
Только осмотрофный способ питания у прокариот
Все перечисленные ниже признаки, кроме двух, используются для описания изображённой на рисунке клетки. Определите два признака, «выпадающие» из общего списка, и запишите в таблицу цифры, под которыми они указаны.
1) осмотрофный способ питания
2) размножение путём продольного деления
3) наличие сократительной вакуоли
4) наличие разнообразных пластид
5) способность к фагоцитозу
Признаки животной клетки:
1) эукариотическая клетка;
2) отсутствует клеточная стенка;
3) на наружной поверхности клеточной мембраны имеется гликокаликс, образованный олигосахаридами;
4) в наружной клеточной мембране присутствует холестерин;
5) мембранные органоиды: ЭПС, аппарат Гольджи, митохондрии, лизосомы, пероксисомы;
6) немембранные органоиды: рибосомы, клеточный центр (центриоли), микротрубочки, микрофиламенты;
7) отсутствуют пластиды (хлоропласты, хромопласты, лейкопласты), отсутствуют крупные центральные вакуоли;
8) запасной полисахарид – гликоген;
9) сократительные вакуоли (характерны для одноклеточных животных – простейших, выполняют функцию осморегуляции;
10) питательные вещества поступают в клетку путем фагоцитоза (для многоклеточных животных – путем заглатывания) – голозойный способ.
На рисунке изображено простейшее (одноклеточное животное) амёба (имеются ложноножки).
(1) осмотрофный способ питания – признак выпадает (у животных – голозойный способ питания, путем заглатывания или фагоцитоза; осмотрофный способ питания характер для клеток с клеточным стенками – бактериальной, грибной, растительной);
(2) размножение путём продольного деления – признак амёбы;
(3) наличие сократительной вакуоли – признак амёбы;
(4) наличие разнообразных пластид – признак выпадает (признак растительной клетки);
(5) способность к фагоцитозу – признак амёбы.
Осмотрофный (от др.-греч. ὄσμος — «толчок, давление» и τροφή — «питание») — питание без захвата твёрдых пищевых частиц — посредством транспорта (пассивного — осмоса, или активного) растворённых питательных веществ через поверхностные структуры клетки. Может использоваться как при гетеротрофном так и при автотрофном способе питания. Данный способ характерен для фотосинтезирующих растений, грибов и большинства микроорганизмов (исключая гетеротрофных простейших).
Источник
Типы питания прокариот
Прокариоты характеризуются многообразием типов питания в отличие от растений (автотрофы) и животных (ге- теротрофы). Для характеристики типов питания используют три критерия: источник углерода, источник энергии, донор электронов (водорода). По источнику углерода делят на ав- тотрофы и гетеротрофы, по источнику энергии – на фото- трофы (используют солнечный свет) и хемотрофы (получа- ют энергию за счет окислительно-восстановительных реак- ций), по донору – электроны (водорода) на литотрофы (неорганические соединения Н2, NH3, H2S, Fe2 + , CO и др.) и органотрофы (используют в качестве донора электрона ор- ганические соединения). Таким образом, выделяют 4 основных типа питания прокариот: фотолитоавтотрофы, фотоорганоавтотрофы, хемолитоавтотрофы, хемоорганоге- теротрофы.
