
Центролецитальная яйцеклетка способ гаструляции
У человека имплантация погружная, или интерстициальная. За одни сутки бластоциста погружается в эндометрий почти наполовину, а через 40 часов — полностью. Дефект, образовавшийся в слизистой оболочке матки, восстанавливается в течение 5 суток. Условно имплантация состоит из двух фаз — прилипания (адгезии) бластоцисты к слизистой оболочке матки и погружения (инвазии) бластоцисты в глубину слизистой оболочки.
На 6-е сутки эмбриогенеза бластоциста прикрепляется к эпителию эндометрия (обычно, эмбриональным полюсом в области задней или вентральной стенки у маточного угла). В адгезии бластоцисты важную роль играют вещества группы интегринов, вырабатываемые эпителиоцитами слизистой оболочки матки. В норме у женщин между 19-24 сутками менструального цикла — т. е. в самые оптимальные для взаимодействия с бластоцистой сроки, наблюдается активная экспрессия гена интегрина в эпителиоцитах слизистой оболочки матки.
Гаструляция
У зародыша человека гаструляция инициируется в конце 1-й недели развития, сразу по окончании дробления и сброса прозрачной зоны, когда зародыш начинает погружаться в стенку матки.
1-я фаза гаструляция продолжается всю 2-ю неделю развития. Материал внутренней клеточной массы расщепляется способом деламинации на два листка — эпибласт (верхнюю часть) и гипобласт (нижнюю часть). Клетки гипобласта — мелкие кубические, их цитоплазма имеет пенистый вид, они формируют тонкий слой под эпибластом и граничат с полостью бластоцисты. Клетки эпибласта более высокие и имеют вид псевдомногослойного призматического эпителия.
Клетки эпибласта разобщаются посредством небольших полостей, при слиянии которых в дальнейшем формируется амниотическая полость. В нижней части амниотической полости остается небольшая группа клеток эпибласта — материал будущего зародыша (эмбриобласт) и внезародышевых органов.
2-я фаза гаструляции происходит на 3-й неделе развития, осуществляется способом иммиграции и завершается формированием трех зародышевых листков — эктодермы, мезодермы, энтодермы. В будущем из материала этих листков возникнут ткани эмбриона и внезародышевых органов. Происходящие во 2-й фазе гаструляции события напоминают таковые у куриного зародыша и представителей млекопитающих.
Инициация иммиграции связана с активной и неравномерной в разных участках эпибласта пролиферацией клеток, которая приводит к формированию клеточных потоков и образованию на поверхности эпибласта структур, известных как первичная полоска и гензеновский узелок. Округлый и плоский зародышевый диск во 2-ю фазу гаструляции превращается в вытянутый с расширенным краниальным и более узким каудальным концами.
Особое морфогенетическое значение в раннем эмбриогенезе принадлежит головному отростку (нотохорду). Головной отросток растет в краниальном направлении между эпи- и гипобластом, пока не достигнет прехордальной пластинки, которая является «индикатором» будущего рта зародыша. Головной отросток дает начало развитию хорды зародыша — своеобразного клеточного стержня, который определяет первичную ось эмбриона и придает ему «жесткость».
Хорда формирует ось скелета зародыша человека и является основой развития костей осевого скелета (позвоночника, ребер, грудины, черепа). Вокруг хорды в будущем сформируется позвоночный столб. Нотохорд оказывает важное индуцирующее влияние на дифференцировку прилежащего к нему сверху участка эктодермы в нервную пластинку и далее — в нервную трубку, из которой будут развиваться головной и спинной мозг. К концу 3-й недели эмбриогенеза хорда почти полностью сформирована и простирается от орофарингеальной мембраны до каудального конца зародыша.
Источник
Типы яйцеклеток. Зависимость между типами яйцеклеток и характером дробления.
Типы яйцеклеток. В процессе развития выявляется закономерность, заключающаяся в том, что чем длиннее эмбриональный период, тем больше желтка должно быть накоплено в яйцеклетке. Продолжительность эмбрионального периода зависит от стадии, на которой зародыш переходит к самостоятельному существованию во внешней среде. Если постэмбриональное развитие идет прямым путем, т.е. без личинки и метаморфоза, то желтка в яйцеклетке должно быть больше. По количеству желтка яйцеклетки хордовых делят на алецитальные, олиго-, мезо- и полилецитальные, т.е. с ничтожно малым, малым, средним и большим количеством желтка (от греч. лецитос — желток). У ланцетника, представителя низших хордовых, яйцеклетка олиголецитальная. У большинства позвоночных в яйцеклетках содержится значительное количество желтка. Среди низших позвоночных (Anamnia) наиболее крупные яйца у миксин (кл. Круглоротые), у акул и химер (кл. Хрящевые рыбы) и у безногих амфибий. У осетровых рыб, а также остальных амфибий яйцеклетки мезолецитальные, т.е. имеют среднее количество желтка. У высших позвоночных (Amniota), таких, как пресмыкающиеся, птицы и яйцекладущие млекопитающие,— полилецитальные, т.е. в яйцеклетке очень много желтка. Эмбриональное развитие у них протекает особенно долго. Эта закономерность нарушена у сумчатых и плацентарных млекопитающих, которые имеют олиго- и алецитальные яйцеклетки соответственно. У сумчатых эмбрион выходит из яйцевых оболочек и матки при незавершенном органогенезе, переносится в сумку, где и продолжает развитие. У плацентарных и человека зародыш выходит из яйцевых оболочек еще раньше, в стадии бластоцисты, но затем переходит к внутриутробному существованию, где и завершает все основные периоды развития, подготавливающие его к появлению на свет. Уменьшение количества желтка в яйцеклетках млекопитающих можно назвать вторичным, поскольку их предки, освоившие наземную среду, имели, как и все амниоты, полилецитальные яйца. Полярность яйцеклеток. При малом количестве желтка в яйцеклетке он обычно распределен в цитоплазме равномерно и ядро располагается примерно в центре. Такие яйцеклетки называют изолецитальными (от греч. изос —равный). У большинства позвоночных желтка много, и он распределен в цитоплазме яйцеклетки неравномерно. Это анизолециталъные клетки. Основная масса желтка скапливается у одного из полюсов клетки — вегетативного полюса. Такие яйцеклетки называют телолецйтальными (от греч. телос — конец). Противоположный полюс, к которому оттесняется свободная от желтка активная цитоплазма, называют анималъным. Если желток все же погружен в цитоплазму и не обособлен от нее в виде отдельной фракции, как у осетровых и земноводных, яйцеклетки называют умеренно телолецитальными. Если желток полностью отделен от цитоплазмы, как у амниот, то это резко телолецитальные яйцеклетки. Распределение желтка имеет решающее значение для всех последующих процессов эмбриогенеза, так как определяет будущую пространственную организацию зародыша.
Дробление — это ряд последовательных митотических делений зиготы и далее бластомеров, заканчивающихся образованием многоклеточного зародыша — бластулы. Как правило, бластомеры располагаются в строгом порядке друг относительно друга и полярной оси яйца. Порядок, или способ, дробления зависит от количества, плотности и характера распределения желтка в яйце. По правилам Сакса — Гертвига клеточное ядро стремится расположиться в центре свободной от желтка цитоплазмы, а веретено клеточного деления — в направлении наибольшей протяженности этой зоны. В олиго- и мезолецитальных яйцах дробление полное, или голобластическое. Такой тип дробления встречается у миног, некоторых рыб, всех амфибий, а также у сумчатых и плацентарных млекопитающих. При полном дроблении плоскость первого деления соответствует плоскости двусторонней симметрии. Плоскость второго деления проходит перпендикулярно плоскости первого. Обе борозды первых двух делений меридианные, т.е. начинаются на анимальном полюсе и распространяются к вегетативному полюсу. Яйцевая клетка оказывается разделенной на четыре более или менее равных по размеру бластомера. Плоскость третьего деления проходит перпендикулярно первым двум в широтном направлении. После этого в мезолецитальных яйцах на стадии восьми бластомеров проявляется неравномерность дробления. На анимальном полюсе четыре более мелких бластомера — микромеры, на вегетативном — четыре более крупных —
макромеры. Затем деление опять идет в меридианных плоскостях, а потом опять в широтных. В полилецитальных яйцеклетках костистых рыб, пресмыкающихся, птиц, а также однопроходных млекопитающих дробление частичное, или меробластическое, т.е. охватывает только свободную от желтка цитоплазму. Она располагается в виде тонкого диска на анимальном полюсе, поэтому такой тип дробления называют дискоидальным.
При характеристике типа дробления учитывают также взаимное расположение и скорость деления бластомеров. Если бластомеры располагаются рядами друг над другом по радиусам, дробление называют радиальным. Оно типично для хордовых и иглокожих. В природе встречаются и другие варианты пространственного расположения бластомеров при дроблении, что определяет такие его типы, как спиральное у моллюсков, билатеральное у аскариды, анархичное у медузы.
38. Типы бластул, их связь с типами организации яйцеклетки и способами дробления. Примеры
Голобластическое (полное). Количество желтка в яйце невелико, и оно полностью подвергается дроблению. Вся яйцеклетка дробится на меньшие клетки. Проникая до самых глубоких частей яйца, борозды дробления распространяются в то же время по всей его поверхности.
— Равномерное. Все образующиеся в результате дробления бластомеры являются почти одинаковыми по своим размерам. В результате такого дробления образуется морула. а затем целобластула. Хар-но для яиц ланцетника, иглокожих (морских ежей и морских звезд), кишечнополостных, немертин, в которых желток распределён равномерно.
— Неравномерное. Образующиеся бластомеры разной величины, бластомсры на анимальном полюсе бластулы мельче, чем бластомеры на вегетативном ее полюсе. Это объясняется тем. что желтка больше на одном полюсе яйца (вегетативном), что и тормозит дробление. В результате образуется амфибластула. стенки которой состоят из нескольких слоев клеток, а бластоцель смешена к анимальному полюсу. Хар-но для мезолецитальных яйцеклеток (со средним количеством желтка), например, для яиц амфибий.
Меробластическое (неполное).Дробится только часть яйцеклетки. т.к. в яйцах желтка настолько много что борозды дробления не могут разделить его целиком.
— дискондально. Дробится только сконцентрированная на анимальном полюсе «шапочка» цитоплазмы, где находится ядро зиготы ядро зиготы. Дробление таких яиц происходит только в области зародышевого диска. В результате происходит формирование дискобластулы. Хар-но для полилецитальных яиц. у которых желток сосредоточен на вегетативном полюсе (у птиц, пресмыкающихся, яйцекладущих млекопитающих, акул, скатов. костистых рыб).
