
- Способы транспорта вещества через клеточную мембрану
- Биология. 11 класс
- § 11. Поверхностный аппарат клетки
- Активный и пассивный транспорт через мембрану
- Виды транспорта через мембрану
- Пассивный транспорт через мембрану
- Диффузия
- Отличие каналов и пор
- Облегченная диффузия
- Активный транспорт
- Первично-активный транспорт
- Вторично-активный транспорт
- Экзоцитоз и эндоцитоз
- Транспорт через несколько слоев клеток
Способы транспорта вещества через клеточную мембрану
• Существует два основных типа мембранных транспортных белков: каналы и переносчики
• Ионные каналы катализируют быстрый и избирательный транспорт ионов по электрохимическому градиенту
• Транспортеры и насосы представляют собой белки-переносчики, которым необходима энергия для транспорта веществ против электрохимического градиента
• В каждой клетке несколько различных мембранных транспортных белков действуют как единое целое
Некоторые из транспортных белков находятся в плазматической мембране, в то время как другие входят в состав мембраны клеточных органелл. Для поддержания определенного состава внутриклеточной среды и клеточных компартментов необходимо, чтобы транспортные белки обладали избирательностью по отношению к определенным метаболитам. В зависимости от характера процесса транспорта мембранные транспортные белки подразделяются на две группы: каналы и переносчики.
Канальные белки содержат поры, через которые при открытом канале с высокой скоростью проходят метаболиты. Белок-переносчик связывает метаболит, находящийся по одну сторону мембраны, после чего конформация его меняется, и метаболит высвобождается с другой стороны мембраны.
Существует несколько типов белков, образующих каналы в мембранах. Порины, присутствующие в клетках некоторых прокариот, в митохондриях и в нексусах, где соединяется цитоплазма соседних клеток, обеспечивают прохождение метаболитов в соответствии с размером их молекул.
К более селективным каналам относятся комплексы ядерных пор и каналы, обеспечивающие перемещение белков через мембрану эндоплазматического ретикулума. В статьях на сайте будут рассмотрены ионные каналы и аквапорины, через которые осуществляется селективный трансмембранный перенос ионов и молекул воды соответственно. К настоящему времени описано более 100 различных типов канальных белков. Все они обладают следующими характеристиками:
• селективностью по отношению к переносимому метаболиту,
• высокой скоростью переноса метаболита и
• воротным механизмом, регулирующим транспорт метаболитов.
Участок канального белка, через который осуществляется трансмембранный транспорт, называется порой канала. Канал может находиться в нескольких различных конфигурациях. Некоторые каналы состоят из одного белка, трансмембранные сегменты которого образуют пору. Другие существуют в виде олигомеров, состоящих из идентичных или разных субъединиц, которые вместе образуют пору.
Наконец существуют каналы, состоящие из двух и более субъединиц, каждая из которых сама образует пору. Часто комплексы олигомеров каналов регулируются или позиционируются на определенных мембранах с участием дополнительных субъединиц.
Большинство канальных белков обладают высокой селективностью в отношении определенных метаболитов, например ионов натрия (Na+), калия (К+), кальция (Са2+), хлорида (Сl-) или воды. Другие канальные белки являются неселективными катионными или анионными каналами. Как будет показано далее, при рассмотрении свойств специфических канальных белков, их поры характеризуются определенными структурными особенностями, т. е. являются селективным фильтром, который позволяет им проявлять избирательность по отношению к различным метаболитам.
![]() Каналы и переносчики представляют собой два основных типа белков мембранного транспорта.
Каналы и переносчики представляют собой два основных типа белков мембранного транспорта.
Растворенные метаболиты с максимальной скоростью диффундируют через пору каналов,
а переносчики связывают метаболит с одной стороны мембраны, затем претерпевают конформационные изменения и высвобождают его с другой стороны.
Этот процесс протекает значительно медленнее.
Направление потока ионов растворенных веществ через канал определяется электрохимическим градиентом. Иными словами, метаболиты движутся через канал в энергетически-выгодном направлении, т. е. по электрохимическому градиенту. Например, для покоящейся клетки, схематически изображенной на рисунке ниже, существует направленный в клетку ионный поток через Na+-, Са2+-, Cl—каналы, и противоположный, проходящий через К+-каналы.
Так как других источников энергии, кроме электрохимического градиента, в данном случае не используется, то такой тип транспорта называется пассивным. Канальные белки обеспечивают высокую скорость транспорта. Так, для ионных каналов скорость составляет 108 в 1 с, что близко к максимальной скорости диффузии ионов в водной среде.