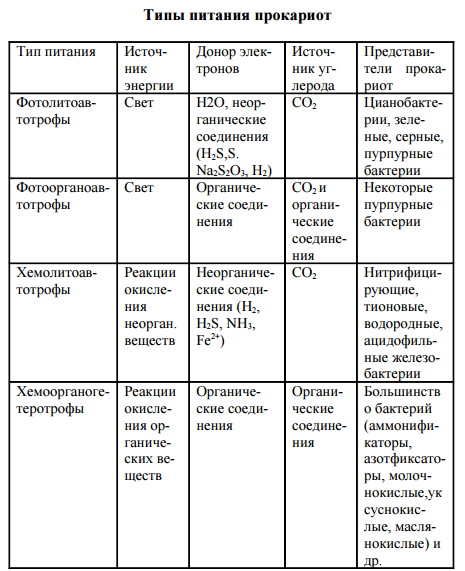
Фотолитоавтотрофы. Бактериальный фотосинтез (фоторедукция). В 1931 г. К. ван Ниль впервые доказал способность бактерий к фотосинтезу. В основе бактериального фотосинтеза лежит превращение световой энергии, поглощаемой фотосинтетическим пигментом, в биохимическую энергию макроэргических связей (АТФ) и далее использо- вание этой энергии для восстановления углекислого газа в процессе биосинтеза. У этих организмов есть пигмент бак- териохлорофилл. В клетках всех фотосинтезирующих бактерий содержатся фотосинтетические пигменты. К ним относятся особые хлорофиллы, получившие название бактериохлорофиллов а, в, с, d, и каротиноиды. По строению бактерио- хлорофиллы близки к хлорофиллу а растений. Так, бакте- риохлорофилл а отличается от хлорофилла а растений тем, что в первом его пирольном кольце в положении 2 стоит ацетильная группа СН3-СО- вместо обычной винильной группы СН2=СН-, а второе пирольное кольцо восстановле- но и содержит на два атома водорода больше. Содержание в клетке фотосинтезирующих прокариот четырех видоизменений бактериохлорофилла обусловли- вает более широкую полосу спектра поглощения световой энергии по сравнению с хлорофиллом растений. Предел спектра поглощения хлорофилла растений лежит в области 700-780 нм, в то время как фотосинтезирующие бактерии используют световую энергию длинноволновой части спектра до 1100 нм. Разница в спектрах поглощения хлоро- филлов растений и бактериохлорофиллов фотосинтезиру- ющих бактерий расширяет экологические ниши последних и позволяет фотосинтезирующим бактериям развиваться в водоемах под слоем водорослей. Помимо бактериохлорофиллов, в клетках фотосинтезирующих бактерий открыты более 20 дополнительных каро- тиноидных пигментов. Особенно разнообразны каротинои- ды в клетках пурпурных бактерий. Каротиноиды фотосин- тезирующих бактерий поглощают световую энергию ко- ротковолновой видимой области спектра с длиной волны 400-550 нм и передают эту энергию на бактериохлоро- филл. В клетке прокариот фотосинтетические пигменты нахо- дятся на инвагинациях ЦПМ – хроматофорах, визикулах (пузырьках), трубочках, тиллакоидах, ламеллах. Ван Ниль назвал этот процесс бактериальным фотосин- тезом, или фоторедукцией.
Для восстановления одной молекулы углекислого газа требуется 1 молекула АТФ (у растений 4 АТФ). В том и другом случае идет восстановление углекислого газа до углеводов. В Определителе Берджи (1974) фотосинтезирующие бактерии представлены тремя семействами (см. выше).
Фотоорганоавтотрофы Фотоорганоавтотрофы представлены немногочисленным семейством Rhodospirillaceae, включающим три рода: Rhodospirillum, Rhodopseudomonas и Rhodomicrobium. Бактерии-фотоорганоавтотрофы способны перестраивать свой обмен и одинаково успешно развиваться как на свету, так и в темноте, переходя соответственно от ана- эробного образа жизни к аэробному. На свету они ведут себя как фотоорганоавтотрофы: усваивая углекислый газ, они восстанавливают его в процессе фотосинтеза до углевода. В качестве доноров электронов несерные пурпурные бактерии используют различные органические вещества — сахара, спирты, органические кислоты, аминокислоты. Характер использования органических веществ различными фотосинтезирующими бактериями существенно раз- личается. Чаще всего органическое вещество выполняет единственную функцию донора электронов при фотоасси- миляции углекислого газа. Однако в некоторых случаях органическое вещество используется фотосинтезирующими бактериями не только в качестве донора электронов, но одновременно и как источник углерода. Попадая в темноту, пурпурные бактерии переходят к хемоорганогетеротрофному типу питания. При этом энергию для процессов жизнедеятельности они получают за счет реакций окисления органического субстрата по циклу Кребса. Непосредственным источником углерода и донором электронов для них являются органические соедине- ния субстрата. В клетках бактерий этой группы имеется универсальный набор дыхательных ферментов (НАД- и ФАД-дегидрогеназы и цитохромы), обеспечивающих им возможность перехода от анаэробного образа жизни на свету к аэробному в темноте и, соответственно, от авто трофного типа питания к гетеротрофному. Таким образом, среди разных групп прокариот нет, да и не может быть, резкой границы между различными типами питания.
