
- БИОЛОГИЯ РАЗМНОЖЕНИЯ РАСТЕНИЙ — Н. Л. Колясникова — 2017
- РАЗДЕЛ 3. ТИПЫ ОПЫЛЕНИЯ
- 3.1 Самоопыление
- 3.2 Перекрестное опыление
- Опыление — виды, способы и роль процесса в жизни растений
- Основные термины
- Энтомофилия — работают бабочки
- Зоофилия — не обойтись без лемура
- Искусственный процесс
- Анемофилия и гидрофилия
- Описание механизма
- Примеры в окружающей природе
БИОЛОГИЯ РАЗМНОЖЕНИЯ РАСТЕНИЙ — Н. Л. Колясникова — 2017
РАЗДЕЛ 3. ТИПЫ ОПЫЛЕНИЯ
Опыление — процесс переноса пыльцы на рыльце пестика. Различают два типа опыления: самоопыление и перекрестное опыление. У покрытосеменных растений преобладает перекрестное опыление. Реже встречается самоопыление. Постоянное самоопыление считают тупиком эволюции, ведущим к деградации.
3.1 Самоопыление
При самоопылении пыльца одного растения попадает на рыльце пестика этого же растения. Этот тип свойственен только обоеполым цветкам.
Различают приспособления к самоопылению: автогамия, гетоногамия, клейстогамия.
Автогамия осуществляется по-разному: при прямом соприкосновении — контактная автогамия, при высыпании пыльцы и оседании под собственным весом — гравитационная автогамия, под действием ветра — ветроавтогамия, с помощью мелких насекомых — трипсоавтогамия. Контактная автогамия характерна для седмичника, майника, копытня, гравитационная описана у одноцветки, ветроавтогамия встречается у голубики.
Гейтоногамия — это соседственное опыление, когда пыльца с одного цветка соцветия прорастает на рыльце пестика другого цветка этого же соцветия. Оно может происходить в конце сезона цветения. Ей способствует ветер, животные, насекомые. Встречается данный способ опыления у видов сем. Астровые, Капустные. Растения с таким способом опыления характеризуются мелкими цветками в густых соцветиях.
Клейстогамия — опыление в закрытом цветке. Различают облигатную и факультативную клейстогамию. Клейстогамные цветки мельче по размерам, части цветка редуцированы. Лепестки рудиментарны или отсутствуют. Число тычинок уменьшено, пыльцевые зерна мельче. Примеры растений — фиалка удивительная, кислица обыкновенная (рис. 20, 21).
Рис. 20. Двойное оплодотворение в клейстогамных цветках кислицы обыкновенной: 1 — опустошенный пыльник клейстогамного цветка после прорастания пыльцевых трубок, остались оболочки пыльцевых зерен, пыльцевые трубки и дегенерировавшие пыльцевые зерна; 2 — проникновение пыльцевой трубки между клетками стенки

Рис. 21. Пыльцевые трубки в пыльнике клейстогамного цветка у фиалки кочкарной — 1; прорастание пыльцевых трубок через разрыв в стенке пыльника клейстогамного цветка у фиалки кочкарной — 2