Транспортные мембранные белки контролируют различные клеточные функции при участии воротного механизма. В основе этого механизма лежит индукция конформационных изменений в белке в ответ на воздействие определенного стимула. Эти изменения приводят к открытию или закрытию канала. Например, открытие и закрытие ионных каналов могут регулироваться лигандами, величиной электрического потенциала, механическими или температурными воздействиями. Активация каналов может происходить очень быстро, что делает их идеальным инструментом обработки поступающих в клетку сигналов.
Например, передача нервного импульса зависит от электрического заряда, который создается ионным каналом в мембране. Каналы также играют важную роль в регуляции объема клетки и внутриклеточного pH, в транспорте солей и воды через эпителиальные клетки, в закислении внутриклеточных органелл и во внеклеточной передаче сигналов.
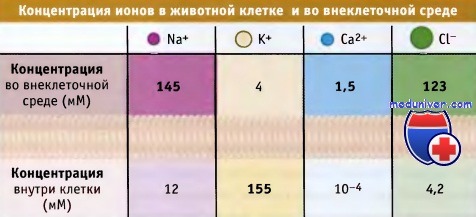
 Транспортеры и насосы представляют собой два основных типа белков-переносчиков.
Транспортеры и насосы представляют собой два основных типа белков-переносчиков.
Транспортеры делятся на три типа, каждый из которых осуществляет один из типов переноса: унипорт, симпорт, или антипорт.
Показано направление электрохимического градиента через мембрану.
В зависимости от типа белка-переносчика транспорт может происходить или в направлении градиента (от высокой концентрации к низкой), или против него.
При транспорте субстратов против градиента концентрации, белки-переносчики используют свободную энергию электрохимического градиента, АТФ или других источников. Поскольку при этом происходит потребление энергии, процесс носит название активный транспорт. Белки-переносчики подразделяются на две группы, транспортеры и насосы.
Для транспортировки субстратов через мембрану транспортеры используют энергию электрохимического градиента. Они подразделяются на унипортеры, симпортеры (или котранс-портеры) и антипортеры (обменники). Для энергозависимого транспорта метаболитов мембранные насосы используют энергию, например высвобождающуюся при гидролизе АТФ. По сравнению с канальными белками, переносчики характеризуются более медленной скоростью транспорта, порядка 1000 молекул в 1 с.
Различают два типа активного транспорта, первичный и вторичный. Белки-переносчики, осуществляющие первичный активный транспорт, в качестве источника энергии используют АТФ и транспортируют метаболиты против электрохимического градиента. Таким образом, они помогают поддерживать градиент концентрации метаболитов по обеим сторонам мембран клетки. Са2+-АТФаза и Na+/К+-АТФаза являются характерными примерами транспортных насосов, которые управляют процессом первичного активного транспорта.
Белки-переносчики, которые участвуют во вторичном активном транспорте, непосредственно не используют энергию АТФ. Вместо этого они используют свободную энергию электрохимического градиента, который возникает за счет первичного активного транспорта. Вторичный активный транспорт происходит с участием симпортеров и антипортеров. Механизмы транспорта метаболитов представлены на рисунке ниже.
В клетке все типы мембранных транспортных белков-каналы, транспортеры и насосы-функционируют совместно. Таким образом, работа какого-либо одного типа транспортного белка зависит от работы других. В данной главе мы рассмотрим несколько примеров такого взаимодействия транспортных белков. Например, ионный градиент по обеим сторонам мембраны поддерживается за счет сложных взаимодействий между различными типами транспортных белков. Для правильного функционирования эпителиальных клеток, например почек, слизистой кишечника и легкого, необходимо протекание транспортных процессов с участием различных ионов и метаболитов. Мы также рассмотрим роль нарушений процессов транспорта в развитии различных заболеваний.
![]() Транспорт растворенных метаболитов через мембрану может быть пассивным или активным.
Транспорт растворенных метаболитов через мембрану может быть пассивным или активным.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
Биология. 11 класс
§ 11. Поверхностный аппарат клетки
Обязательным компонентом поверхностного аппарата любой клетки является цитоплазматическая мембрана (плазмалемма). Она отделяет и защищает внутреннее содержимое клетки от внешней среды. Важнейшим свойством цитоплазматической мембраны является избирательная проницаемость. Различные вещества проходят через плазмалемму с разной скоростью, а для некоторых она практически непроницаема. Таким образом, цитоплазматическая мембрана обеспечивает обмен веществ между клеткой и внеклеточной средой и постоянство химического состава клетки.