ХемолитоавтотрофыХемолитоавтотрофы представлены микроорганизмами, способными в качестве основного источника углерода усваивать углекислый газ и синтезировать в клетке органические соединения, используя энергию реакций окисления неорганического субстрата. Для хемолитоавтотрофов неорганические вещества субстрата выступают донорами электронов в реакциях энергетического метаболизма и в процессе хемоассимиляции углекислого газа. Заслуга открытия процесса хемосинтеза принадлежит С. Н. Виноградскому. Он определил химизм как жизнь без органики. Им впервые была показана возможность образования органических веществ из неорганических в клетках бактерий, помимо процесса фотосинтеза. К хемолитоавтотрофам относится большинство видов нитрифицирующих, тионовых бактерий, некоторые виды из группы одноклеточных железобактерий и водородные бактерии. Хемолитоавтотрофные бактерии характеризуются специфичностью в отношении использования окисляемого субстрата. Нитрифицирующие бактерии для процесса хемоассимиляции углекислого газа получают энергию от окисления аммиака и нитритов. Процесс нитрификации проходит в две фазы. Первая фаза заключается в аэробном окислении аммиака до нитритов нитрозными бактериями родов Nitrosomonas, Nitrosococcus, Nitrosolobus, Nitrosospira: NН3 + 11 /202 = HN02 + Н2О + 274,7 кДж. Вторая фаза нитрификации предусматривает окисление нитритов в нитраты нитратными бактериями родов Nitrobacter, Nitrospina, Nitrococcus: HN02 + 11 /202 =НNО3 + 87,5 кДж. При процессах окисления неорганического субстрата выделяется сравнительно небольшое количество энергии, и усваивается она клеткой с низким КПД, всего 5-10%. Поэтому для получения энергии на процессы жизнедеятельности нитрифицирующим бактериям приходится перерабатывать огромное количество субстрата. К хемолитоав- тотрофам относится большинство видов тионовых бактерий: Thiobacillus denitrificans, Т. thiooxidans, Т. acidophilus, Thiomicrospira pelophila и др. Они ведут процесс хемоасси- миляции углекислого газа, получая энергию за счет окисления восстановленных или частично восстановленных соединений серы, сероводорода, элементарной серы, тиосульфата и сульфита. Конечным продуктом окисления обычно является сульфат.