Помимо клейстогамных цветков у этих растений образуются и хазмогамные цветки с открытым цветением. Факультативная клейстогамия не постоянна, возникает при определенных условиях. У них нет признаков редукции в цветке. Она связана с неблагоприятными факторами среды, встречается у ячменя, овса, пшеницы, ковыля и др.
3.2 Перекрестное опыление
При перекрестном опылении пыльца с цветков одного растения попадает на рыльца пестиков других растений данного вида.
Различают следующие приспособления к перекрестному опылению: двудомность, дихогамия, самонесовместимость, суточный ритм цветения.
Двудомность — разделение мужских и женских цветков по разным растениям одного вида. К двудомным видам относятся: облепиха, ива, тополь, осина, щавель, крапива, спаржа и др. Адаптивное значение разделения полов заключается в обеспечении перекрестного опыления и в дифференциации половых форм по экологическим нишам. По фенологии первыми зацветают мужские растения. Длительность цветения женских растений дольше. Тычиночные цветки крупнее пестичных. Число мужских цветков в соцветии и на растении в целом больше, чем женских.
Дихогамия — разновременное созревание пыльников и рылец. Это функциональная раздельнополость. Цветок выступает то в мужской, то в женской фазе. У разных видов встречаются разные типы: сначала мужская стадия — протерандрия, или сначала женская — протерогиния.
Протерандрия встречается часто. Широко представлена у растений класса двудольные. Иногда тычиночная и пестичная стадии четко разграничены по времени. Рыльца становятся восприимчивыми к пыльце, когда все тычинки данного цветка завяли.
Протерогиния встречается у растений класса однодольные и примитивных двудольных (сем. Лютиковые).
Самонесовместимость — это неспособность пыльцевых трубок прорастать в столбик на всю его длину и совершать оплодотворение. Гены, ответственные за самонесовместимость обозначаются S. Если пыльца и рыльце несут одинаковые аллели S-гена, то оплодотворение не происходит. Различают гетероморфную и гомоморфную самонесовместимость. Гетероморфная делится на ди- и тристилию. При гомоморфной самонесовместимости цветки одинаковы по строению. Гомоморфная самонесовместимость встречается чаще. Различают два генетических контроля: спорофитный и гамето- фитный. При спорофитной несовместимости способность пыльцы к прорастанию определяется генотипом спорофита, то есть растения, на котором идет образование микроспор. При спорофитном контроле подавление прорастания пыльцы происходит рано, уже на рыльце. Поверхность рылец сухая, покрыта пелликулой. Столбик закрытый. Пыльцевые зерна трехядерные. Это тип самонесовместимости встречается у растений сем. Астровые, Капустные, Гвоздичные и др.
При гаметофитной несовместимости способность к прорастанию пыльцы определяется ее генотипом (то есть генотипом самого гаметофита) (рис. 22).
Рис. 22. Рост пыльцевых трубок и оплодотворение при гаметофитной несовместимости

При гаметофитном контроле пыльцевые трубки сначала растут быстро, затем рост их тормозится и прекращается. Рыльца влажные, столбик — открытый. Пыльцевые зерна двуядерные. Гаметофитная самонесовместимость отмечена у растений сем. Бобовые, Лилейные, Розоцветные, Пасленовые и др.
Дистилия — разностолбчатость. Впервые описана у примулы. Встречаются два типа цветков: длинностолбчатые и короткостолбчатые. Для них характерен диморфизм пыльцы: у короткостолбчатых растений пыльца крупнее, сосочки рыльца короче. Легитимным (законным) является опыление пыльцой с коротких тычинок на рыльце короткого столбика и, наоборот, пыльцой с длинных тычинок на рыльце длинного пестика. Также дистилия детально изучена у гречихи (рис. 23).
Рис. 23. Дистилия у гречихи: а — короткостолбчатая; б — длинностолбчатая форма

При тристилии (дербенник) три типа цветков — короткие столбики и длинные и средние тычинки, средние столбики и длинные и короткие тычинки, длинные пестики и средние и короткие тычинки.
Дистилия контролируется одним супергеном: генотип растений с короткостолбчатыми цветками SS и Ss, а растений с длинностолбчатыми цветками — ss. Между цветками одного морфотипа само- и перекрестное оплодотворение не осуществляется (иллегитимный вариант опыления), но оно возможно между растениями разных морфотипов (легитимный вариант опыления). При тристилии генетическое строение различных форм основано на двух локусах М и S: длинностолбчатые цветки: mmss, среднестолбчатые: Мmss или ММss,короткостолбчатые: МmSs, mmSs, ММSs, ММSS, mmSS.
Псевдосовместимость. У многих растений появление псевдосовместимости связано с искусственным опылением. На уровень псевдосовместимости влияют свет, температура, влажность. Существует псевдосовместимость «конца сезона», или при опылении бутонов, при удалении верхушки столбика, при использовании растительных гормонов.
Суточный ритм цветения. Большинство цветков реагируют на смену дня и ночи. Некоторые цветки открываются и закрываются несколько раз. Другие распускаются лишь один раз. В зависимости от приуроченности раскрывания цветков к определенному суточному периоду различают растения с утренним (одуванчик), дневным (люцерна), вечерним (матиолла), ночным (тропические растения), круглосуточным (брусника) и порционным взрывчатым (сем. Мятликовые) цветением. Суточный ритм распускания цветков рассматривается рядом ученых как проявление биоритмов.
Методика наблюдений за суточным ходом цветения растений предложена А.Н. Пономаревым. Суточную динамику распускания цветков определяют в 50 соцветиях исследуемого вида в течение суток в трехкратной повторности. Через каждые два часа подсчитывается число вновь распустившихся цветков. При этом необходимо регистрировать температуру и относительную влажность воздуха (рис. 24).
Рис. 24. Суточный ход распускания цветков Astragalus onobrychis 30.06.1992 г.: температура, °С, — влажность, — число цветков