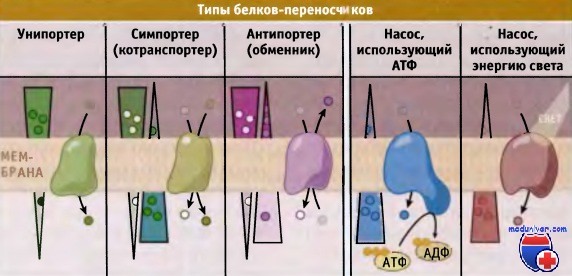
Химический состав и строение плазмалеммы. Цитоплазматическая мембрана состоит преимущественно из липидов и белков. В состав плазмалеммы также входят углеводы, которые являются компонентами сложных липидов и белков — гликолипидов и гликопротеинов. Толщина цитоплазматической мембраны составляет около 7,5 нм.
Около половины массы плазмалеммы составляют липиды, представленные в основном фосфолипидами. Они формируют основу строения мембраны — липидный бислой (двойной слой), в котором гидрофобные хвосты молекул погружены внутрь, а гидрофильные головки располагаются снаружи (см. рис. 6.3). Кроме фосфолипидов, в состав двойного слоя входят и другие липиды например стероиды.
*С липидным бислоем связаны белки. Молекулы интегральных белков пронизывают двойной слой липидов насквозь. Полуинтегральными называют белки, погруженные лишь в один из двух липидных слоев (внешний либо внутренний). Периферические белки примыкают к поверхности бислоя с наружной или внутренней стороны (рис. 11.1).*
Мембранные липиды и белки связаны между собой не ковалентными связями, а за счет гидрофобных и электростатических взаимодействий. В связи с этим молекулы данных веществ могут двигаться вдоль плоскости мембраны. Таким образом, плазмалемма является динамичной структурой — молекулы белков перемещаются в подвижном, текучем липидном бислое, как в жидкости. *Данная модель организации мембраны получила название жидкостно-мозаичной. Она была предложена американскими биологами Дж. Сингером и Г. Николсоном в 1972 г.* Свойство текучести обусловливает пластичность плазмалеммы, благодаря чему она способна, например, быстро восстанавливать свою целостность после незначительных повреждений.
*Движение в плоскости плазмалеммы происходит достаточно легко. Однако переход белков с одной стороны мембраны на другую (так называемый флип-флоп переход) практически невозможен. Подобное перемещение липидов происходит, но очень редко, при участии специальных белков флиппаз. Поэтому состав наружного и внутреннего слоев плазмалеммы различается.*
*Текучесть мембраны зависит от ее липидного состава и температуры. С увеличением содержания остатков ненасыщенных жирных кислот или температуры подвижность молекул возрастает. Стероиды , например холестерин в организме животных, регулируют текучесть плазмалеммы и придают ей жесткость. Стероиды располагаются в свободном пространстве между гидрофобными хвостами фосфолипидов, ограничивая подвижность их молекул друг относительно друга (см. рис. 11.1). Таким образом, стероиды обеспечивают стабильность мембраны и поддержание ее структуры. Кроме того, они регулируют проницаемость плазмалеммы.*
В клетках эукариот, кроме плазмалеммы, имеются внутренние мембраны, ограничивающие ядро и мембранные органоиды. Для всех биологических мембран характерен общий план строения — их основу составляет липидный бислой, с которым связаны молекулы белков.
Источник
Активный и пассивный транспорт через мембрану
Ты уже шаришь какое строение у биологических мембран, так что можно переходить к тому, как вся эта система работает. И начнем мы с транспорта веществ через мембрану. Довольно важная тема, без нее мы не поймем, как формируется потенциал действия, как в клетку попадают энергетические субстраты и вода. Так что присаживаемся поудобнее и поехали!