— Поверхностное.В глубине желтка происходят первые синхронные деления, не сопровождающиеся образованием межклеточных границ. Ядра, окруженные небольшим количеством цитоплазмы, равномерно распределяются в желтке. Когда их становится достаточно много, они мигрируют в кортикальную цитоплазму, где затем после образования межклеточных границ возникает бластодерма. Дробление происходит только в поверхностном слое цитоплазмы, а центральная часть яйца, занятая желтком, не дробится. Такой характер дробления приводит к образованию перибластулы. Хар-но для яиц насекомых, клещей и большинства других членистоногих.
В зависимости от одновременности делений бластомеров дробление делится на:
• Синхронное.Бластомеры делятся одновременно. Образуется четное число бластомеров. Хар-но для яиц ланцетника, иглокожих.
• Асинхронное. Бластомеры делятся не одновременно. При этом может быть стадия с нечетным числом бластомеров. Например, у земноводных бластомеры анимального полюса делятся быстрее, чем бластомеры вегетативного полюса. Первые дробления у большинства яиц проходят строго синхронно, а затем длительность клеточных циклов бластомеров изменяется и дробление идет асинхронно. Хар-но для яиц земноводных, млекопитающих.
В зависимости от взаимного пространствен расположения бластомеров различают:
• Радиальное. Плоскости последовательных дроблений проходят через яйцо перпендикулярно друг к другу, и бластомеры располагаются симметрично относительно любой плоскости, т.е. бластомеры анимального полюса располагаются точно над бластомерами вегетативного полюса. Хар-но для яиц ланцетника, иглокожих, губок, медуз.
Спиральное.Характеризуется утерей элементов симметрии уже на стадии четырех, (а иногда и двух) бластомеров. В результате наклонного положения митотического веретена все верхние бластомеры сдвигаются в одном направлении по отношению к нижним, т.е. бластомеры анимального полюса располагаются между бластомерами вегетативного полюса. Кольчатые черви, плоские черви, моллюски, немертины.
• Билатеральное. На ранних этапах дробления проявляется билатеральная симметрия. При таком дроблении каждый бластомер правой половины эмбриона соответствует подобному бластомеру левой половины, т.е. образующиеся бластомеры располагаются по бокам исходного бластомера. Круглые черви (нематоды, коловратки), асцидии.
• Анархическое.Беспорядочное расположение бластомеров. Некоторые медузы.
Целобластула возникает в результате полного, равномерного дробления. Один слой клеток (бластодерма) расположен по периферии. Это бластула с однослойной стенкой (бластодермой) и обширным бластоцелем. Хар-на для ланцетника, для иглокожих (морские звезды, морские ежи).
Амфибластула. Образуется в результате полного неравномерного дробления. Дно бластулы занимает все вегетативное полушарие, а бластоцель смещен в анимальную половину. Хар-на для амфибий и кольчатых червей.
Дискобластула. Образуется в результате дискоидального дробления. В телолецитальных яйцах диск бластомеров выгибается над желтком, а между ними возникает полость, которую также называют бластоцелем. Дискоидальное дробление наблюдается лишь в области анимального полюса, где образуется дисковидное скопление клеток —бластодерма От желтка она отделенаподзародышевой полостью. Хар-на для птиц и рептилий (пресмыкающихся).
4) Перибластула. Образуется при поверхностном дроблении. Это заключительная стадия дробления в центролецитальных яйцах членистоногих. Бластоцеля нет, он заполнен желтком. Характерна для насекомых, клещей и других членистоногих.
5) Морула. Бластоцель вообще не возникает, дробление заканчивается на стадии плотного комка клеток (напоминает тутовую ягоду). Это ранняя бластула у ланцетника (когда еще нет бластоцеля). Таким образом, у ланцетников морула является предшественником целобластулы. У млекопитающих стадия морулы предшествует бластоцисте. У некоторых животных (например, у некоторых кишечнополостных) морула — это самостоятельный тип бластулы.
6) Стерробластула — это бластула с однослойной стенкой и очень маленьким центрально расположенным бластоцелем. Бластомеры очень крупные, бластоцеля нет или очень маленький. Хар-на для некоторых членистоногих и моллюсков.
7) Плакула. Это двуслойная пластинка. Хар-на для плоских и кольчатых червей.
8) Бластоциста (бластула типа бластодермического пузырька). Хаар-на для млекопитающих. В ней выделяют наружный слой — трофобласт и внутреннюю массу — эмбриобласт. Трофобласт образует ворсинки, с помощью которых бластоциста имплантируется в слизистую матки.
39. Способы гаструляции, их связь со способами дробления и типами бластул. Примеры. Способы формирования мезодермы.
Инвагинация, или впячивание. Вегетативн полюс бластулы впячивается внутрь, едва не смыкаясь с противоположным. В результ бластоцель почти полностью исчезает и из шара образуется двуслойн зародыш. Внешн слой клеток — эктодерма (наружн листок), а внутрен — энтодерма (внутрен листок). Образующася полость наз полостью первичн кишки, или гастроцелем, вход в нее — первичным ртом, или бластопором. Края бластопора наз губами. Наблюд у животных с изолецитальным типом яиц (ланцетник).
Иммиграция .Активн перемещением части клеток бластодермы в бластоцель с формирова¬нием из них внутреннего энтодермального слоя. Характерна для кишечно¬полостных.
Эпиболия,или обрастание. Мелкие клетки анимальн полюса более активн делятся, обраст и покрыв снаружи крупные богатые желтком клетки вегетативн полюса, из котор и образ-ся внутрен слой. Хаар-но для умеренно телолецитальн яиц земноводн.
Деламинация,или расслоение. Эмбриональн клетки делятся параллельн поверхности бластулы, в результат чего образ наруж и внутрен зародышевые листки. Встреч у птиц, пресмыкающихся и насекомых.
40. Строение нейрулы. Гисто- и органогенез.
НЕЙРУЛА – заключит стадия гаструляции у высших хордовых животных. на этой стадии происход процесс гистогенеза. и орагногенеза.
Производные зародышевых листков
после гаструляции устанав основн план строен тела животных в виде 3 зародыш листков, и можн говорить о новом этапе в развит зародышей — об органогенезе.зародышевые пласты являются гомологичными образованиями.
Производные эктодермы.Из материала наружн зародыш листка — эктодермы — образ нервная пластинка, дающая начало центральн и периферич нервной системе, а также ганглиозная пластинка, из котор формируются ганглии вегетативной нервной системы, клетки мозгового слоя надпочечников, пигментн клетки, задняя доля гипофиза и эпифиз. Из эктодермыразвиваются такжеэпидермис кожи и его производные (волосы, когти, ногти, рога, перья, копыта, панцири, чешуи, чешуйки и т.д), кожн железы (сальные, потовые, молочные и т.д), компоненты органов чувств (органов зрения, слуха, обоняния), эмаль (поверхностный слой) зубов, эпителий ротовой полости и эпителий прямой кишки (т.е. у многих низших животных — эпителий передней и задней кишки).
Производные энтодермы.развиваются — эпителий средн кишки, пищеварит железы (печень, поджелудочн железа), эпителий лёгких. Также образ — ротовая полость, передняя доля гипофиза, глоточная или жаберная область, язык, щитовидн железа, зачаток желчного пузыря.
У зародыш человека и у всех высших позвоночн животн 1я паражаберных мешков участв в возникн наружн слух отверстий, полости среднего уха и евстахиевой трубы.2я параглоточных щелей у зародыша человека приним участие в образ небных миндалин. За счет энтодермальн клеток, отделяющихся от стенок 3го, 4го и 5го жаберн мешков, развив железы внутрен секреции: тимус (вилочковая железа), околощитовидные железы.
Производные мезодермы.В ходе органогенеза происх сложн преобразов клеточн материала мезодермы, приводящ к формиров хрящевого и костного скелета, соединительнотканного слоя кожи, скелетных мышц, а также органов кровенос, выделительн и пол систем организма. Из мезодермы образхорда.
41. Гибридологический метод, Законы Менделя, их цитологические основы.
Гибридологический метод — Анализ закономерностей наследования отдельных свойств и признаков организмов при половом размножении, а также изменчивости генов и их комбинаторики. Метод разработан Г.Менделем. Принципы гибридологического метода: 1) использование в качестве исходных родительских форм гомозиготных по анализируемым признакам особей (т.е. чистые линии); 2) учет при скрещивании не всего многообразия признаков, а лишь одной или нескольких пар альтернативных вариантов признаков; 3) индивидуальный анализ потомства от каждой особи; 4) количественный учет проявлений изучаемых признаков у всех особей.
1)закон единообразия гибридов первого поколения (1865г) – при скрещивании гомозиготных особей, отличающихся по одной паре альтернативных признаков, все потомки в F1 имеют единый фенотип и генотип, и у всех гибридов проявляется доминантный вариант признака родителей.
2) закон расщепления гибридов второго поколения (1865г.) – при моногибридном скрещивании гетерозиготных особей во втором поколении наблюдается расщепление в отношении 3:1 (3/4 особей с доминантным признаком и 1/4 особей с рецессивным признаком.) по фенотипу, и 1:2:! по генотипу.
3) закон независимого наследования признаков (1865г.) – аллели 2х или нескольких генов, если они располагаются в негомологичных хромосомах, и определяемые ими признаки передаются потомству независимо друг от друга, комбинируются во всевозможных сочетаниях. 9:3:3:1
42.Сцепленное наследование. Опыты Моргана. Хромосомная теория наследственности. Кроссинговер, его биологическое значение. Карты хромосом.
Кроссинго́вер -(другое название в биологии перекрёст) — явление обмена участками гомологичных хромосом во время конъюгации при мейозе.
Хромосомная теория Основн полож:
1. Материальными носителями наследственной инф является хромосомы с расположенными в них генами.
2. Гены располагаются в хромосомах в линейной последовательностисти. 3. Аллели 1 гена располагаются в идентичных локусах гомологичных хромосом. 4. В гомологичных хромосомах могут располагаться различные аллели одного и того же гена. 5. Набор генов в каждой из негомологичных хромосом уникален. 6. Каждый биологический вид характеризуется определен набором хромосом — кариотипом. 7. Гены, локализованые в одной хромосоме, образуют группу сцепления и, как правило, наследуются вместе. 8.Числ групп сцепления у представит гомогаметного пола (XX) равно n, у представителей гетерогаметного пола (XY) n+1 (n — гаплоидный набор хромосом). 9. После конъюгации в профазе I делен мейоза происходит обмен участками гомологичных хромосом -кроссинговер, что привод к генетической рекомбинации. 10. Сила сцепления между генами обратно пропорциональна расстоянию между ними (чем больше расстояние между генами, тем больше вероятность кроссинговера). 11. За единицу расстояния между генами принимается 1 морганида, которая означает вероятность 1% кроссинговеров.