Различают два способа перекрестного опыления: биотическое и абиотическое.
Биотическое опыление. Опыление первоначально осуществлялось жуками. Они питались пыльцой. Затем с появлением нектарников главными опылителями стали перепончатокрылые, двукрылые, чешуекрылые. Таким образом, пыльца и нектар стали первичными аттрактантами. Пыльца содержит белки, жиры, углеводы, ферменты, витамины. У энтомофильных растений в пыльцевых зернах больше жиров и белка, у анемофильных — углеводов. Пыльца относительно долговечна. Нектар — это водный раствор сахаров: фруктозы, сахарозы и др. Также нектар включает аминокислоты, белки, органические кислоты, витамины и др. Концентрация сахара достигает 74% (у конского каштана). Выделение нектара зависит от времени дня, возраста и стадии развития цветка. Нектарники встречаются на разных органах цветка — на чашелистике, лепестках, цветоложе, тычиночных нитях.
Для привлечения опылителей помимо пищи служат вторичные аттрактанты: окраска и запах. Окраска обусловлена пигментами: флавоноидами, каротиноидами, антоцианом, антофеином. Часто окраска неоднородна, имеет узор или рисунок. Он служит указателем пути к нектару. У некоторых растений в течение жизни наблюдается изменение окраски, что связано с изменением рН клеточного сока вакуолей в клетках лепестков после осуществившегося оплодотворения. Например, у медуницы цветки сперва розово-пурпурные, затем — синие. Это сигнал для опылителей, что пыльцы нет.
Существует предпочтение окраски для разных опылителей. Пчелы не различают красный цвет, но восприимчивы к ультрафиолету. При одновременном цветении растений с синими и желтыми венчиками, пчелы в первую очередь опыляют синие цветки. В географическом плане тоже есть дифференциация по окраске. В тропиках преобладают красные цветки, в высокогорьях — белые. Желтые цветки — в пустынях и в Приполярье. Синие цветки — на равнинах в умеренном климате.
Аромат энтомофильных цветков разнообразен. Различают более 500 запахов. Изредка это трупный запах гниения у аронников, рафлезий. Иногда запах обусловлен аминами (боярышник, рябина). Чаще запах связан с эфирными маслами. Источник запаха — лепестки, тычинки, стаминодии, нектарники, пыльца. Интенсивность запаха колеблется и достигает максимума в определенное время суток. Например, петуния — ночью, клевер — днем.
Характеристика способов биотического опыления представлена в таблице 4.
Источник
Опыление — виды, способы и роль процесса в жизни растений