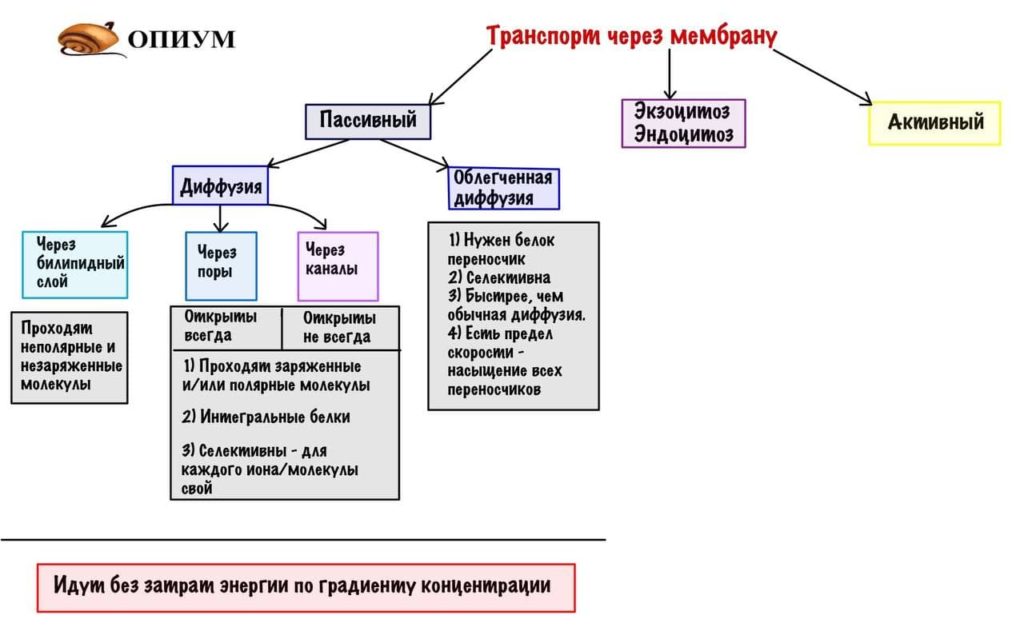
Виды транспорта через мембрану
Начнем с небольшой классификации. Транспорт можно разделить на пассивный и активный (никаких шуток про геев мне тут), такое разделение основано на затратах энергии. При пассивном транспорте — затрат энергии нет, а при активном транспорте — есть. Это может быть энергия заключенная в АТФ, либо энергия градиента концентрации. Не пугаемся, дальше все будем разбирать подробнее. Еще есть особенный транспорт — экзоцитоз и эндоцитоз (транспортируются макромолекулы), их скорее можно отнести к активному транспорту, но мы рассмотрим их отдельно.
Пассивный транспорт через мембрану
Здесь всего два вида — простая диффузия и облегченная диффузия. В чем отличие? При облегченной диффузии для молекулы, которая будет проходить через мембрану нужен проводник — белок переносчик. Для простой диффузии же переносчик не требуется, она и сама справляется.
Диффузия
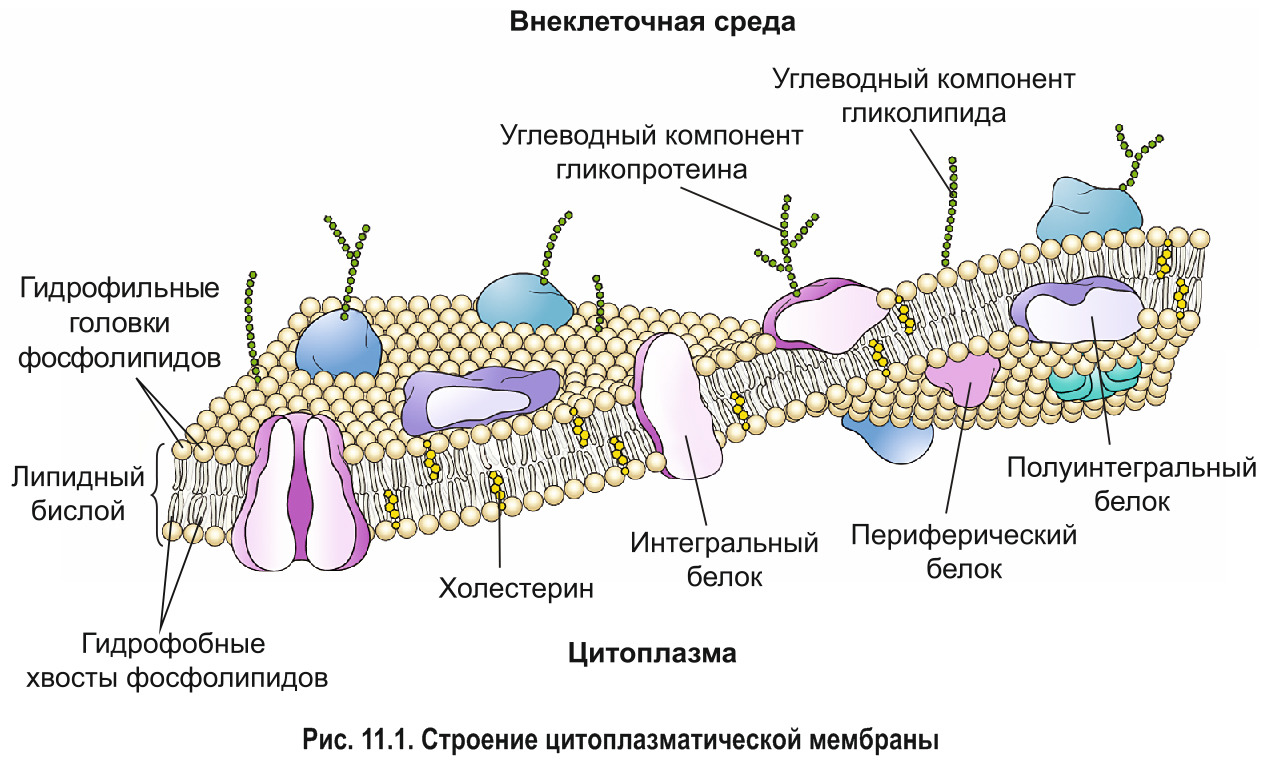
Она идет по градиенту концентрации — если на одной стороне мембраны вещества много, то оно будет стремиться перейти на другую сторону. На самом деле диффузия зависит не только от градиента концентрации — еще на нее влияет заряд мембраны и частиц, которые пытаются пройти через мембрану, и давление.
Вспоминаем, что основа мембраны — это амфифильные липиды. Если вещество растворимо в липидах, то оно без проблем перейдет на другую сторону. Лучше всего растворимы в липидах неполярные и незаряженные молекулы (CO2,O2, стероидные гормоны). Но через мембрану могут проходить полярные молекулы и незаряженные (тонированные), но здесь все решает размер. Вода пройдет через мембрану, а вот глюкоза — нет. Вода хоть и проходит через мембрану, но хуже чем неполярные и незаряженные молекулы, поэтому для нее есть специальные поры (аквапорины). Кстати, возможно аквапорины еще нужны потому, что вода в растворе переходит в ион гидроксония.
А вот совсем не могут пройти заряженные и полярные молекулы — ионы натрия, калия, гидроксония. Поэтому простую диффузию разделим еще на два варианта:
1) Диффузия жирорастворимых веществ. Молекула растворима в мембране, то есть она неполярная и незаряженная. Она спокойно проходит через мембрану. Избранные молекулы проходят так — O2, CO2, стероидные и тиреоидные гормоны.
2) Но что делать простым пацанам с района, которые не растворимы в жирах? Не парьтесь, для них есть интегральные белки — они образуют каналы и поры. Это просто трубы. Эти каналы очень избирательны, например, у ионов K + , Na + и молекул воды свои собственные каналы, которые не пропускают другие вещества. Это связано с их строением, так у натриевого канала очень много отрицательных групп, которые буквально вытягивают натрий. Поры же пропускают молекулы только определенного размера (если меньше, то пройдет).
Отличие каналов и пор
Отличие каналов и пор в том, что первые открыты не всегда, а вторые постоянно. У натриевых и калиевых каналов есть шляпка (или ворота), которая открывается двумя способами — в зависимости от типа клетки. Первый способ — изменение электрического заряда мембраны (или ее потенциала). Потеря отрицательного заряда клеткой ведет к открытию натриевых каналов — это важно для потенциала действия. Второй способ — действие химического вещества. Есть ацетилхолиновые каналы, которые открываются под действием(угадайте сами чего)… Так нервные клетки передают сигнал о сокращении мышцам.
Представьте два входа в клуб и один отдельный выход из него. Перед каждым входом стоит фейсер, но в первом нет двери — там просто дверной проем, а у второго и третьего дверь есть и для нее нужен ключик. Первый вход — это пора, молекула воды подходит к нему и фейсер пропускает ее, но иногда в толпе воды может затеряться другая молекула, например, мочевина. Фейсер по ошибке пропускает ее, ну а что — за всеми не углядишь, и она попадает внутрь клетки. Ко второму входу подходит ион натрия, фейсер его пропускает, но войти он не может пока дверь не откроют ключом — изменением потенциала или ацетилхолином. У выхода из клуба тоже стоит фейсер, вот такой вот странный клуб, к нему подходит ион калия и ждет пока дверь откроется таким же ключом. Фейсеры — это часть канала или поры, которая отвечает за узнавание ионов и молекул, а дверь или проем — это сам канал. Ну вы поняли.
