
Способы транспорта веществ через плазмолемму фагоцитоз пиноцитоз
• Для захвата материала из окружающей среды клетка использует несколько различных механизмов
• В последующих превращениях захваченного материала важную роль играют ферменты деградации и низкое значение pH в эндосомах и лизосомах
Эндоцитоз — процесс, при котором эукариотическая клетка захватывает материал из окружающей среды при образовании везикул на плазматической мембране. Во многом являясь противоположностью экзоцитоза, эндоцитоз обладает несколькими функциями:
• поглощение питательных веществ;
• регуляция экспрессии белков на клеточной поверхности, например рецепторов гормонов и переносчиков глюкозы, что дает возможность клетке контролировать процесс захвата лигандов;
• захват и деградация внеклеточного дебриса;
• восстановление мембраны, встроенной в плазматическую мембрану при секреции.
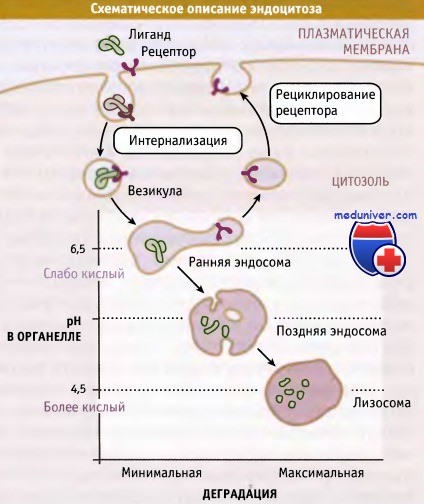 Среди органелл, участвующих в эндоцитозе, существует градиент pH и активности ферментов деградации.
Среди органелл, участвующих в эндоцитозе, существует градиент pH и активности ферментов деградации.
При эндоцитозе макромолекулы или рециклируют в плазматическую мембрану, или разрушаются.
Наряду с этим, процесс эндоцитоза используется для проникновения в клетку патогенными организмами, такими как бактерии, простейшие и вирусы.
При эндоцитозе образующиеся на плазматической мембране везикулы сливаются с органеллами. Эти органеллы обладают двумя важными особенностями: в люмене у них поддерживается кислая среда, и находятся протеолитические ферменты, оптимальная активность которых проявляется при кислых значениях pH.
Органеллы характеризуются различной степенью кислотности среды и различным содержанием ферментов деградации. Они подразделяются на ранние и поздние эндосомы и лизосомы, которые обладают широким спектром протеолитической активности соответственно от минимального в эндосомах до максимального в лизосомах.
В лизосомах находятся ферменты деградации, активные по отношению практически ко всем биополимерам (белкам, липидам, углеводам, РНК, ДНК), попадающим в них путем эндоцитоза. Хотя в течение долгого времени лизосомы рассматривались как терминальные органеллы на пути эндоцитоза, существуют данные о том, что они иногда могут сливаться с плазматической мембраной.
Протонный насос, функционирующий за счет энергии АТФ, называется вакуолярной АТФазой (v-АТФаза) и переносит ионы Н+ из цитозоля в люмен определенных органелл. При этом pH внутри органелл, по сравнению с цитозолем, уменьшается. Значение pH цитозоля составляет 7,4. Внутри ранних эндосом поддерживается слегка кислое значение pH (6,5-6,8), а в поздних эндосомах и в лизосомах значение pH достигает 4,5.
Величина pH в различных эндосомальных компартментах регулируется разными факторами, такими как содержание и активность v-АТФазы, а также ионной проводимостью и свойствами других транспортных АТФаз, присутствующих в мембранах различных эндосом. Важно отметить, что pH в каждом компартменте зависит от выполняемой им функции.
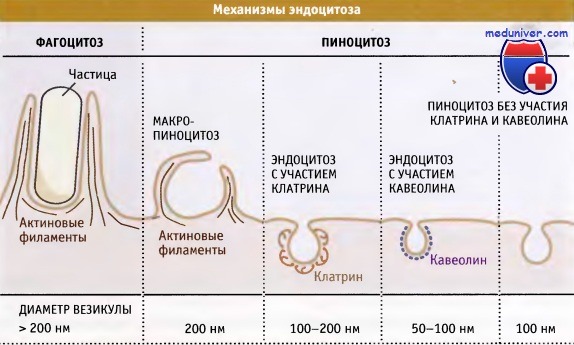 Существует несколько механизмов эндоцитоза.
Существует несколько механизмов эндоцитоза.
Исторически сложилось так, что эндоцитоз стали подразделять на фагоцитоз («поедание клеткой») и пиноцитоз («впитывание клеткой»), что отражает объем попадающего в клетку материала. Специализированный фагоцит, например макрофаг, может заключать материал в везикулу, диаметром до 10 мкм. Сходство с процессом поглощения пищи подчеркивается также сильно кислой внутренней средой фагосом, оптимальной для активности ферментов, расщепляющих поглощенные белки, липиды и углеводы.
Почти все клетки обладают способностью к фагоцитозу, однако этот процесс наиболее характерен для специализированных клеток иммунной системы, таких как макрофаги и дендритные клетки, поглощающих патогенные микроорганизмы и помогающих формировать ся защитной реакции у организма хозяина. Макрофаги также участвуют в очистке организма от стареющих или апоптотических клеток путем фагоцитоза, происходящего даже в отсутствие инфекции. Поскольку настоящие фагоциты обладают специфическими рецепторами, запускающими фагоцитоз, они более эффективны в захвате частиц по сравнению с другими клетками. Например, макрофаги и некоторые другие фагоцитирующие клетки экспрессируют рецепторы для молекул антител.