сцепл наслед Морган изучал на мухах дрозофилах. изучал наследование 2х признаков: цвет тела: В-серое, в-черное; второй признак длинна крыльев: Wg-длинные, wg-короткие.
Морган скрещивал 2 чистые линии: одна с двумя доминантными аллелими, другая-сдвумя рецессивными. Гибриды первого поколения в соответствии с первым законом Менделя – были единообразные. Затем Морган провёл анализирующее скрещивание гибридной самки. он ожидал получить расщепление 1:1:1:1, но получилось 4е фенотипических класса: 2 из которых большие с родительским сочетанием признаков, и два минорных: с новым сочетанием признаков. Такое расщепление происходит в том случае если изучаемые гены располагаются в одной хромосоме образуя группу сцепления.
самка образ 4 сорта гамет: 2 не кроссинговерных, 2 кроссинговерных.
43. Семейно-генеалогический метод, область его применения. Типы наследования признаков у человека. Характеристика аутосомно-доминантного и аутосомно-рецессивного типов наследования признаков у человека.
семейно—генеалогический метод основан на изучении наследования признака в семьях на протяжении ряда поколений. Этот метод применим в том случае, когда сведения об анализирующем признаке сохраняются в памяти у членов семьи или каких то документах
достоинства человека как объекта для ген исслед:
1) высокая численность (особей) доступных для изучения популяции человека; 2)значительное число и разнообразие известных у человека мутации и хромосом анамалий.
3) высокий уровень изученности биохимии и физиологии человека в норме и при различных заболеваниях.
недостатки человека как объекта для ген исслед:
1) невозможность целенаправленного подбора родительских пар и экспериментальных браков. 2) низкая плодовитость. 3) высок продолжительности жизни, и как следствие – медленная смена поколений (продолжит жизни поколения равна 25-30 годам, аминимал период жизни до наступлен полов зрелости составл 13-35 лет, что позволяет проследить одному исследователю лишь 2, 3 поколения.) 4) большое число групп сцеплений – 23 и значительно наследованное разнообразие. 5) большая фенотипическая изменчивость людей.
Моногенными или менделирующими признаками наз – такие признаки, развитие которых контролируется одним геном. Наследование моногенных признаков, как привило осуществляется в соответствии с законами Менделя.
Однако, при этом следует иметь в виду, что мутации разных генов могут приводить в формированию сходных фенотипических проявлений. (признаков)
Пигментная КСЕРОДЕРМА развив в резулт нарушений сисемму репарации ДНК, которая устраняет изменения в структуре ДНК (тиминовые димеры), возникающие под действием солнечных лучей.
Эта система включ несколько генов, кодирующих синтез ферментов участвующих в репарации ДНК.: 1)Геликаза – разрывает двойные связи а молек ДНК; 2)эндонуклеаза – вырезает поврежденный участок. 3) экзонуклеаза – удаляет поврежденный участок; 4) ДНК-полимераза строит новую копию; 5) лигаза сшивает разрыв сахаро-фосфатного остова
Мутация любого из семи генов системы репарации ДНК приводит к развитию заболевания.
семейно-генеалогический метод основан на изучении наследования признака в семьях на протяжении ряда поколений. Этот метод применим в том случае, когда сведения об анализирующем признаке сохраняются в памяти у членов семьи или каких то документах.
С-г метод получил широк применен в разработке след теоретич и прикладных вопросов:
1) установления наследственного характера признака 2) определения типа наследования; 3) опред пенентрантности 4)изучения сцепления генов и картирования хромосом 5)исследования интенсивности мутацион процесса 6) изучения взаимодействия генов. 7) уточнения генетической природы патологических состояний и составления прогнозов здоровья при медико-генетическом консультировании.
44. Характеристика Х-сцепленного доминантного, Х-сцепленного рецессивного и Y-сцепленного типов наследования признаков у человека.
Х-сцепленный тип наследования
Гены, локализованные в Х-хромосоме, так же как и при аутосомном наследовании, могут быть доминантными и рецессивными. Главной особенностью Х-сцепленного наследования является отсутствие передачи соответствующего гена отца – сыну, т.к. мужчины, будучи гемизиготными (имеют только одну Х-хромосому) передают свою Х-хромосому только дочерям.
Если в Х-хромосоме локализуется доминантный ген, такой тип наследования называется Х-сцепленным доминантным. Для него характерны следующие признаки:
Если болен отец, то все дочери будут больны, а все сыновья здоровы;
Больные дети появляются только в том случае, если болен один из родителей;
У здоровых родителей все дети будут здоровы;
Заболевание прослеживается в каждом поколении;
Если мать больна, то вероятность рождения больного ребенка равна 50%, независимо от пола;
Болеют как мужчины, так и женщины, но в целом больных женщин в семье в 2 раза больше, чем больных мужчин.
При локализации в Х-хромосоме рецессивного гена тип наследования называется Х-сцепленным рецессивным. Женщины почти всегда фенотипически здоровы (носители), т.е. гетерозиготы. Тяжесть болезни зависит от степени поражения репродуктивной системы. Для этого типа наследования характерны:
Заболеванием поражаются преимущественно лица мужского пола;
Заболевание наблюдается у мужских родственников пробанда по материнской линии;
Сын никогда не наследует заболевание отца;
Если пробанд – больная женщина, её отец обязательно болен, а также поражены все её сыновья;
В браке между больными мужчинами и здоровыми гомозиготными женщинами все дети будут здоровы, но у дочерей могут быть больные сыновья;
В браке больного мужчины и женщины-носительницы дочери: 50% -больные, 50% — носительницы; сыновья: 50% — больные, 50% — здоровые.
В браке между здоровым мужчиной и гетерозиготной женщиной вероятность рождения больного ребенка составит: 50% — для мальчиков и 0% — для девочек.
Сестры-носительницы имеют 50% больных сыновей и 50% дочерей-носительниц.

Родословная с Х-рецессивным типом наследования

Родословная с Х-доминантным типом наследования
Y-сцепленный тип наследования
В редких случаях наблюдается отцовский или голандрический тип наследования, обусловленный присутствием мутаций в генах Y-хромосомы.
При этом болеют и передают через Y-хромосому свое заболевание сыновьям только мужчины. В отличие от аутосом и Х-хромосомы, Y-хромосома несет сравнительно мало генов (по последним данным международного каталога генов OMIM, всего около 40).
Небольшая часть таких генов гомологична генам X-хромосомы, остальные присутствующие только у мужчин, участвуют в контроле детерминации пола и сперматогенеза. Так, на Y-хромосоме находятся гены SRY и AZF, ответственные за программу половой дифференцировки.
Мутации в любом из этих генов приводят к нарушениям развития яичек и блоку сперматогенеза, что выражается в азооспермии. Такие мужчины страдают бесплодием, и потому их заболевание не наследуется. Мужчин с жалобами на бесплодие необходимо обследовать на наличие мутаций в указанных генах. Мутациями в одном из генов, расположенных на Y-хромосоме, обусловлены некоторые формы ихтиоза (рыбья кожа), и совершенно безобидный признак — оволосение ушной раковины.
Признак передаётся по мужской линии. В Y-хромосоме имеются гены, отвечающие за оволосение ушной раковины, сперматогенез (азооспермия), интенсивность роста тела, конечностей, зубов.

Родословная с Y-сцепленным типом наследования
45. Моногенное наследование. Типы взаимодействия аллелей одного гена, их характеристика. Множественный аллелизм.
Моногенным называется такой тип наследования, когда наследственный признак контролируется одним геном. Закономерности моногенной наследственности изучал выдающийся ученый Г. Мендель. Он экспериментально обосновал наличие единиц наследственности (наследственных задатков, наследственных факторов) и описал их основные свойства — дискретность, стабильность, специфичность аллельного состояния. Принципиально новым вкладом Г. Менделя в изучение наследования стал метод гибридизации (гибрид — это совокупность). Анализируя результаты моно-и дигибридного скрещивания гороха, он пришел к выводу, что: 1) развитие наследственных признаков зависит от передачи потомкам наследственных факторов; 2) наследственные единицы, которые контролируют развитие отдельного признака — парные: один происходит от отца, второй — от матери. В функциональном отношении факторы имеют свойства доминантного и рецессивного признаков, доминантный признак — которая проявляет себя, рецессивный признак — в одинарной дозе себя не проявляет. 3) наследственные факторы передаются в ряду поколений, не теряя своей индивидуальности, т.е. характеризуются постоянством; 4) в процессе образования половых клеток парные аллельные (формы, состояния) гены попадают в разные гаметы (закон чистоты гамет). Восстановление таких пар происходит в результате оплодотворения; 5) материнский и отцовский организмы в равной степени участвуют в передаче своих наследственных факторов потомкам.
Полное доминирование.Доминантный аллель в гетерозиготе полностью подавляет действие рецессивного аллеля, в результате чего фенотипы гетерозигот и доминантных гомозигот не отличаются друг от друга. подавляют друг друга не гены, а генные продукты. ПРИМЕРЫ: 1)окраска семян гороха 2)форма поверхности семян 3)цвет глаз у человека. (фенотип АА = фенотип Аа)
Неполное доминирование. Доминантн аллель гена не полностью подавл действ рецессив аллеля, вследствие чего у гетерозигот доминант признак выражен менее ярко (промежут признак), чем у доминантнт гомозигот. Расщеплен по фенотипу всегд совпадает с расщеплен по генотипу. Возможные причины:
1.«Эффект дозы»: у доминантн гомозигот экспрессируются 2 доминантн аллеля, а у гетерозигот — только 1.2. Рецессивн аллель в гетерозиготе частичн подавляет действ доминантн аллеля.
Пример: 1)окраска венчика у ночной красавицы 2)окраска шерсти у норок 3) серповидно-клеточная анемия у человека. (фенотип АА > фенотип Аа)
Сверхдоминирование.У гетерозигот доминан признак проявл ярче, чем у доминантн гомозигот. Расщепление по фенотипу всегда совпадает с расщеплен по генотипу. *Возможные причины: 1. У доминантн гомозигот 2 доминантн аллеля частично подавляют экспрессию друг друга (отрицат обратная связь). 2. Рецессивн аллель в гетерозиготе потенцирует (усиливает) действ доминантн аллеля. Пример: Устойчивость к ржавчинным грибам у овса. (фенотип АА -9 – 10 -8 в расчете на клеточное деление. Для эукариот она составляет 10 -6 – 10 -5 на одно поколение).