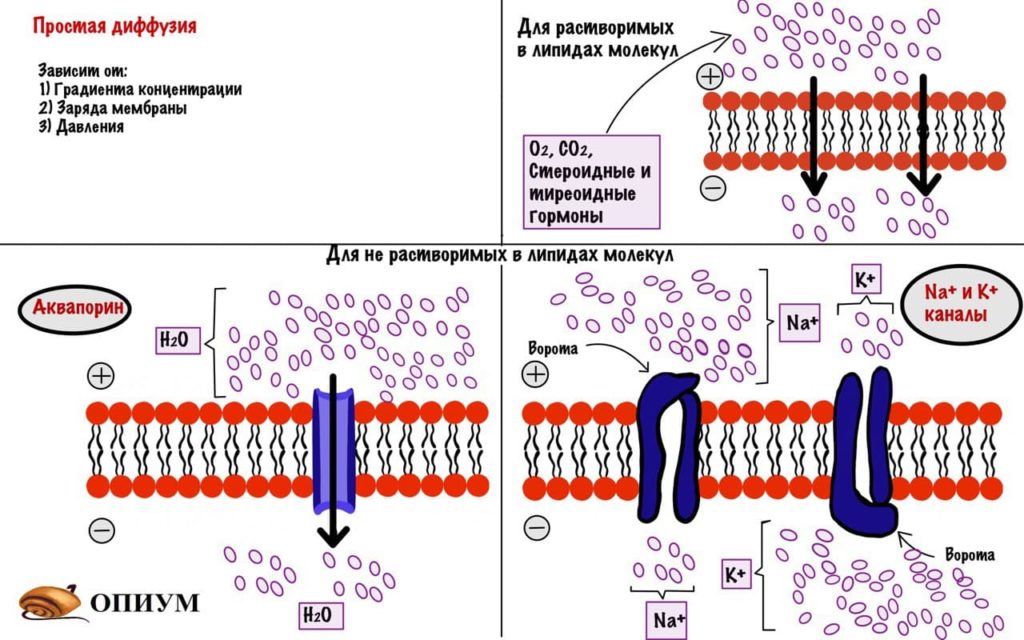
Почему клеткам просто не забить: убрать фейсеров и держать двери постоянно открытыми? Смотрите — внутри клетки много калия, но мало натрия. В межклеточном веществе наоборот, очень много натрия и мало калия. А это значит, что калий стремится сбежать из клетки, а натрий войти в клетку.
Во-первых, такая разница ионов создает заряд внутри клетки — отрицательный, если бы каналы были постоянно открыты, то такой разницы бы не существовало, клетка стала бы незаряженной. Что не очень хорошо, так как она не сможет создать потенциал действия. Во-вторых, натрий это любимчик воды и если в клетке его будет много, то будет много и воды. Клетка просто лопнет от такой тусовки.
Облегченная диффузия
Здесь молекуле, которая идет тоже по градиенту концентрации, необходим переносчик. Все это из-за того, что молекула слишком большая для перехода через мембрану самостоятельно. Переносчик — это интегральный белок, который пронизывает мембрану, у него тоже есть фейсер (только здесь это участок связывания). При взаимодействии молекулы с переносчиком — он изменяет свою структуру (конформационные изменения белка) и переносит молекулу в клетку, а затем возвращается обратно.
Такой механизм характерен для переносчиков глюкозы — ГЛЮТов в жировой и мышечной ткани. Однако ГЛЮТы не всегда находятся на поверхности клетки, а только после еды — повышенный уровень глюкозы в крови вызывает секрецию инсулина из Б-клеток островков Лангерганса. Инсулин действует на жировую и мышечную ткань и взывает к ГЛЮТам, которые встраиваются в мембрану. Ой, как-то на автомате получилось. Еще таким способом транспортируются аминокислоты.
Смотрите, еще один прикол. Эти переносчики могут работать в обе стороны, все зависит от градиента концентрации глюкозы. Если ее будет слишком много в клетке, то они могут выкидывать ее в кровь. Прикольненько?