Основные термины
С определения «опыление — это…» уже в 3 классе школьники начинают знакомство с этапами полового размножения семенных представителей флоры. Процесс подробно описывает, как происходит опыление у растений: во время цветения пыльца покидает пыльник — орган формирования микроспор, находящийся в тычинке, и улавливается верхней частью пестика — рыльцем, что характерно для покрытосеменных растений. Голосеменные используют для этого семяпочку, расположенную на поверхности семенных чешуек.
Природа разумно распределила роли: тычинки являются мужскими органами, пестики и семяпочки — женскими. Новая жизнь в виде семени образуется при условии удачного захвата пыльцы и своевременного оплодотворения. Выделяют два основных типа опыления растений:
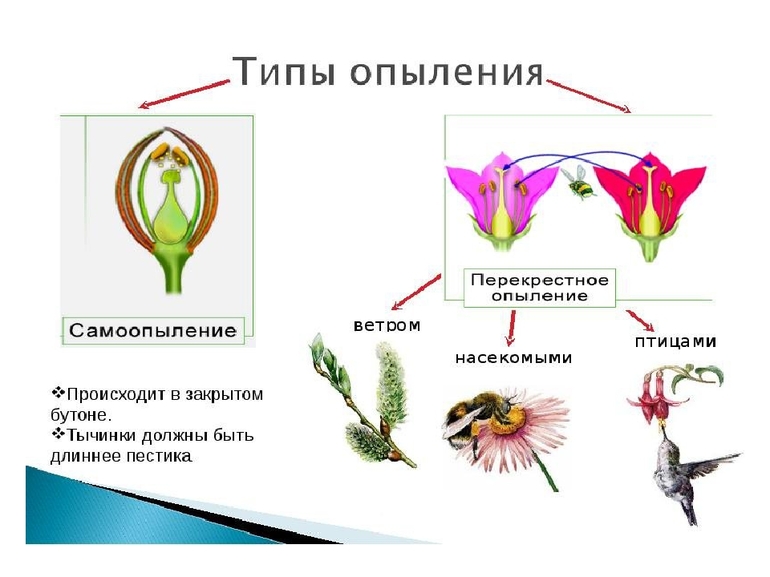
- Во всех случаях, когда цветки свободно опыляются своей пыльцой, говорят об идиогамии или самоопылении. Вариантом считается форма гейтоногамии — соседственного опыления, когда в пределах одного растения пыльца на рыльце попадает от разных цветков той же особи. К самоопылителям относятся ячмень, пшеница, овёс, горох и просо — их бутоны не раскрываются до конца, но даже мелкие и невзрачные цветки дают полноценные семена. Самоопыление на постоянной основе считается тупиком развития, когда процессы микроэволюции затухают, а вид расщепляется на чистые линии.
- Участие специального посредника, который доставляет пыльцевые зёрна от тычинок к рыльцам пестиков между цветками различных особей, делает опыление перекрёстным или ксеногамией. Это основной тип размножения для однодомных и двудомных растений: в соцветиях обнаруживается разделение полов или в популяции равноправно сосуществуют одно- и двуполые цветки.
Обмен генами и интеграция возможных мутаций помогают виду лучше приспосабливаться к окружающей среде, поддерживают высокий уровень жизнеспособности популяции, отвечают за целостность и единство сообщества. Создаётся широкое поле деятельности для естественного отбора. Положительные и отрицательные стороны обоих типов приводятся в таблице:
| Наименование | Достоинства | Недостатки | Примеры |
| Самоопыление | Адаптация к неблагоприятным условиям. Позволяет оперативно восстановить популяцию однолетних растений | Малое количество семян. Слабое потомство | Кукуруза, пшеница, фиалка |
| Перекрёстное опыление | Разнообразие способов переноса пыльцы. Яркая окраска и сильный запах цветов | Зависимость от внешних факторов. Исчезновение опылителей сокращает популяцию. Особые требования к расположению и форме цветков | Маргаритка, лютик |
Если перекрёстное опыление происходит при помощи живых организмов, его считают биотическим, а насекомые или животные, участвующие в нём, носят название агентов опыления или опылителей. Биотическими считаются энтомофилия, зоофилия и искусственное опыление, во всех остальных случаях речь идёт об абиотических процессах — анемофилии и гидрофилии.
Энтомофилия — работают бабочки

Опылителями выступают насекомые: осы и пчёлы из улья, реже — муравьи, жуки, улитки, бабочки и мухи. Для этого двуполые растения стараются их привлекать: запасаются сладким нектаром, берут на вооружение приятные запахи, кричащую окраску и крупные размеры соцветий. Насекомые легко переносят липкую пыльцу с одного цветка на другой, обеспечивают продолжение рода калины и гречихи, ромашки и шалфея, молочая и мака.
Цветочная пыльца крупнодисперсная, клейкая и надёжно прилипает к маленьким телам и конечностям. Отдельные виды растений «помогают» процессу — обладают удобной чашеобразной формой соцветия. Попавшее в этот кратер насекомое буквально обваливается в пыльце, а качество опыления возрастает.
Зоофилия — не обойтись без лемура