Индуцированные — вызываются направленным воздействием разнообразных факторов химической, физической или биологической природы. (Эти факторы получили название мутагенов).
По типу клеток, в которых они возникают:
Генеративные мутации — происходят в половых клетках (передаются в ряду поколений);
Соматические мутации — происходят в соматических клетках (не передаются в ряду поколений);
По влиянию на организм:
летальные (семилетальные) — смертельные или снижающие жизнедеятельность;
нейтральные — не оказывающие какого-нибудь влияния на жизнедеятельность орагнизма;
полезные — повышающие жизнедеятельность, имеющие адаптивное значение
По характеру нарушения генетического аппарата:
— Геномные мутации — изменение числа хромосом в кариотипе клеток организма. (Существует два типа таких мутаций: полиплоидия и гетероплоидия или анеуплоидия):
Полиплоидия — кратное увеличение числа гаплоидных наборов хромосом в клетках организма. (Например, в клетках пшеницы разных сортов можно обнаружить 14, 28, или 42 хромосомы ( при n=7, это соответствует 2n, 4n и 6n). В зависимости от количества хромосомных наборов различают формы триплоидные ( 3n), тетраплоидные (4n), пентаплоидные (5n), гексаплоидные (6n) и т.д.);
Анеуплоидия (трисомия, моносомия, нуллисомия) — изменение в геноме количества отдельных хромосом. При гетероплоидии в нормальном хромосомном наборе либо отсутствуют или имеются в избыточном количестве одна или более хромосом. Различают моносомию (2n-1), нуллисомию (2n-2) ,трисомию ( 2n+1) и полисомию ( 2n+х) по отдельным хромосомам.
— Хромосомные мутации — хромосомные перестройки или аберрации которые характеризуются изменениями структуры хромосом. Они разделяются на внутри хромосомные перестройки, когда изменяется структура одной хромосомы, (делеции, дупликации, инверсии), и межхромосомные аберрации, затрагивающие одновременно две негомологичные хромосомы (транслокации):
Делеция — потеря какого-либо участка хромосомы – промежуточного или концевого. (Делеция одних и тех же локусов в обеих гомологичных хромосомах обычно летальны, так как утрачивается значительный объем генетической информации);
Дупликация (повторение) – присутствие одного и того же участка хромосомы более чем в одном экземпляре в одной хромосоме или в разных негомологичных хромосомах;
Инверсия – поворот отдельного участка хромосомы на 180°; при этом число генов в хромосоме остается прежним, а изменяется лишь их последовательность. Инверсия может возникнуть при образовании хромосомой петли с последующим разрывом ее основания и растяжением петли в стороны;
Инсерция — тип хромосомной перестройки, заключающийся в появлении вставки в каком-либо участке нуклеотидной последовательности;
Транслокация — обмен участками в пределах хромосомы (нереципрокные транслокации) , или между негомологичными хромосомами (реципрокные и робертсоновкие транслокации).
— Генные или точковые мутации — изменения химической структуры генов, воспроизводимые в последующих циклах репликации. Генные мутации возникают в результате замены одной или нескольких пар азотистых оснований в структуре ДНК на другие, а также выпадения или добавления пар оснований, что приводит к нарушению порядка считывания генетической информации
Частота возникновения спонтанных мутаций у человека составляет 10 -7 на один нуклеотид при каждом делении клетки.
Генные мутации подразделяются:
а) по механизму возникновения:
-замены одной или нескольких пар азотистых оснований на другие
-выпадения или добавления нескольких пар оснований, (внутригенные перестройки):
— перемещения мобильных элементов
-Alu — повторов (300-500 п.о. — до 100 000 копий на геном)
— длинных вствочных элементов (6500 п.о. — до 50 000 копий на геном)
— динамические мутации (CAG)n (GGG)n
— мутации сайта сплайсинга
б) по влиянию на биосинтез белка
-нейтральные замены, например, ОНП через каждые 1000 п. о.
-миссенс — другой кодон
— беспорядочная последовательность аминокислот (сдвиг рамки считывания
в) по влиянию на биологическую (функциональную) активность белка
— появление новой функции
— появление белков с «токсическими» свойствами
Мутации, встречающиеся в популяции с частотой 1 % и выше называются полиморфизмами.
Мутации по типу замены азотистых оснований могут возникать либо спонтанно, либо под влиянием химических факторов результатом чего служит изменение структуры основания уже включенного в спираль ДНК
Таутомеризация – изменение положения протона, меняющего химические свойства молекулы. В результате таутомеризации аденини приобретает свойства гуанина , гунин-аденина, цитозин –тимина , а тимин-цитозина.
«Динамические» мутации характеризуются нарастанием числа триплетных повторов в кодирующих или некодирующих частях генов.
Увеличение числа повторов по-разному сказывается на функциях соответствующих генов. В случае нейродегенеративных заболеваний, например при хорее Гентингтона, нарастание числа кодонов CAG ведет к синтезу белков с длинными участками, состоящими из глутаминовой аминокислоты. Такие белки, обладают токсичным действием, вызывая гибель нервных клеток.
Увеличение числа триплетных повторов (CGG, CTG, GAA) , например, при синдроме ломкой Х-хромосомы, нарушает транскрипцию и приводит к дефициту соответствующих белков
Правильное удаление интрона возможно лишь тогда , когда на границе с экзоном находятся незаменимые канонические нуклеотиды: ГУ на одном и АГ на другом. Замены этих нуклеотидов приводят к нарушению сплайсинга. Такие мутации получили название мутации сайта сплайсинга.
Молчащие мутации представляют собой замену нуклеотидов, при которых не изменяется смысл кодонов, так как образующиеся кодоны кодируют одну и ту же аминокислоту.
Миссенс мутации представляют собой замену одной или двух нуклеотидов в кодоне, в результате чего изменяется аминокислотная последовательность в молекуле белка – замена одной аминокислоты на другую.
Нонсенс мутации – это такие мутации, которые приводят к появлению стоп-кодона и преждевременной остановке синтеза белка. В результате образуется белок с укороченной полипептидной цепью.
Робертсоновские транслокации хромосом Этот тип перестройки включает две акроцентрические хромосомы, соединенные в области центромеры, с утратой коротких плеч. Полученный в результате сбалансированный кариотип имеет только 45 хромосом, включая транслоцированную, состоящую из длинных плеч двух хромосом. Поскольку короткие плечи всех пяти пар акроцентрических хромосом содержат многочисленные копии генов рРНК, потеря коротких плеч двух акроцентрических хромосом неопасна. Робертсоновские транслокации могут быть как моноцентрическими, так и псевдодицентрическими, в зависимости от позиции точечного разрыва в каждой акроцентрической хромосоме.
Гемоглобинопатии. Это группа наследственных заболеваний, вызываемых первичным дефектом пептидных цепей гемоглобина и связанным с этим нарушением его свойств и функций. Гемоглобинопатии относятся к аутосомно-рецессивным генетическим заболеваниям. Качественные гемоглобинопатии развиваются вследствие мутаций генов, ответственных за синтез определенных аминокислот в бета-цепи глобина. В результате происходит замена одной аминокислоты на другую (глутаминовой кислоты на валин, лизин и пр.). Это приводит к образованию аномального гемоглобина, гораздо менее растворимого, чем нормальный гемоглобин А, придающего красным кровяным тельцам иную форму (мишеневидную, серповидную), что нарушает их функции и уменьшает продолжительность жизни Гемоглобинопатии подразделяются на качественные, обусловленные нарушением структуры (последовательности аминокислот) гемоглобина, и количественные, характеризующиеся снижением образования глобиновых цепей
Серповидно-клеточная анемия (Гемоглобинопатия S). Наиболее частый вид. Подразделяется на гомозиготную форму (собственно СКА) с яркой клинической симптоматикой и гетерозиготное носительство (серповидно-клеточную аномалию), имеющее бессимптомное или легкое течение.
количественным аномалиям гемоглобина талассемия. Наиболее распространенный вариант. Подразделяется на малую талассемию (гетерозиготное носительство с бессимптомным течением или легкой гемолитической анемией) и большую талассемию (анемию Кули) с развернутой тяжелой клинической картиной.
56. Наследственные энзимопатии, механизмы их возникновения. Понятие о болезнях накопления и о болезнях недостатка метаболита. Болезни аминокислотного обмена на примере фенилкетонурии и альбинизма.
Наследственные энзимопатии – это заболевания, вызванные наследственными нарушениями биосинтеза ферментов или их структуры и функции.

Полное или частичное нарушения биосинтеза ферментов вызывают дефекты генов регуляторных белков, которые контролируют синтез ферментов:

Нарушение структуры и функции ферментов вызывают дефекты генов этих ферментов:

У образовавшегося фермента наблюдаются структурные изменения, которые проявляются в изменении его каталитической активности (как правило, она исчезает), чувствительности к активаторам и ингибиторам, сродству к субстратам, оптимумам рН, температуры. В связи с этим изучением констант фермента является решающим в постановке диагноза врожденных энзимопатий.
Фенилкетонури́я — аутосмно-рецессивное заболевание, связанное с нарушением работы фермента фенилаланин-4-гидроксилазы, следовательно, метаболизма аминокислот, главным образом фенилаланина. Сопровождается накоплением фенилаланина. Этот избыток идёт в обходной путь метаболизма с образованием фенилуксусной и фенил –ПВК кислот.
Приводит к тяжёлому поражению ЦНС, проявляющемуся, в частности, в виде нарушения умственного развития. 1 : 10 000 новорожденных, в России 1 : 8000.
— Впервые месяцы жизни ребенок может быть беспокойным или, наоборот, вялым, сонливым.
— Гипопигментация кожи, волос, радужной оболочки (результат снижения уровня тирозина).
— От больного ребенка исходит своеобразный «мышиный запах».
— К 6-ти месяцам выявляется задержка психомоторного развития.
— В это же время экзематозные изменения на коже и судорожные приступы. \
— После 3-х лет умственная отсталость (в 95% случаев это имбецильность или идиотия), нарушение поведения (чаще возбуждение, расторможенность, психотические расстройства), судорожным синдромом.
При своевременной диагностике патологических изменений можно полностью избежать, если с рождения и до полового созревания ограничить поступление в организм фенилаланина с пищей.
Альбинизм – группа наследственных патологий, характеризующихся нарушениями или полным отсутствием пигментации кожи, волос, радужной оболочки глаза. Основная причина развития альбинизма – нарушение работы фермента тироксингидроксилазы, следовательно, нарушения метаболизма аминокислоты тирозина, и, как следствие, полный блок или ослабление синтеза и отложения пигмента меланина.