Некоторые виды тионовых бактерий (Thiobacillus ferrooxidans) способны получать энергию за счет окисления не только соединений серы, но и закисного железа (Fe2+) (разрушение труб). Хемолитоавтотрофный тип питания характерен для некоторых одноклеточных ацидофильных железобактерий – Leptospirillum ferrooxidans, Thiobacillus ferrooxidans и представителей рода Sulfolobus. Оптимальный рост ацидофильных железобактерий наблюдается при рН ниже 4,5 (2-3). Они используют энергию окисления Fe2+ до Fе3+ для ассимиляции С02, который служит основным или единственным источником углерода. Реакции окисления железа сопровождаются незначительным выделением энергии, поэтому железобактерии перерабатывают большие количества субстрата: 2Fe2+ + 1/202 + 2Н+ = 2FеЗ+ + Н2О + 33 кДж. Хемолитоавтотрофные бактерии в природе являются геологическими агентами. Они принимают участие в процессах образования полезных ископаемых и осуществляют важнейшие звенья круговорота азота, серы, железа. К факультативным хемолитоавтотрофам относятся водородные бактерии, являющиеся представителями 20 различных родов – Рsеudоmоnаs, Аlcаligеnеs, Nocardia и др. Они способны осуществлять ферментативное окисление водорода кислородом воздуха с образованием воды. Водородные бактерии ферментом дегидрогеназой активируют молекулярный водород и далее используют его для получения энергии и в качестве донора электронов для восстановления углекислого газа до углевода: 6Н2 + 202 + С02 = (СН2О) + 5Н2О. Являясь факультативными хемолитоавтотрофами, водородные бактерии в качестве источника энергии и углеро- да способны использовать и различные органические со- единения (сахара, органические кислоты, спирты). Таким образом, водородные бактерии могут служить еще одним 87 примером возможного перехода микробной клетки от одного типа питания к другому – от хемолитоавтотрофного к хемоорганогетеротрофному. Бактерии, переходящие от одного типа питания к другому, получили название миксо- трофов.
Хемоорганогетеротрофы. К хемоорганогетеротрофам относится большинство прокариот. Источником углерода для них являются самые разнообразные органические соединения. Энергию для жизнедеятельности они получают за счет окислительно- восстановительных реакций органического субстрата, и донором электронов в реакциях метаболизма также выступают различные органические вещества. Хемоорганогетеротрофы наиболее широко распространены в природе. Им принадлежит роль санитаров нашей планеты, так как они ведут процессы минерализации самых разнообразных, подчас сложных органических веществ. Помимо органических соединений как источника углерода, хемоорганогетеротрофы нуждаются в углекислом газе для реакций карбоксилирования промежуточного обмена. Хемоорганогетеротрофные микроорганизмы подразделяют на сапрофитов и паразитов. Сапрофиты потребляют органические вещества опада. Паразиты живут за счет органических веществ живой клетки. Выделяют факультативных и облигатных паразитов. Факультативные паразиты развиваются на обычных органических средах, но, попадая в клетку-хозяина, переходят к паразитическому образу жизни. К ним относится большинство патогенных бактерий, вызывающих заболевания человека, – возбудители пневмонии, менингита, гонореи, дизентерии, брюшного тифа, сибирской язвы, коклюша, туберкулеза и др. Облигатные (строгие) паразиты развиваются исключительно за 88 счет органических веществ клетки-хозяина. Типичным примером облигатных паразитов являются риккетсии и вирусы. В основе всех вышерассмотренных типов питания прокариот лежат различные сочетания их энергетического и конструктивного метаболизма. Всем типам питания соот- ветствуют определенные группы прокариотных организ- мов. Основной массе бактерий присущ один вполне определенный (облигатный) тип питания. Однако среди многообразного мира прокариот встречается много видов бактерий, способных переходить от одного типа питания к другому. Так, среди цианобактерий, зеленых и пурпурных бактерий имеются виды, переходящие от фотолитоавто- трофного типа питания к фотоорганогетеротрофному. Некоторые виды обширного рода Thiobacillus способны переходить от хемолитоавтотрофного типа питания к хемоорга- ногетеротрофному. Эта особенность питания прокариот обусловливает широкие возможности существования микроорганизмов в различных условиях среды и значительно расширяет их экологические ниши.
Источник
Бактерии-прокариоты – живые «кирпичики» эволюции
Первые живые организмы (прокариоты), послужившие исходным материалом для эволюции жизни на Земле, имели очень примитивное строение. Бактерии относят к прокариотам, поскольку у них нет ядра и других органелл, присущих более развитым формам жизни.

