двухслойной личинки планулы (или паренхимулы, а затем планулы).
Изучение эмбрионального развития, и особенно гаструляции Cnidaria имеет очень длинную историю. Фактически, классификация типов гаструляции и первые описания многих гаструляционных морфогенезов, сделанные в XIX веке, были основаны на изучении эмбриогенеза книдарий (Metschnikoff, 1886). В тот же период данные по развитию книдарий и других низших Metazoa (губок) начинают использоваться для построения теорий происхождения многоклеточности и онтогенеза животных (Metschnikoff, 1886; Haeckel, 1874).
Одна из самых удачных классификаций типов гаструляции (гаструляционных морфогенезов) была предложена О.М. Ивановой-Казас (1975, с. 50). Эта классификация подчёркивает тесную связь типов гаструляции с морфологией бластулы, которая, в свою очередь, зависит от особенностей дробления. В статье приводится её упрощённый вариант (рис. 3).

Итак, если в результате дробления получается целобластула (рис. 3, а), гаструляция может осуществляться за счёт первичной (клеточной) деламинации (рис. 3, б), униполярной иммиграции клеток (рис. 3, в), мультиполярной иммиграции клеток (рис. 3, г) и инвагинации (рис. 3, д). Если в результате дробления получается морула (плотная группа клеток) (рис. 3, е), гаструляция может осуществляться за счёт вторичной (морульной) деламинации (рис. 3, ж). Если в ходе дробления получается бластула, состоящая из клеток, резко различающихся по размеру (амфибластула, рис. 3, з), то гаструляция скорее всего будет осуществляться за счёт эпиболии (рис. 3и). Существует также довольно редкий вариант гаструляции – изгибание плакулы (рис. 3, к, л). Если целобластула по каким-то причинам имеет плоскую (дисковидную) форму, то ее можно называть плакулой (рис. 3, к), а гаструляция представляет собой последовательное преобразование этого диска в чашу и сферу с отверстием – бластопором (рис. 3, л). Интересно, что у книдарий обнаружены практически все варианты гаструляционных морфогенезов, приведенные на этом рисунке. При таком разнообразии уместно задаться вопросом, существует ли строгая связь между типом гаструляции и филогенетическим положением тех книдарий, для которых он характерен. Попробуем выяснить так ли это, рассмотрев особенности гаструляционных морфогенезов у представителей разных таксонов книдарий. В статье рассматриваются гаструляция 4-х классов книдарий — Anthozoa, Scyphozoa, Staurozoa и Hydrozoa (информации по раннему развитию Cubozoa в литературе отсутствует). Однако здесь мы остановимся (в качестве примера) на анализе данных по гаструляции Anthozoa и Hydrozoa,
Гаструляция у Anthozoa
В классе Anthozoa эмбриогенез лучше всего изучен у подкласса Hexacorallia. За последние два десятилетия было подробно изучено развитие модельных видов: актинии Nematostella vectensis и склерактинии Acropora millepora . Развитие немодельных склерактиний сейчас очень активно изучает эмбриолог Нами Окубо (Токийский Университет). Наш анализ развития этой группы в значительной степени опирается на её данные (Okubo et al., 2013). Обобщив всю имеющуюся в литературе информацию, мы выделили пять основных вариантов путей развития Hexacorallia (рис. 4, а-д):

Насколько жестко тот или иной путь развития связан с таксономическим положением вида? Самое необычное развитие, при котором формируется плакула (вариант 5), наблюдается только у некоторых родов склерактиний. Однако при более внимательном анализе их развития выясняется, что эмбрион на прегаструляционной стадии далеко не всегда может быть с полным основанием описан как плакула: очень часто полость бластоцеля бывает хорошо выражена. Полностью соответствует варианту 5 развитие представителей лишь нескольких родов, например, рода Acropora . У других родов склерактиний рот формируется непосредственно из отверстия бластопора, а клетки архентерона (гастрального впячивания) не деэпителизуются.
Судя по всему, эмбрионы разных видов Hexacorallia можно выстроить в ряд постепенных переходов – от целобастулы до плакулы. Так, эмбрион актинии Nematostella приобретает форму изогнутой плакулы или чаши несколько раз на протяжении стадии «пульсирующей бластулы». Мы предполагаем, что уплощенная или чашеобразная форма ранних эмбрионов актиний и склерактиний может быть временной, как у Nematostella , и эмбрионы принимают ее несколько раз до начала гаструляции. Насколько это предположение справедливо, можно будет сказать только после более детального исследования эмбриогенеза представителей этих кораллов методом. Если оно справедливо, то вариант развития 3 может быть полностью сведен к варианту 1.
Таким образом, у проанализированных таксонов Hexacorallia внутригрупповое разнообразие путей развития сопоставимо с межгрупповым. Значительная часть этого разнообразия связана с накоплением в яйцеклетках желтка, затрудняющего морфогенетические движения, а также с необходимостью транспортировки симбионтов в клетки эндодермы. При этом разнообразие самих гаструляционных морфогенезов невелико.
Гаструляция у Hydrozoa
По данными молекулярной филогении, Hydrozoa – самая молодая группа книдарий. Именно в процессе изучения этой группы были открыты все основные гаструляционные морфогенезы, кроме инвагинации. В работе, посвященной эмбриональному развитию книдарий, Мечников (Metschnikoff, 1886) предложил разделять гаструляционные морфогенезы на гипотропные (т.е. униполярные, поляризованные) и мультиполярные (аполярные) по отношению к орально-аборальной оси книдарий. К поляризованным морфогенезам относятся униполярная иммиграция, инвагинация и эпиболия, а к мультиполярным – первичная (клеточная) деламинация, вторичная (морульная) деламинация, смешанная деламинация (комбинация первичной и вторичной деламинации) и мультиполярная иммиграция (см. рис. 3). Нужно подчеркнуть, что Мечников не рассматривал эти морфогенезы как дискретные варианты («Некоторые из этих способов нельзя резко разграничить, так как они связаны переходами»).