57. Наследственные энзимопатии, связанные с нарушением углеводного обмена на примере сахарного диабета и галактоземии. Мультифакториальные болезни, доказательства их наследственной природы.
Сахарный диабет (СД) — это группа метаболических нарушений углеводного обмена, которые обусловлены дефицитом инсулина (абсолютным или относительным), приводящим к хронической гипергликемии. Организм не способен переработать глюкозу, что приводит к повышению уровня глюкозы в крови.
Тип наследования аутосомно-рецессивный.
Заболевание связанное с нарушением обмена углеводов. Выделяют три типа заболевания в зависимости от ферментов участвующих в превращении галактозы в клетках печени в глюкозу — галактоземия I типа (недостаточность галактозо-1-фосфат уридил трансферазы), галактоземия II типа (недостаточность галактокиназы), галактоземия III типа (недостаточность галактозо-4-эпимеразы).
Наиболее часто встречается галактоземия I типа (частота 1:15000 новорожденных). Ген заболевания картирован на хромосоме 9р13.
Токсическое действие заключается в ингибировании бактерицидной активности лейкоцитов.
— Галактоземия тип I, манифестирует на 1-2 неделе жизни. У ребенка отмечаются частые срыгивания, плохая прибавка массы тела, нарушения стула.
— Затем присоединяются симптомы поражения печени — гепатомегалия или гепатоспленомегалия, гипербилирубинемия, повышение уровня печеночных ферментов, гипоальбуминемия, нарушение свертываемости крови, асцит.
— Нередко отмечаются гипогликемические состояния.
— Формируется катаракта в первый месяц жизни.
Без соответствующего лечения дети погибают в течение первого полугодия жизни от острой печеночной недостаточности.
Описаны также доброкачественные варианты заболевания, протекающие без выраженных изменений со стороны печени, что вероятно, обусловлено генетической гетерогенностью галактоземии I типа и влиянием средовых факторов.
Основным методом лечения является диетотерапия. Назначение безлактозной смеси приводит к быстрому купированию основных клинических симптомов и восстановлению функции печени. Однако у большинства пациентов, находящихся на строгой диете в дальнейшем возникают неврологические и эндокринологические расстройства.
Мультифакториальные заболевания – это заболевания, возникающие при неблагоприятном сочетании ряда факторов: генетических особенностях и влияния внешней среды. Само наличие генетической предрасположенности к заболеванию не обязательно приведет к развитию этого заболевания. Однако при наличии неблагоприятных факторов внешней среды, человек с наследственной предрасположенностью имеет значительно большую вероятность заболеть, чем люди, не имеющие такой предрасположенности. Для примера можно взять сахарный диабет. Сахарный диабет — гетерогенное по своей природе заболевание, в этиологии и патогенезе которого участвуют как внутренние (генетические, иммунные), так и внешние (вирусные инфекции, интоксикации), факторы, взаимодействие которых приводит к нарушению углеводного обмена.
58. Наследственные энзимопатии, связанные с нарушением липидного, пуринового и минерального обмена на примере синдромов Тея-Сакса, Леша-Найана и болезни Вильсона-Коновалова.
Болезнь Тея-Сакса (БТС)
Амавротическая идиотия (болезнь Тея-Сакса) относится к заболеваниям, связанным с нарушением липидного обмена.
Причина — снижение активности фермента гексозаминидазы А в организме. В результате происходит разрушение аксонов нервных клеток
Тип наследования болезни — аутосомно-рецессивный. Ген локализован на длинном плече 15-й хромосомы
Заболевание проявляется тогда, когда в нервных клетках мозга накапливается опасное количество ганглиозидов, что в результате приводит к преждевременной смерти этих клеток. Липид ганглиозид откладывается в клетках мозга, печени, селезенки и других органах
Признаки. Болезнь проявляется первые месяцы жизни. Ребенок становится вялым, малоподвижным, безразличным к окружающим. Задержка психического развития приводит к снижению интеллекта до степени идиотии. Отмечается мышечная гипотония, судороги, характерный симптом «вишневой косточки» на сетчатке глаза
К концу первого года жизни наступает слепота — атрофия зрительных нервов. Позднее развивается полная обездвиженность. Смерть наступает в 3-4 года
На сегодня не существует никаких эффективных лекарств или иных методов лечения для этой болезни.
Синдром Леша-Наяна — встречается редко (1:300000 новорожденных) и наследуется по Х-сцепленному рецессивному типу. Болезнь развивается в грудном возрасте, проявляется мышечным гипертонусом, повышенной рефлекторной возбудимостью, олигофренией, импульсивной склонностью ребенка к самоповреждениям. Выделяют также атипичную форму взрослых, которая проявляется симптомами подагры.
Первичный дефект: это нехватка фермента гипоксантин-фосфорибозилтрансферазы (ГФРТ), необходимого для синтеза ДНК. Он катализирует превращение свободных пуриновых оснований — гуанина и гипоксантина — донуклеотидов. При недостатке этого фермента конечным продуктом распада основ является мочевая кислота.Высокое содержание ее солей приводит к формированию уратов и развития почечнокаменной болезни.
Фермент ГФРТ находится во всех соматических клетках. Поэтому возможна точная диагностика путем определения его активности в клетках. На практике для этого используют культивируемые фибробласты кожи, а при пренатальной диагностике — амниотические клетки. Выявление активности фермента в культивируемых клетках позволяет, путем совместного культивирования клеток от разных больных, дифференцировать генетически различные формы в случае метаболического кооперирования (частичного обновления активности ГФРТ в клетках больных)
Болезнь Вильсона—Коновалова — наследственное заболевание, при котором происходит хроническая интоксикация организма медью вследствие нарушения её транспорта и накопления.
Причиной заболевания является мутация гена, отвечающего за синтез белка, который осуществляет транспорт меди (церулоплазмин). Этот ген имеет название AT- P7B, и он находится на длинном плече хромосомы 13 (13q14.3-q21.1)..
Аутосомно-рецессивный тип заболевания
Заболеваемость у новорожденных 1 на 30000 — 1 на 55000. Чаще болеет мужской пол, чем женский (4:1)
Признаки. Медь оказывается слабо связанна с альбумином и аминокислотами крови, легко отщепляется от них, в большом количестве выделяется с мочой и откладывается в тканях, главным образом в печени, головном мозге и роговице. Избыток свободной меди подавляет активность окислительных и некоторых других ферментов, что приводит к гибели клеток. Поражения печени с распадом ее ткани и снижением барьерной функции ведет к аутоинтоксикации продуктами гепатолиза и чужеродными продуктами, поступающими из кишечника.
Согласно классификации Н.В. Коновалова различают пять форм болезни Вильсона:
Брюшная форма развивается преимущественно до 40 лет. Характерный признак — тяжелое поражение печени по типу цирроза печени, хронического гепатита, фульминантного гепатита.
Ригидно-аритмогиперкинетическая форма манифестирует в детском возрасте. Начальные проявления — мышечная ригидность, амимия, смазанность речи, трудности при выполнении мелких движений, умеренное снижение интеллекта. Для этой формы заболевания характерно прогрессирующее течение, с наличием эпизодов обострения и ремиссии.
Дрожательная форма возникает в возрасте от 10 до 30 лет. Преобладающим симптомом является тремор. Кроме того, могут наблюдаться брадикинезия, брадилалия, тяжелый психоорганический синдром, эпилептические приступы.
Экстрапирамидно-корковая форма встречается весьма редко. Ее начало схоже с началом какой-либо из вышеперечисленных форм. Для нее характерны эпилептические припадки, экстрапирамидные и пирамидные нарушения и выраженный интеллектуальный дефицит.
59. Популяционно-статистический метод. Закон Харди-Вайнберга. Применение закона Харди-Вайнберга для определения частоты встречаемости скрытых носителей аутосомно-рецессивных и Х-сцепленных рецессивных признаков.
Математическое обоснование современной эволюционной теории нашло отражение в законе Харди-Вайнберга. Который был сформулирован независимо друг от друга математиком Г. Харди (Англия) и врачом В. Вайнбергом (Германия) в 1908 г.
Этот закон утверждает, что частоты аллелей и генотипов в данной популяции будут оставаться постоянными из поколения в поколение при выполнении следующих условий:
1) численность особей популяции достаточно велика (в идеале — бесконечно велика),
2) спаривание происходит случайным образом (т. е. осуществляется панмиксия),
3) мутационный процесс отсутствует,
4) отсутствует обмен генами с другими популяциями,
5) естественный отбор отсутствует, т. е. особи с разными генотипами одинаково плодовиты и жизнеспособны.
Иногда этот закон формулируют иначе: в идеальной популяции частоты аллелей и генотипов постоянны. (Поскольку описанные выше условия выполнения данного закона и есть свойства идеальной популяции.)
Математическая модель закона отвечает формуле:
Наследственность сама по себе не изменяет частоты аллелей и генотипов по определенному локуcу.
В свободно скрещивающейся популяции частоты генотипов связаны с частотой генов квадратичными соотношениями.
Р-частота доминантного аллеля
q- частота рецессивного аллеля
Медицинское значение генетического полиморфизма определяется тем, что это явление лежит в основе наследственно обусловленных индивидуальных реакций людей на действие факторов внешней среды, лекарственные препараты и т.п.
60. Генетический полиморфизм на примере наследования групп крови человека по системе АВО. Применение закона Харди-Вайнберга для определения частоты встречаемости генотипов людей с различными группами крови.
Закон Харди — Вайнберга — положение популяционной генетики, гласящее, что в популяции бесконечно большого размера, в которой не действует естественный отбор, не идет мутационный процесс, отсутствует обмен особями с другими популяциями, не происходит дрейф генов, все скрещивания случайны — частоты генотипов по какому-либо гену (в случае если в популяции есть два аллеля этого гена) будут поддерживаться постоянными из поколения в поколение и соответствовать уравнению:
Р-частота доминантного аллеля
q- частота рецессивного аллеля
2pq- доля гетерозигот
61. Эволюция органов дыхания у беспозвоночных Основные направления эволюции дыхательной системы. Строение органов дыхания у круглоротых и рыб.
Эволюционные предпосылки возникновения дыхательной системы: 1. Появление целома (вторичной полости тела, пространства между стенкой тела и внутренними органами у высших многоклеточных животных, ограниченного собственными эпителиальными оболочками мезодермального происхождения) и гомономной сегментации тела (сегменты тела одинаковые по строению). 2. Появление в каждом сегменте параподий – сложных боковых выростов тела, состоящих из верхней и нижней лопастей, снабженных щетинками (через поверхность параподии, обильно снабжаемой кровью, осуществляется газообмен).