Если интересно, что быстрее: диффузия или облегченная диффузия, то вот график. Видим, что вначале быстрее облегченная диффузия, а потом обычная. Почему? Просто белки могут связать только одну молекулу, когда молекул глюкозы становится очень много, то все переносчики связаны с ней. Наступает насыщение переносчиков, и они не могут быстрее работать. Диффузия же не зависит от переносчиков, но она немного медленнее.
Пассивный транспорт все, поэтому давайте суммируем все и добавим в нашу начальную схему.
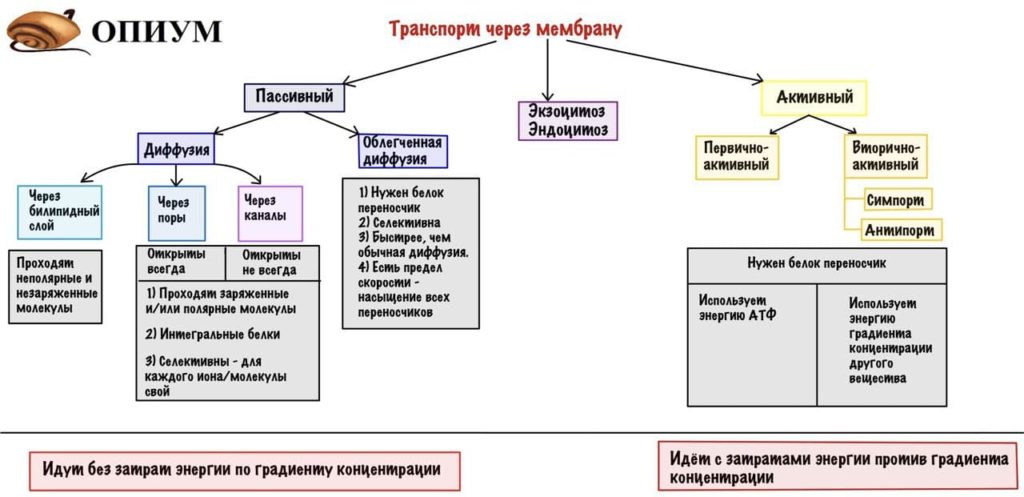
Активный транспорт
Здесь для переноса вещества через мембрану необходимо приложить энергию. Но зачем, а главное почему? Потому что такой транспорт идет против градиента концентрации, а без прикладывания энергии молекулу или ион просто не вытолкнуть. Разделяется на два варианта: первично-активный транспорт и вторично-активный транспорт, отличие между ними поймете чуть ниже.
Первично-активный транспорт
Здесь для того, чтобы перенести молекулы/ионы вещества на другую сторону мембраны используется энергия молекул АТФ. Классический вариант — натрий-калиевый насос. Этот насос представляет из себя белок, а именно фермент — АТФазу (помните, что «не все белки — ферменты, но все ферменты — белки» — десятая заповедь от кафедры биохимии). Занимается тем, что переносит ионы натрия из клетки, а ионы калия внутрь клетки. То есть работает против градиента концентрации, ведь натрия очень много вне клетки, а калия наоборот мало.
У насоса есть участки связывания — два для калия и три для натрия. Состоит из двух субъединиц — альфа и бета, альфа это и есть переносчик, а бета похоже якорит его в мембране. На один цикл: переноса трех ионов натрия из клетки и двух ионов калия внутрь клетки, требуется одна молекула АТФ. Как видим, этот насос создает разницу потенциалов, так как в обмен на три заряженных иона внутрь клетки поступает только два — этому пареньку мы обязаны за отрицательный заряд внутри клетки. Действует такой насос во всех клетках, он не дает клетке лопнуть из-за избытка натрия (вспоминаем про воду).
Кроме такого насоса есть еще несколько — Ca ++ и H + — АТФазы. Избыток кальция вредит клетке, так как он может запустить апоптоз. Водородный насос действует в париетальных клетках желудка и дистальном отделе канальца нефрона — в первом случае он создает кислую среду в желудке для функционирования пепсина. Да и вообще, из внешней среды поступает много всякой заразы, которой неприятно встречаться с кислотой. Во втором случае насос перемещает ионы водорода в просвет канальца. Полезная штука, а то прикинь — позанимался спортом и умер от ацидоза, не круто.