Для многих гидроидов характерна стадия морулы (рис. 3е; 5а, в, ж, з). Морфология морулы у разных видов заметно различается. Иногда морула представляет собой рыхлый агрегат клеток (рис. 5а). В этом случае гаструляция осуществляется за счет своеобразной формы вторичной деламинации. Для стадии гаструлы тогда характерны неправильная форма эмбриона, а также высокий уровень морфологической и морфогенетической изменчивости (рис. 5, д), который резко снижается при переходе к стадии паренхимулы (рис. 5м). Морула может быть плотной, и тогда вторичная деламинация, не связана с существенным изменением формы эмбриона вплоть до стадии паренхимулы (рис. 5в, з). Из паренхимулы, как правило, формируется личинка — планула, у которой эпителизована не только эктодерма (как у паренхимулы), но и эндодерма (рис. 5н).
Морула также может формироваться в процессе первичной деламинации. Так протекает развитие у гидроидов, которые формируют бластулу со слабо выраженным бластоцелем (рис. 5, б). Благодаря ориентации митотических веретен перпендикулярно поверхности эмбриона дочерние клетки отшнуровываются в полость бластоцеля (рис. 5, е, ж). В результате получается плотная морула (рис. 5з). Окончательное обособление эктодермы от эндодермы осуществляется за счет вторичной деламинации.
У других гидроидов первичная деламинация начинается на относительно поздней стадии. В этом случае успевает сформироваться бластула с хорошо выраженной полостью (рис. 5, г, и), которая постепенно также превращается в плотную морулу (рис. 5, з).
Если в ходе дробления получается бластула, гаструляция может происходить за счет униполярной или мультиполярной иммиграции (рис. 5, к, л).
Униполярная иммиграция — очень распространенный морфогенез. Он связан с униполярным формированием колбовидных клеток, которые постепенно выселяются в бластоцель (рис. 5, к). Этот процесс приводит к формированию паренхимулы (рис. 5, м). Мультиполярная иммиграция, напротив, описана для очень небольшого числа видов гидроидов.
Инвагинационный мофогенез не найден до сих пор ни у одного гидроида. Скорее всего, отсутствие инвагинации среди гаструляционных морфогенезов можно считать признаком, характерным для класса Hydrozoa.
Итак, гаструляционные морфогенезы Hydrozoa очень разнообразны. Еще в XIX веке предпринимались попытки найти закономерности, объясняющие их распространение среди представителей этого класса. Мечников обратил внимание на связь способа гастуляции с экологией эмбриональных стадий. Судя по всему, эмбрионы, ведущие подвижный образ жизни (становящиеся личинками на стадии бластулы), формируют эндодерму с помощью униполярной иммиграции. С другой стороны, эмбрионы, развивающиеся до стадии планулы в материнском организме или лежащие на дне используют мультиполярные морфогенезы (Metschnikoff, 1886). При внимательном анализе большего числа видов обнаруживается, что представители близких родов могут использовать совершенно разные способы гаструляции. Учитывая, что уже описаны виды гидроидов с индивидуально изменчивой гаструляцией, можно сделать вывод, что в классе Hydrozoa разнообразие гаструляционных морфогенезов связано скорее с экологией эмбрионального развития, чем с таксономическим положением вида, причем внутригрупповое разнообразие не уступает межгрупповому.
Заключение
Наш анализ гаструляции Cnidaria показал, что не существует дискретных, четко различающихся путей развития. Мы увидели, что разные варианты гаструляционных морфогенезов распределены по классам и отрядам книдарий в значительной степени мозаично; четкой приуроченности к тем или иным таксонам, как правило, не наблюдается. Скорее следует говорить о существовании непрерывной изменчивости способов развития.
Судя по всему, способ гаструляции у книдарий часто менялся в ходе эволюции. Он подвержен значительной внутригрупповой изменчивости, порой не уступающей межгрупповой. Возможно, способ гаструляции книдарий определяется не столько эволюционной историей рассматриваемого вида и его таксономической принадлежностью, сколько эволюционно пластичными адаптивными признаками, такими как размер яйцеклетки и содержание в ней желтка, число клеток бластулы, наличие фототрофных симбионтов, экология личинки. Более того, ход гаструляции подвержен внутривидовой (индивидуальной) изменчивости. Из этого следует, что гаструляционные морфогенезы книдарий могут иметь лишь весьма ограниченное применение при решении частных вопросов филогенетики. С другой стороны, сравнительное изучение онтогенеза низших Metazoa проливает свет на общие закономерности морфогенетических процессов и их эволюции, что крайне важно для понимания ранних этапов становления животного царства.
Источник