В отличие от всех других позвоночных у круглоротых в жаберных щелях развиваются жаберные мешки, имеющие энтодермальное происхождение. Внутренняя поверхность жаберных мешков образует многочисленные складки слизистой оболочки, в которых расположена густая сеть мелких кровеносных сосудов — капилляров. У миноги каждый жаберный мешок (всего их 7 пар) открывается наружу самостоятельным наружным отверстием. Внутреннее отверстие жаберного мешка соединяет его полость с дыхательной трубкой, которая представляет собой слепой вырост, спереди соединенный с ротовой полостью и отграниченный от нее подвижной складкой — парусом.
У плавающей миноги вода поступает в ротовую полость, оттуда проходит в дыхательную трубку и, пройдя сквозь жаберные мешки, через их наружные отверстия выбрасывается наружу. При этом кислород, растворенный в воде, проникает в капилляры и связывается пигментом крови, а насыщающий венозную кровь углекислый газ переходит в воду и с ней выводится наружу.
В том случае, когда минога питается (или если она присосалась к какому-либо предмету), ток воды через ротовую полость становится невозможным. Парус при этом закрывает вход в дыхательную трубку, не пропуская воду, а жидкая пища проходит из ротовой полости в пищевод. При этом дыхание осуществляется иным путем: под воздействием мускулатуры стенок тела жаберные мешки сжимаются, и вода выталкивается через наружные жаберные отверстия (активный выдох); далее, благодаря упругости хрящевой жаберной решетки жаберная область снова расширяется, и вода через те же наружные отверстия вновь засасывается в жаберные мешки (пассивный вдох).
62. Эволюция дыхательных путей и легких утетрапод. Филогенетически обусловленные аномалии развития дыхательной системы у человека.
Дыхательная система земноводных
1. У земноводных имеются ноздри, снабженые клапанами и открывающиеся хоанами в ротоглоточную полость.
У хвостатых амфибий в нее открывается гортань, состоящая из хрящей. Наиболее развиты пара черпаловидных, образующих гортанную щель.
Для бесхвостых амфибий характерно гортанно-трахеальное хрящевое кольцо.
2. Органы дыхания — парные мешковидные или крупноячеистые легкие с упругими стенками.
У бесхвостых они подвешиваются к нижней части гортанной камеры, а у хвостатых соединяются с ней длинной трубкой – трахеей, в стенке которой имеются хрящевые элементы, не позволяющие трубке спадаться.
Акт дыхания в связи с отсутствием грудной клетки происходит следующим образом:
Животное открывает клапаны ноздрей и опускает ротовое дно: воздух заполняет ротовую полость.
После этого клапаны закрываются и ротовое дно поднимается: воздух через гортанную щель проталкивается в легкие, которые несколько растягиваются.
Затем животное снова открывает клапаны ноздрей: эластичные стенки легких спадаются и воздух выталкивается из них.
Дыхательная система пресмыкающихся
На передней части головы расположены ноздри, открывающиеся хоанами в ротовую полость. У крокодилов хоаны открываются в глотку, благодаря чему крокодилы могут дышать во время захватывания пищи.
Из хоан воздух попадает в гортань, состоящую из перстневидного и двух черпаловидных хрящей, а оттуда – в трахею.
В нижней части трахея разделяется на два главных бронха, которые соединяются с легкими, но не разветвляются в них.
Легкие представляют собой мешки, имеющие на внутренней поверхности мелкоячеистое строение. У змей одно легкое.
Появляется грудная клетка за счет срастания ребер с грудиной
Дыхание осуществляется путем изменения объема грудной клетки за счет работы межреберных мышц.
Газообмен через кожу у рептилий отсутствует.
Дыхательная система птиц
Как и у млекопитающих, начальная часть дыхательной системы птиц представлена трахеей и бронхами. Первичные бронхи входят в легкие, где дают ряд ответвлений (вторичные и третичные бронхи) и в конце концов открываются в воздушные мешки. Воздушные мешки представляют собой тонкостенные полости, образующиеся как выросты бронхов и располагающиеся между внутренними органами; ответвления воздушных мешков заходят в некоторые трубчатые кости. У большинства птиц имеется пять пар воздушных мешков, функционально объединяющихся в две группы: передние и задние. К передним мешкам относятся шейные, межключичные (обычно сливаются в один непарный мешок; его выросты проникают в кости пояса передних конечностей и в плечо) и переднегрудные. Более объемистые задние мешки — заднегрудные и брюшные — размещены от печени до конца тазовой области; выросты брюшных мешков проникают в полости бедренных костей.
При вдохе коракоид, вилочка и грудинные отделы ребер отклоняются вперед, что вызывает движение грудины вперед и вниз, сопровождающееся латеральным движением ребер, расположенных позади легких. В результате создается пониженное давление внутри полости тела, обеспечивающее растяжение эластичных воздушных мешков и засасывание в них воздуха. При выдохе грудина приводится в прежнее положение, грудная клетка сжимается, давление в полости тела возрастает, и воздух выталкивается из воздушных мешков. Несмотря на то что дыхательные движения состоят, как и у всех животных, из вдохов и выдохов, через легкие птиц воздух идет все время в одном направлении. При вдохе наружный воздух поступает в задние воздушные мешки и частично в легкие; передние воздушные мешки на этой фазе дыхательного цикла заполняются воздухом из системы третичных бронхов, т. е. из легких собственно; вдыхаемый воздух в эту группу мешков не попадает. При выдохе мешки сжимаются, и воздух из задних мешков выталкивается в легкие, т. е. в систему парабронхов, а из передних мешков — в трахею и через нее наружу. На этой фазе дыхательного цикла воздух из задних мешков не поступает в первичные бронхи, а из передних — в легкие. Таким образом, через газообменные структуры легочной ткани воздух и при вдохе, и при выдохе идет в одном направлении, что обеспечивает непрерывность процесса газообмена. Экологические преимущества однонаправленного потока воздуха в легких связаны прежде всего с отсутствием «холостых» фаз дыхательного цикла; газообмен в легких птиц происходит непрерывно как при вдохе, так и при выдохе.
Дыхательная система млекопитающих
У млекопитающих развитие дыхательной системы достигает вершины.
Носовая полость разделяется на преддверие, дыхательный и обонятельный отделы. Функция преддверия — улавливание относительно крупных частиц (пыли), проникающих с воздухом. Мелкие пылевые частицы улавливаются в дыхательном отделе носовой полости, выстланном слизистой оболочкой с мерцательным эпителием, снабженным обильными кровеносными капиллярами; здесь происходит не только дальнейшая очистка воздуха от механических примесей, но и его обеззараживание с помощью слизи, содержащей активные бактерицидные вещества; одновременно воздух, проходя между складками носовой раковины, увлажняется и согревается протекающей по капиллярам кровью и отдает ей часть кислорода. Обонятельный отдел представляет расширение с вдающимися в него выростами боковых стенок — образуется лабиринт полостей, выстланных обонятельным эпителием. Пройдя через хоаны в глотку, воздух попадает в гортань.
В основании гортани лежит кольцеобразный перстневидный хрящ, характерный для всех наземных позвоночных. Переднюю и боковые стенки гортани образует имеющийся только у млекопитающих щитовидный хрящ. Черпаловидные хрящи расположены по бокам спинной стороны гортани. К переднему краю щитовидного хряща примыкает тонкий надгортанник, прикрывающий вход в гортань при прохождений пищи через глотку.
Голосовые складки — парные складки слизистой оболочки, расположенные между щитовидным и черпаловидными хрящами.
К гортани прилегает трахея, которую поддерживают хрящевые кольца. Трахея разделяется на два бронха, отличающихся от нее лишь меньшим диаметром; достигнув легких, они делятся на все более мелкие трубочки, образуя бронхиальное древо. Самые мелкие трубочки – бронхиоли — открываются в расширения, выстланные дыхательным эпителием, — пузырьки- альвеолы размером 25- 400 мкм, в стенках которых ветвятся многочисленные капилляры. Такая структура легких увеличивает их дыхательную поверхность, которая в 50-100 раз превышает поверхность тела.
Механизм дыхания млекопитающих двойной. При так называемом реберном -дыхании с помощью межреберной мускулатуры изменяется объем грудной клетки; при диафрагмальном — тот же объем меняется при опускании и подъеме мускульной грудобрюшной преграды-диафрагмы. В разном сочетании оба механизма функционируют у всех млекопитающих; у хищников преобладает первый механизм, у копытных- второй.
Филогенетически обусловленные пороки развития органов дыхания у человека
Эзофаготрахеальные свищи — врожденные пороки развития пищевода и трахеи, развивающиеся в эмбриогенезе человека на этапе формирования верхних дыхательных путей уже после образования вентролатеральных выпячиваний стенок эмбриональной глотки.
Бронхолегочные кисты — округлые полости в легких, которые развиваются в результате нарушения дифференцировки альвеол на более поздних стадиях эмбрионального периода онтогенеза.
Кистозная гипоплазия легких, при которой оказываются недоразвитыми целые доли легкого. Такие участки представляют собой многочисленные полости, связанные с крупными бронхами, со сравнительно небольшой дыхательной поверхностью и существенно ослабленным газообменом. Тяжелая степень кистозной гипоплазии легких нередко приводит к летальному исходу вследствие дыхательной недостаточности.
Гипоплазия диафрагмы — порок вспомогательной дыхательных мышцы – встречается различной степени тяжести: от небольших дефектов строения купола до полной ее аплазии, несовместимой с жизнью.
63. Эволюция кровеносной системы у беспозвоночных. Основные направления эволюции кровеносной системы беспозвоночных и позвоночных.
Основные направления эволюции кровеносной системы
Эволюция органов кровообращения у позвоночных шла по пути формирования двухкамерного сердца и постепенного увеличения количества его камер, а также с постепенным уменьшением количества дуг аорты.
Появление трехкамерного сердца и правой и левой дуг аорты связано с формированием легочного дыхания и второго круга кровообращения.
Появление четырехкамерного сердца и сохранение только одной дуги аорты связано с полным разделением артериального и венозного кровотоков, что создало условия для формирования теплокровности.
У низших беспозвоночных животных, т.е. у губок, кишечнополостных и плоских червей, доставка питательных веществ и кислорода от места их восприятия до частей тела происходит путем диффузных токов в тканевых жидкостях. Но у некоторых животных появляются пути, по которым идет циркуляция. Так возникают примитивные сосуды.