Представители царства прокариотов
Название «прокариоты» происходит от древнегреческих слов «перед» и «ядро», т. е. это организмы, существовавшие еще до появления в клетках ядер. Это своеобразные предки эукариотов – видов, которые имеют оформленное клеточное ядро.
Прокариоты – это одноклеточные бактерии, в которых нет четко оформленного ядра клетки, ограниченного ядерной оболочкой, и дополнительных мембранных органоидов. Вместо этого прокариоты используют структуру, состоящую из ДНК (дезоксирибонуклеиновой кислоты), белков и РНК (рибонуклеиновой кислоты).
Исключение составляют фотосинтезирующие организмы, например, цианобактерии (сине-зеленые водоросли), которые имеют плоские цистерны – мембранная структура, обеспечивающая процесс жизнедеятельности клетки. Эта группа бактерий содержит зеленые и синие пигменты и участвует в процессе фотосинтеза, насыщая атмосферу кислородом.
Еще один представитель царства прокариотов – археи (лат. «древний, первозданный»). Эти одноклеточные бактерии интересны не только тем, что у них нет ядра, но и особенностями питания. Так, они могут существовать и находить пищу в самых экстремальных условиях – в горячих источниках и соленых озерах. Археи широко распространены в океанах, почве, болотах, даже в организме человека. Эти бактерии играют одну из ведущих ролей в круговороте углерода и азота на нашей планете.
Итак, к прокариотам относятся все бактерии, включая сине-зеленые водоросли и археи. Некоторые ученые относят к царству прокариотов и вирусы, но общего мнения на их счет пока не существует, так как некоторые биологи не считают вирусы живыми организмами.
Считается, что прокариоты появились около 3,5 млрд лет назад, и только через 2,5 млрд лет после них возникли первые эукариотные клетки. Все микроорганизмы, входящие в царство прокариотов, так или иначе способствовали созданию и поддержанию жизни на Земле.
Характеристика и строение
В среднем размер клетки-прокариота от 1 до 10 мкм. Мы уже знаем, что бактерии – это прокариоты, у которых нет клеточного ядра. Вместо него в клетке находится единственная крупная кольцевая или линейная молекула ДНК. Эта молекула содержит основную часть генетического материала клетки и называется нуклеоид («подобный ядру»). Прокариоты считают предками митохондрий и пластид – энергетических станций клеток эукариотов.
Нити ДНК и белки (гистоны) в прокариотических клетках не взаимодействуют, в отличие от клеток эукариот. Однако, по аналогии, комплекс ДНК и белков называют хромосомой. Хромосома прокариота крепится к мембране клетки, как правило, в одной точке. При удвоении хромосомы копии расходятся в разные стороны, образуя новые клетки, т.е. размножаются простым делением.
В отличие от эукариотов в этих микроорганизмах нет митохондрий, эндоплазматической сети и других органоидов. То есть как более примитивные организмы прокариоты не содержат тех мембранных структур, которые упорядочивают строение эукариотов.
Среда обитания прокариотов практически ничем не ограничена. Выживать в любых условиях бактериям помогают особенности их способов получения пищи:
- Осмотрофный – питание без захвата твердых частиц, т.е. поглощение питательных веществ, растворенных в окружающей среде, напрямую через поверхность клетки.
- Автотрофный – синтез органики из неорганических веществ, осуществляемый фотосинтезом (энергия света) или хемосинтезом (энергия химических реакций). Характерные представители – сине-зеленые водоросли.
Клетки прокариотов могут иметь различную форму: прямые или изогнутые палочки, шарики. Часто их клетки образуют колонии в виде нити или грозди, могут быть неподвижными или передвигаться с помощью жгутиков.
Эукариоты и прокариоты различаются настолько сильно, что ученые-систематики относят их к самым крупным подразделениям живой природы – царствам.