Вторично-активный транспорт
Тут одна молекула идет по градиенту концентрации и энергия, которая создается ей, используется для переноса другой молекулы. Представляете, сколько всего ионов натрия во внеклеточной жидкости? Вот и я не представляю, но очень много, а в клетке же наоборот его очень мало. Такая разница создает просто огромную энергию, которая идет на работу белка переносчика. Этот белок переносчик, как вы уже поняли — интегральный белок и имеет два участка связывания. Эти участки могут находиться на одной стороне белка или на разных. Поэтому такой транспорт можно разделить на два варианта:
1) Молекула, которая идет против градиента концентрации, переносится в одну сторону с молекулой, которая идет по градиенту концентрации. Это называется котранспорт (или симпорт). Так переносятся молекулы глюкозы и аминокислот из кишечника и канальцев нефрона. Натрий идет по градиенту концентрации внутрь клетки и захватывает с собой глюкозу или аминокислоты. Тут ты можешь сказать : «Чет странно, ведь в кишке много глюкозы после еды, почему она идет против градиента?». И да, это верно, в кишечнике много глюкозы. Но клеток очень много, а глюкоза растянута по всей поверхности кишки. Вот и получается, что в кишке ее много, но возле каждой клетки маловато. Такая же тема с аминокислотами.
2) Молекула идет против градиента концентрации, но не в одну сторону с переносимым по градиенту концентрации веществом — контртранспорт (или антипорт). Так происходит транспорт ионов водорода в проксимальных канальцах нефрона: водород попадает в просвет канальца, а натрий внутрь клетки.
Заметили кое-что? Клетка всегда чего-то боится: потерять или перебрать. Не всосать глюкозу и аминокислоты в кишечнике, либо смыть их в унитаз. И здесь она работает не только на свое благо, а на благо всего организма. Ведь ей не очень и нужна эта глюкоза, в ней ее достаточно, но она заботится не только о себе. А говорят, что коммунизм не построить , а он уже существует в организме каждого из нас. Ну это так, просто к слову пришлось… Перебрать же она боится, потому что из-за этого погибнет — поэтому натрий-калиевый насос работает постоянно, как и кальциевый.
Ну что сведем все это опять в нашу табличку?
 Если не очень хорошо видно, то в конце есть файл со всеми схемами. Извиняйте.
Если не очень хорошо видно, то в конце есть файл со всеми схемами. Извиняйте.
Все что мы разбирали до этого относится к небольшим по размерам молекулам, а что делать с большими? Для этого есть две легенды, о которых ниже.
Экзоцитоз и эндоцитоз
Начнем с экзоцитоза и сделаем это на каком-нибудь примере. Пусть это будут пищеварительные ферменты в поджелудочной железе. Синтезировала значит клетка липазу, но она ведь внутри клетки — это значит проку от нее мало. Нужно ее как-то переместить в проток поджелудочной железы, хорошо было бы использовать белок переносчик. А тут проблемка. Липаза слишком большая — ее не засунуть в белок переносчик. Но ничего — у клетки есть выход.
Все ферменты, белки плазмы, пептидные гормоны и так далее, синтезируются в упаковке — пузырьке (по строению он амфифильный). Оно и правильно, представьте — липаза попадает в цитоплазму клетки и просто переваривает ее. Эти пузырьки направляются к мембране, сливаются с ней и попадают в кровь, межклеточное вещество или проток поджелудочной железы. В общем куда им надо, туда они и попадают.
Теперь эндоцитоз. Все тоже самое только наоборот — это мое лучшее объяснение… Ладно, шутки кончились. На клеточной мембране есть определенный участок с рецепторами — окаймленная ямка. На рецепторах накапливаются макромолекулы, а потом ямка погружается в клетку и охватывает их, образуя пузырек. Этот пузырек направляется к лизосоме, где из него образуются мономеры. Эти мономеры клетка использует по своему усмотрению. Посмотрите картинку и все поймете, базарю.
Таким способом идет фагоцитоз лейкоцитами, а еще так в клетку попадают липопротеиды низкой плотности — это переносчики холестерина и жирных кислот.
Транспорт через несколько слоев клеток
Буквально пару слов. Разберем на примере кишки — там несколько слоев (три, ну ладно — четыре, если с подслизистой). Через все должна пройти глюкоза, но как? Это похоже на эстафету: сначала из кишечника вторично-активным транспортом глюкоза попадает в клетку, потом в следующую клетку уже по облегченной диффузии. Так она доходит до крови, а дальше уже идет по своим делам. Всё!
Хочешь задать вопрос, похвалить или наговорить гадостей? Тогда залетай в телегу. Там ты сможешь предложить новый формат или разбор темы. А если серьёзно, то эти статьи пишутся для вас, поэтому мне важна обратная связь.
Источник