Дальнейшая эволюция кровеносной системы связана с развитием в стенках сосудов мышечной ткани, благодаря чему они могут сокращаться, а еще позже эволюция связана с превращением жидкости, заполняющей сосуды в особую ткань – кровь, в которой образуются различные кровяные клетки.
Впервые кровеносная система появилась у кольчатых червей, она замкнутая. Имеется 2 сосуда – спинной и брюшной, связанные между собой кольцевыми сосудами, идущими вокруг пищевода. Движение крови происходит в определенном направлении – на спинной стороне к головному концу, на брюшной – назад благодаря сокращению спинного и кольцевых сосудов
У членистоногих незамкнутая кровеносная система. На спинной стороне имеется пульсирующий сосуд, разделенный на отдельные камеры, так называемыми сердца, между которыми имеются клапаны. При последовательном сокращении сердец кровь поступает в сосуды, а затем изливается в щелевидные пространства между органами. Отдав питательные вещества, кровь медленно стекает в околосердечную сумку, а потом через парные отверстия в сердца.
У моллюсков кровеносная система также незамкнутая. Сердце состоит из нескольких предсердий, куда впадают вены и одного достаточно развитого желудочка, от которого отходят артерии.
64. Эволюция сердца и кругов кровообращения у позвоночных.


65. Эволюция дуг аорты у позвоночных. Филогенетически обусловленные аномалии развития кровеносной системы у человека.
В эмбриональном периоде онтогенеза позвоночных формируется 6 пар эмбриональных дуг аорты.
Хрящевые рыбы. У хрящевых рыб I пара редуцируется, из остальных 5 пар формируются приносящие жаберные артерии.
Костные рыбы. У представителей всех классов, начиная с костных рыб, I и II пары эмбриональных дуг редуцируются. У костных рыб из III, IV, V, VI пар развиваются 1-4 пары приносящих жаберных артерий.
Наземные позвоночные. У наземных позвоночных (тетрапод) V пара редуцируется, сохраняясь в виде рудимента только у хвостатых амфибий. Из III пары развиваются сонные артерии. Из VI – легочные (у амфибий легочно-кожные). Из IV пары формируются собственно дуги аорты.
У амфибий и рептилий имеется 2 дуги – правая и левая.
У хвостатых амфибий и черепах легочные артерии сохраняют связь с дугами аорты в виде боталлова протока.
Птицы. У птиц сохраняется только правая дуга аорты,
Млекопитающие. У млекопитающих сохраняется – только левая (остаток правой преобразуется в плечеголовной ствол).
Филогенетически обусловленные пороки развития сердца у человека
Шейная эктопия сердца — нарушение смещения сердца в переднее средостение (после 20 суток эмбриогенеза), его задержка в области первичной закладки, т.е. позади головы, как и у предковых форм. Приводит к гибели ребенка сразу после рождения.
Сохранение двухкамерного сердца к моменту рождения.
Врожденные дефекты межпредсердной перегородки встречаются с частотой 1 случай на 1000 рождений. Иногда межпредсердная перегородка самопроизвольно зарастает к двум месяцам после рождения.
Врожденные дефекты межжелудочковой перегородки встречаются с частотой от 2 до 5 случаев на 1000 рождений. Иногда межжелудочковая перегородка самопроизвольно зарастает к двум годам после рождения.
Филогенетически обусловленные пороки развития дуг аорты у человека
Персистенция правой и левой дуг аорты – производных четвертой пары эмбриональных жаберных дуг, — порок, при котором обе дуги срастаются в системную спинную аорту позади пищевода и трахеи, что приводит к нарушению глотания и удушью (частота 1 случай на 200 вскрытий детей, умерших от пороков сердца).
Нарушение редукции правой дуги аорты с редукцией левой встречается вдвое чаще и зачастую не имеет клинических проявлений.
Персистенция боталлова протока между дугой аорты и легочным стволом проявляется смешением артериальной и венозной крови (частота 0,5-1 на 1000 новорожденных).
Персистенция первичного эмбрионального ствола – единственного сосуда, отходящего от сердца обычно в области дефекта межжелудочковой перегородки, является тяжелейшим пороком развития, несовместимым с жизнью.
Транспозиция сосудов – относительно редкий порок развития, при котором аорта отходит от правого желудочка, а легочный ствол от левого, также заканчивается летальным исходом.
Филогенетически обусловленные пороки развития полых вен у человека
Персистенция двух верхних полых вен. Тяжесть клинических проявлений зависит от того, куда впадают сосуды.
а) при впадении обоих вен в правое предсердие клинические проявления отсутствуют;
б) при впадении левой полой вены в левое предсердие происходит попадание венозной крови в большой круг кровообращения;
в) при впадении обоих вен в левое предсердие порок несовместим с жизнью.
2.Отсутствие нижней полой вены – редкий сосудистый порок, который компенсируется развитием непарной и полунепарной вен, являющихся гомологами задних кардинальных вен.
66. Эволюции выделительной системы у беспозвоночных. Основные направления эволюции выделительной системы у беспозвоночных и позвоночных.
Выделительная система имеет мезодермальное происхождение, а точнее образуется из нефротома сомитов. Она выполняет функцию удаления жидких продуктов обмена веществ из организма.
Впервые выделительная система как самостоятельная система появилась у плоских червей в виде протонефридиев. Протонефридии начинаются в глубине паренхимы клетками звездчатой формы-терминальных клеток с мерцательным пламенем.
У круглых червей выделительная система представляет 1-2 одноклеточными кожными железами, заменившими протонефридии. От желез отходят выросты в виде двух боковых каналов, лежащих в боковых валиках гиподермы.
Другой тип выделительной системы появляется у кольчатых червей, имеющих гомономную сегментацию и вторичную полость тела (целом). Это метамерно повторяющаяся в каждом сегменте пара метанефридиев, каждая из которых состоит из воронки, открывающейся в целом одного сегмента, называемая нефростомом, отходящего от него канальца, прободающего дессипимент и выделительной поры (нефропора) в другом сегменте.
У моллюсков и членистоногих вторичная полость редуцируется, и нефридии выделения значительно меняются. У ракообразных и большинства моллюсков они образуют компактные органы, напоминающие почки позвоночных. У ракообразных и насекомых вместо нефридиев появляются выделительные трубки, которые начинаются в миксоцеле и впадают в кишечник на границе среднего и заднего отделов. Они называются мальпигиевыми сосудами.


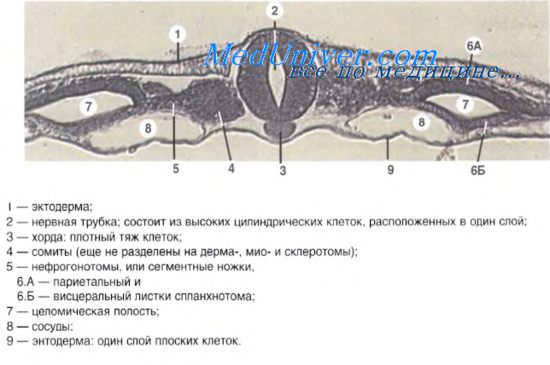
67. Эволюция почки у позвоночных. Эволюция нефрона.
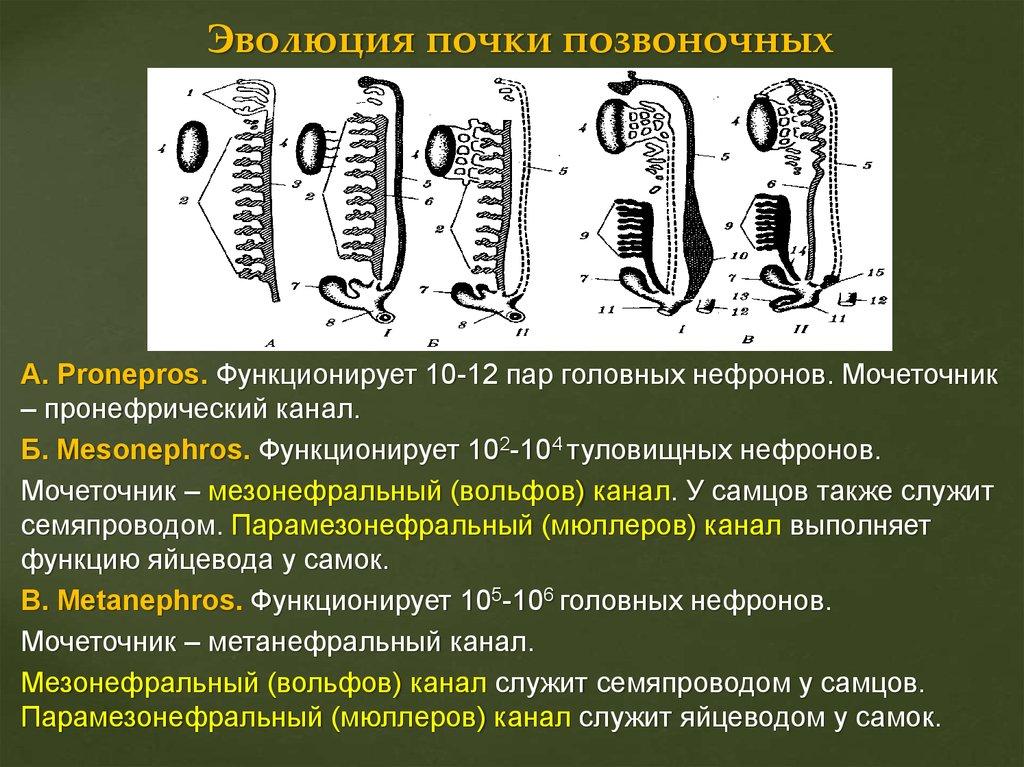


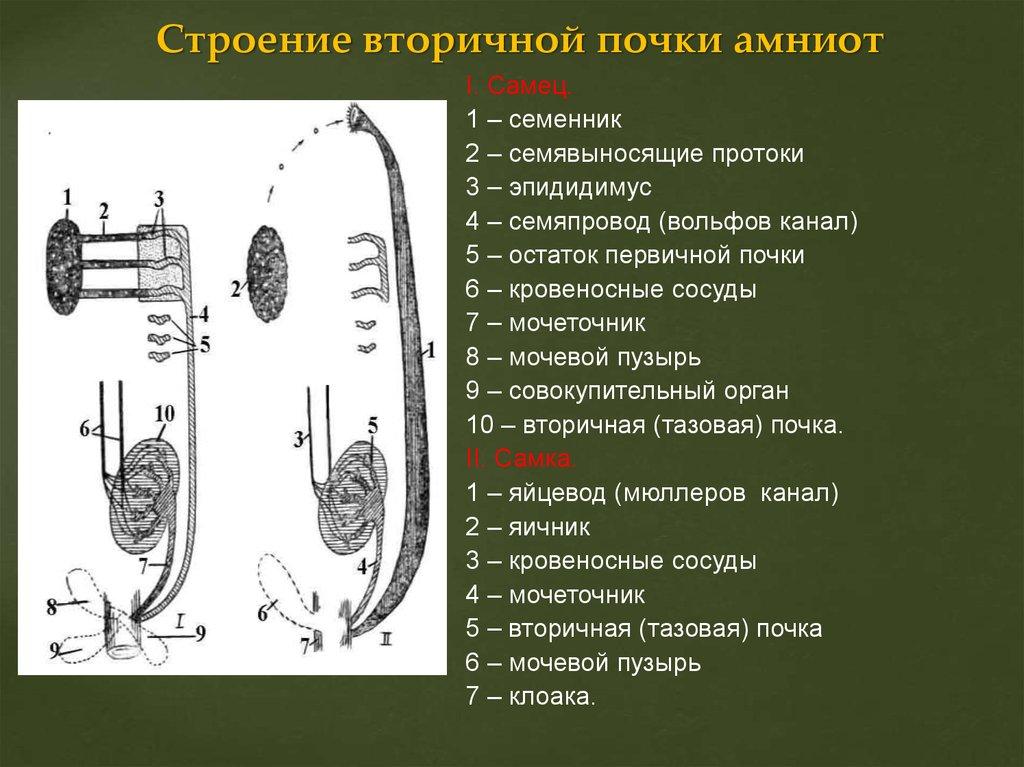
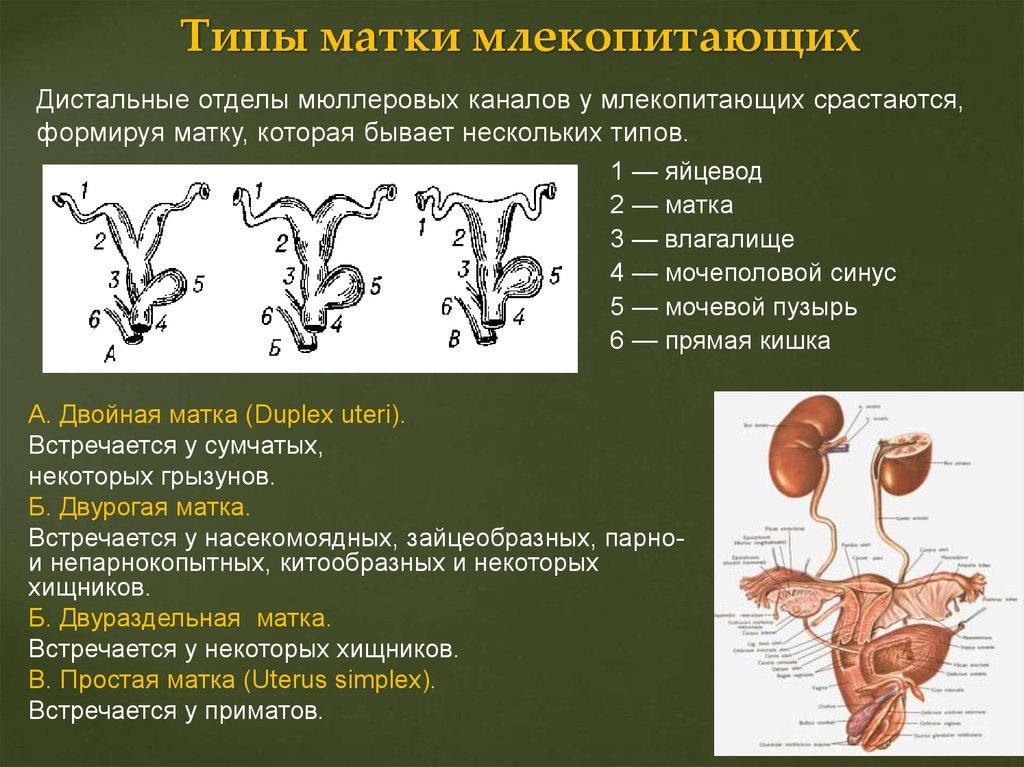
68. Вольфов и Мюллеров каналы, их функции у самцов и самок анамний и амниот. Типы матки млекопитающих. Филогенетически обусловленные аномалии развития мочеполовой системы у человека.