Процесс происходит при активном участии животных, относящихся к группе позвоночных. Различают орнитофилию, где роль агентов опыления играют птицы (нектарницы, близкие родственницы воробьёв, уникальные вертолётики колибри или медососы с особыми щёточками на языке) и хироптерофилию, когда за дело берутся насекомоядные летучие мыши, мелкие обезьяны, австралийские сумчатые грызуны или мадагаскарские лемуры. Зоофилией характеризуются тропические растения с яркими призывно окрашенными цветами.
Искусственный процесс
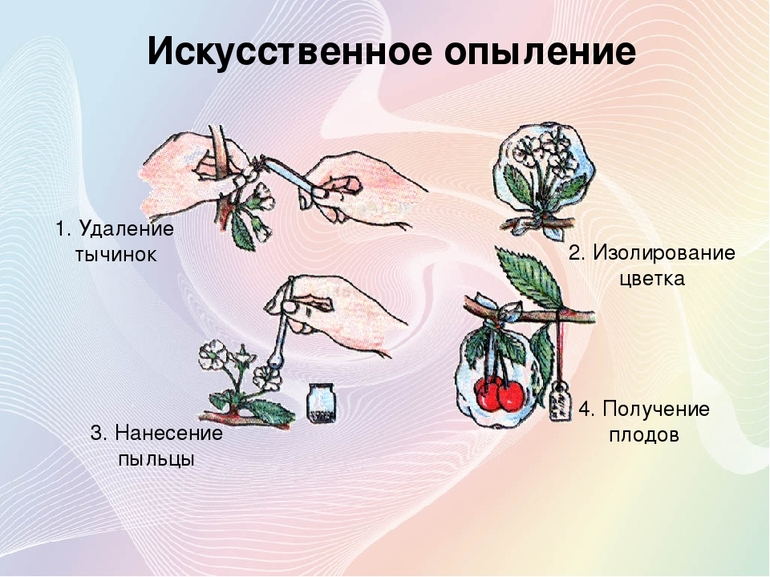
Для нужд овощеводства, плодового и декоративного садоводства или лесного хозяйства пыльцу с тычинок на пестики переносят специально обученные люди. Чтобы не отстать от матери-природы, они используют различные приспособления. Эти агротехнические приёмы широко применяются для скрещивания и получения новых видов и сортов растений.
Простейший пример — дополнительное опыление ржи: два человека хватаются за концы верёвки, растягивают их и движутся по ржаному полю, касаясь цветущих растений. От сотрясения пыльца осыпается с тычинок и легко оказывается на рыльцах пестиков. Существенно повысить урожайность позволяет обработка порядка 10% цветков, проведённая в первой половине сухого солнечного дня при температуре не выше +30 °C.
Анемофилия и гидрофилия

Опыление при помощи ветра широко распространено среди злаковых, хвойных и лиственных растений. Например, скромные цветки крапивы или конопли складываются из чашелистиков, обладают простыми околоцветниками, поэтому непривлекательны для опылителей. Зато они имеют достаточное количество сухой и мелкой пыльцы, а пыльники, подвешенные на тонких удлинённых нитях, легко выбрасывают её наружу. Берёзовая и ольховая, ореховая и тополиная пыльца будет лучше попадать на рыльце пестика, если он станет широким и длинным, да ещё и высунется из цветка, как в знаменитом соцветии «серёжка».
Гидрофилия — опыление с участием воды, а иногда ему на помощь приходят и слизни. Немногочисленные гидрофильные растения (элодея и взморник, наяда и руппия) имеют нитевидную пыльцу или рыльца, раскрывают пыльники под водой, где и создаются наилучшие условия для продолжения рода.
Значение опыления нельзя переоценить: ему отводится важнейшая роль в размножении любых цветковых растений — от малых причудливых форм до гигантских деревьев. До 80% представителей биологического царства относятся к биотическому типу опыления, а из оставшихся двадцати около 19% опыляются ветром.
Описание механизма
Цветковые растения получили своё название, потому что имеют морфологический орган — цветок, способный к половому размножению и привлечению опылителей. Для образования завязей, формирования плодов и развития семян первым условием является опыление, то есть перенос частиц пыльцы. Природа позаботилась о том, чтобы всё проходило успешно.
Это может осуществляться ветром — тогда целые облака невесомых пылинок поднимаются и передвигаются в потоках зефира. Многие деревья (дубы, ясени и сосны), сельскохозяйственные злаковые и кукуруза «заботятся» об увеличении вероятности попадания на нужные рыльца, вырабатывая заведомо большие количества пыльцы. Её частички очень лёгкие, почти невесомые, чтобы беспрепятственно «проплывать» по воздуху, а у некоторых имеются газовые пузырьки, способствующие более долгому путешествию.
Но чаще всего опыление осуществляют разные насекомые. Чтобы привлечь внимание, растения окрашиваются в очень яркие цвета и источают сильный аромат. Если их размеры малы, они группируются в пышные соцветия или окружают себя разноцветными листьями — прицветником, как это делает мексиканская красавица пуансеттия. По сравнению с теми, что опыляются ветром, пыльцевые зёрна таких цветов обычно более крупные, шероховатые и клейкие, чтобы уверенно прилипать к насекомым.
Для привлечения опылителей медовые железы растений специально выделяют нектар — сок, богатый различными сахарами (в основном это сахароза и фруктоза). Кроме того, в состав входят:
- кислоты (аспарагиновая и глютаминовая);
- минеральные соли;
- ферменты;
- ароматические компоненты.
Нектары безобидных рододендронов, багульников, азалии и чемерицы содержат токсичные вещества, а ядовитых белены, болиголова, олеандра и наперстянки — относительно безопасны. Растения вырабатывают неодинаковые объёмы нектара, например, единичный цветок малины — чемпион в этом отношении, но для учёта общего количества продукта на заданной площади учитывают ещё и численность цветков в соцветии.