Поставщики кислорода и азота
Одним из ярких представителей царства прокариотов являются цианобактерии (сине-зеленые водоросли). Эти бактерии считают наиболее близкими к первым ископаемым микроорганизмам, найденным учеными. Возраст предков сине-зеленых водорослей составляет примерно 3,5 млрд лет.
Сине-зеленые водоросли – это единственные бактерии, способные выделять кислород. Правда, для самих водорослей это побочный продукт, но для существования жизни на нашей планете это одно из основных условий.
Сине-зеленые водоросли очень сложно организованы и имеют различные формы и размеры. Эти микроорганизмы способны даже менять свой цвет от темно-синего до светло-зеленого в зависимости от спектрального состава света.
Кроме способности выделять кислород, сине-зеленые водоросли имеют еще одну очень полезную черту – они могут связывать атмосферный азот и делать его доступным для других живых организмов. Эта последняя способность делает сине-зеленые водоросли незаменимыми для всех растений, которые не могут самостоятельно добывать азот из окружающей среды.
Вирусы – живая или неживая материя?
Вирусы поражают все типы живых организмов: растения, животных, бактерии, даже сами вирусы. Вирусы бактерий называют бактериофагами, а вирусы других вирусов – вирусы-сателлиты (простите за тавтологию).
Вирусы относят к неклеточной форме жизни. Они занимают положение между живой и неживой материей. У вирусов нет цитоплазмы и других клеточных органоидов. Отсутствие собственного обмена веществ роднит вирусы с неживой природой.
Вместе с тем вирусы отлично живут и размножаются внутри клеток других организмов, что делает их схожими с живыми существами. Но вне клетки-хозяина вирус существует только в кристаллической форме.
При изучении особенностей строения и поведения вирусов становится понятно, почему наука никак не определится с их принадлежностью.
Царство грибов
Грибы в систематике живой природы стоят особняком. Долгое время считалось, что грибы относятся к растениям, не содержащим хлорофилл. Однако современная наука выделяет грибы в отдельное царство, сочетающее признаки растений и животных. Исследования показали, что грибы подразделяются на несколько больших групп, некоторые из них даже не являются родственными.
Отсутствие хлорофилла роднит грибы с животными, так как они используют гетеротрофный способ питания. То есть грибы поглощают готовые органические соединения, растворенные в окружающей среде, и выделяют мочевину, как животные. В этом их отличие от растений, которые вырабатывают пищу с помощью процесса фотосинтеза.
В то же время грибы имеют клеточные стенки, возможность неограниченного роста и не способны передвигаться, как и растения.
Грибы относят к эукариотам, но по размеру генома они приближаются к прокариотам. То есть в клетках грибов есть ядро с ДНК-структурой, но процесс передачи генной информации может происходить и при помощи вирусов. Размер генома грибов и размер хромосом значительно меньше, чем у других видов эукариотов.
Точное определение царства грибов отсутствует, но их изучение необходимо для понимания эволюции жизни на Земле.
Горизонтальный перенос генов
Впервые этот процесс был описан в Японии в 1959 г. Горизонтальный перенос генов имеет широкое распространение в царстве прокариотов и даже у некоторых эукариотных клеток. В отличие от привычного нам вертикального переноса генов (от предка к потомку) смысл горизонтального процесса заключается в передаче генного материала организму, не являющемуся потомком исходной клетки. Именно такой принцип использует современная генная инженерия.
Открытие такого обмена генным материалом повлияло на учение об эволюции жизни. Раньше ученые считали, что виды не могут обмениваться наследственной информацией между собой. Однако прокариоты передают генную информацию как самостоятельно, так и с помощью вирусов-бактериофагов. То есть в прошлом нашей планеты, населенной древними микроорганизмами, существовал массированный перенос генетической информации, что не могло не повлиять на то, каким путем пошел процесс эволюции на Земле.
Образование высшее филологическое. В копирайтинге с 2012 г., также занимаюсь редактированием/размещением статей. Увлечения — психология и кулинария.
Источник