69. Эволюция нервной системы у беспозвоночных. Основные направления эволюции нервной системы у беспозвоночных и позвоночных.
Впервые нервная система появляется у кишечнополостных животных. Нервная система кишечнополостных является диффузной, т. е. у них отсутствуют выраженные скопления нервных клеток, образующих более-менее равномерную сеть.
не имеет выраженных центров;
нейроны располагаются в эктодерме;
нейроны имеют многочисленные одинаковые отростки, с помощью которых они контактируют друг с другом и иннервируемыми клетками тела;
по анатомической организации диффузная нервная система напоминает сеть;
нейроны обладают одновременно и сенсорными (чувствительными) и эффекторными функциями — они и воспринимают раздражения и передают нервные импульсы к клеточным комплексам тела, запуская их реакцию;
для нейронов характерна общая генерализорванная реакция.
Такая нервная система может организовывать только простые движения — например, гидра сжимается в комочек, если к ней прикоснуться иголкой. У медуз, в связи с их подвижным образом жизни, сложилась более совершенная нервная система: имеется скопление нервных клеток в виде кольца по краю зонтика. Также у медуз есть отолитовый аппарат (орган равновесия) и имеется функциональное разделение нейронов на две группы, отвечающие за плавательную и пищевую активность. У более высокоорганизованных животных нервные клетки располагаются более тесно друг к другу, образуя нервные узлы. Благодаря синаптическим контактам нервных клеток, образующих узлы, в них становится возможна обработка поступающей информации и выработка команд, поступающих к рабочим органам: железам и мышцам.
У плоских червей возникает билатеральная симметрия, соответственно, у них дифференцируется головной и хвостовой конец тела. Нервная система плоских червей лестничного типа.
1. несколько крупных ганглиев в головном конце тела
2. два (или больше) нервных ствола,
3. нервные стволы соединенных друг с другом перемычками (комиссурами)
4. к головному концу смещаются нервные элементы и органы чувств: тактильные рецепторы и хеморецепторы, а у свободноживущих червей — и световые рецепторы.
5. Нейроны головных узлов получают, перерабатывают информацию, поступающую от органов чувств, и передают нервные импульсы к мышечным клеткам, что позволяет животным осуществлять несложные движения в виде ползания и плавания.
У кольчатых червей обнаруживается симметричное строение тела. Нервная система кольчатых червей узлового типа.
и нервной системы, которая представлена двумя цепочками узлов, состоящих из нервных клеток и нервных волокон. У них впервые в процессе эволюции появляется нервная система узлового типа.
В брюшной области узлы одной стороны соединяются с узлами другой стороны каждого сегмента, таким образом образуются своеобразные автономные «микропроцессоры», управляющие органами одного сегмента. Такое строение нервной системы обеспечивает высокую надежность жизнедеятельности кольчатых червей, что позволяет им сохранять жизнь даже при расчленении тела червя на несколько частей. Мощный надглоточный узел, соединенный с подглоточным узлом, а через него и с брюшными узлами, свидетельствует о зарождении центральной нервной системы у этих животных. Для этих червей характерно многообразие рецепторов (хеморецепторы, фоторецепторы, рецепторы равновесия, осязания, болевые рецепторы и барорецепторы, позволяющие ощущать давление), и головной нервный узел у них крупный, он выполняет роль головного мозга. Развитая нервная система и мускулатура позволяют кольчецам вести разнообразную жизнь и использовать разные виды растительной и животной пищи. Для них характерны все типы рефлексов, они способны к условнорефлекторному обучению и обладают ассоциативной памятью, что позволяет им проявлять относительно сложное поведение.
Узловая нервная система в процессе эволюции получила дальнейшее развитие у моллюсков и членистоногих.
У моллюсков тело напоминает мышечный мешок, в котором обнаруживается нервных волокон, берущих начало от трех пар узлов. Цельные узлы являются сложным аппаратом и достигают наивысшего развития у головоногих моллюсков (кальмаров, осьминогов).
Нервная система членистоногих (особенно насекомых) развивалась в направлении усложнения и усовершенствования различных функций. У некоторых видов насекомых (перепончатокрылых) не только нервная система, но и органы чувств достигают вершины развития среди беспозвоночных животных. Таким образом, нервная система беспозвоночных способна не только обеспечивать различной сложности безусловно-рефлекторные двигательные акты, но и являться основой для некоторых форм научения.

70. Развитие головного мозга в эмбриогенезе позвоночных. Типы головного мозга позвоночных. Филогенетически обусловленные аномалии развития нервной системы у человека.
Головной мозг развивается из ростральной части нервной трубки. Бо́льшая часть головного мозга (95 %) является производной крыловидной пластинки.
Эмбриогенез мозга проходит через несколько стадий.
Стадия трёх мозговых пузырей — у человека в начале четвёртой недели внутриутробного развития ростральный конец нервной трубки формирует три пузыря: Prosencephalon (передний мозг), Mesencephalon (средний мозг), Rhombencephalon (ромбовидный мозг, или первичный задний мозг).
Стадия пяти мозговых пузырей — у человека в начале девятой недели внутриутробного развития Prosencephalon окончательно делится на Telencephalon (конечный мозг) и Diencephalon (промежуточный мозг), Mesencephalon сохраняется, а Rhombencephalon делится на Metencephalon (задний мозг) и Myelencephalon (продолговатый мозг).
В процессе формирования второй стадии (с третьей по седьмую недели развития) головной мозг человека приобретает три изгиба: среднемозговой, шейный и мостовой. Сначала одновременно и в одном направлении формируются среднемозговой и мостовый изгибы, потом — и в противоположном направлении — шейный. В итоге линейный мозг зигзагообразно «складывается».
При развитии мозга человека можно отметить определённое сходство филогенеза и онтогенеза. В процессе эволюции животного мира первым сформировался конечный мозг, а затем — средний мозг. Передний мозг является эволюционно более новым образованием головного мозга. Также и во внутриутробном развитии ребёнка сначала формируется задний мозг как самая эволюционно древняя часть мозга, а затем — средний мозг и потом — передний мозг. После рождения с младенческого возраста до совершеннолетия происходит организационное усложнение нейронных связей в мозге.
Типы мозга позвоночных.
Выделяют три типа головного мозга: ихтиопсидный, зауропсидный и маммальный.
Источник