Осы, бабочки и шмели охотно употребляют нектар в пищу. Медоносныепчёлы, собирая и пряча его в соты улья, производят мёд. Акации специально выделяют сладкий секрет для привлечения муравьёв, защищающих древесину от зубов травоядных животных. Спектр опылителей может быть широким (эуфилия), подчиняться опылению несколькими родственными или только определённой жизненной формой (олигофилия) или требовать один вид насекомых (монофилия).

Если говорят, что опылитель обладает полилектией, это свидетельствует о чрезвычайно высоком уровне приспособленности к опылению, он способен обслуживать представителей различных семейств. Посещение ограниченной группы, состоящей из одного семейства или растений с однотипными соцветиями, свидетельствует о наличии олиголектии. В случае монолектии опылитель обязательно питается одним видом или родом растений и опыляет только их.
После этого наступает очередь оплодотворения, которое происходит у всех по-разному: чаще период длится несколько недель, но иногда для полноценного слияния половых клеток требуются месяцы. Пыльца, находящаяся на рыльце, для этого должна созреть, обладать достаточной жизнестойкостью и иметь сформированный женский гаметофит (зародышевый мешок). В него и проникает пыльцевая трубка, растущая в направлении завязи через семенной зачаток.
Оказавшись возле яйцеклетки, она спешит разорваться и высвободить два спермия: один соединяется с яйцеклеткой и даёт жизнь зародышу, второй сливается с диплоидным ядром, образуя триплоидную клетку и формируя эндосперм. Такое двойное оплодотворение позволяет получить семя, защищённое кожурой, а из завязи затем возникнет желанный плод.
Примеры в окружающей природе
От опыления зависит сохранение вида, поэтому так важно, чтобы всё проходило успешно. Растения, в отличие от животных, не могут самостоятельно передвигаться в поисках партнёра для размножения и вынуждены уповать на помощь союзников — ветра, воды или живых существ. Самоопыляющимся растениям присущи следующие отличительные признаки:
- цветки не имеют запаха и не выделяют нектара;
- пестики располагаются ниже тычинок;
- пыльца созревает ещё на стадии бутонизации, и опыление осуществляется в нераспустившемся цветке, как у гузмании или арахиса.

Томаты «умеют» самоопыляться факультативно: их тычинки оказались сросшимися, поэтому пестик без труда оплодотворяется своей пыльцой. Малопрогрессивному потомству, полученному таким образом, угрожает вырождение. Этого не случится, если некоторые цветки подвергнутся внутривидовому опылению и приобретут несколько иные родительские зачатки. Тогда они будут обладать более высокими способностями к приспособлению и выживут в процессе естественного отбора.
Мужские деревья двудомных тополя и облепихи интересны только цветками с пыльцой, а женские особи плодоносят. В случае тополя пушистые семена — продукт жизнедеятельности женских деревьев. Если высаживать для озеленения исключительно мужские черенки, можно навсегда избавиться от надоедливого пуха. Оранжевые плоды облепихи дают только женские кусты, но если рядом не посадить мужское растение, то урожая не будет, а обычное соотношение составит 10:1.
Однополые цветки однодомной кукурузы собраны в разнотипные соцветия: мужские — «метёлка» на макушках, женские — «початок» в листовых пазухах. Другие характерные представители однодомных — тыква и огурец — тоже образуют цветки разного типа в пределах одного растения. По внешним признакам они почти неотличимы, но после опыления мужские сразу отмирают и осыпаются, а из оплодотворённых женских будут формироваться завязи.
Источник