Пиноцитоз представляет собой общий термин, включающий несколько механизмов захвата. Обычно при этом образуется небольшая эндоцитозная везикула, достигающая 0,1-0,3 мкм в диаметре. Наиболее полно изученной разновидностью пиноцитоза является эндоцитоз, опосредуемый рецепторами. В ранних исследованиях по захвату липопротеинов низкой плотности (ЛНП) соответствующими рецепторами было получено много концептуальных сведений, способствующих нашему пониманию механизмов эндоцитоза. Большое количество рецепторов, расположенных на поверхности клеток, связывает лиганды, к числу которых относятся питательные вещества, ростовые факторы, гормоны, антитела или антигены.
Комплексы «рецептор-лиганд» поглощаются, накапливаясь в специфических областях плазматической мембраны, которые называются «окаймленные ямки». Окаймленные ямки отпочковываются, образуя везикулы, которые сливаются с ранними эндосомами. В кислой среде внутри эндосом связь некоторых лигандов с рецепторами разрывается, причем рецепторы направляются назад, на поверхность клетки, а лиганды — в поздние эндосомы и потом в лизосомы. Некоторые комплексы рецептора и лиганда в ранних эндосомах не диссоциируют, а вместо этого, так же как свободные лиганды, направляются в лизосомы.
Существует несколько вариантов основного пути эндоцитоза. Некоторые клетки содержат лизосомы, в которых происходит лишь частичная деградация поглощенного материала, что существенно для выработки иммунного ответа на захваченные патогены. Хорошим примером являются дендритные клетки, представляющие собой клетки лейкоцитарного происхождения, присутствующие в крови и во всех тканях организма.
Они инициируют почти все иммунные реакции, что обусловлено их уникальной способностью стимулировать В- и Т-лимфоциты, узнающие и уничтожающие патогены. Дендритные клетки захватывают циркулирующие антигены или патогенные микроорганизмы и доставляют их в специальный компартмент лизосом. Эти лизосомы не обладают способностью к полному перевариванию белков, что благоприятствует образованию коротких пептидов (10-15 остатков аминокислот), связывающихся с белками главного комплекса гистосовместимости класса II (МНС). Еще одна особенность этих лизосом позволяет комплексам пептидов и МНС класса II выходить из них за счет образования длинных трубочек, по которым содержимое транспортируется к плазматической мембране. Затем комплексы пептида и МНС класса II выходят на поверхность мембраны, где они стимулируют эффекторные клетки.
 Электронная микрофотография макрофага,
Электронная микрофотография макрофага,
поглощающего эритроцит.
Еще один вариант эндоцитоза предполагает, что поглощенный клеткой материал полностью минует лизосомный путь. Этот процесс носит название трансцитоз, он характерен для специализированных эпителиальных клеток, выстилающих полости тела, например внутреннюю поверхность кишечника.
Функция эпителиальных клеток состоит в регуляции транспорта материалов между организмом и окружающей средой. Эти клетки тесно примыкают друг к другу и образуют непрерывный слой. Они поляризованы, причем «апикальная» поверхность обращена в сторону люмена кишки, а «базальная» — в противоположную сторону, к крови. Обычно трансцитоз начинается с образования покрытой клатрином везикулы на апикальной или базолатеральной мембране. Эта везикула сливается с ранней эндосомой, что приводит к образованию трансцитозной везикулы, принадлежащей к типу рециклирующих везикул. Трансцитозная везикула переносится на противоположную поверхность клетки и сливается с мембраной.
При всасывании в кишечнике питательных веществ некоторые транспортные везикулы, которые образуются при эндоцитозе на апикальной поверхности, продвигаются прямо на базальную сторону и таким образом доставляют интернализированные компоненты без риска их разрушения в лизосомах. Еще одним примером трансцитоза является перенос факторов гуморального иммунитета от матери к новорожденному. Иммуноглобулины, присутствующие в материнском молоке, в кишечнике ребенка захватываются апикальными рецепторами, которые транспортируют их на другую сторону эпителия и доставляют в плазму крови.
Наряду с переносом через окаймленные ямки, на плазматической мембране образуются другие типы эндоцитозных везикул. Кавеолы представляют собой небольшие инвагинации в поверхности мембран, покрытые кавеолином. По-видимому, в кавеолах накапливаются некоторые рецепторы и мембранные липиды, которые не переносятся через окаймленные ямки. Эти рецепторы и липиды попадают в клетку, когда кавеола отпочковывается, образуя везикулу. Плазматическая мембрана также образует более крупные гетерогенные структуры, которые называются макропиносомы.
Обычно они образуются в ответ на действие факторов роста и представляют собой большие вакуоли, похожие по размеру на фагосомы. В них могут содержаться крупные капли внеклеточной жидкости. Материал, попавший в клетку с помощью кавеол и макропиносом, достигает тех же эндосом и лизосом, так же как и материал, поглощенный через окаймленные ямки. В некоторых случаях содержимое кавеол может храниться в специальной эндосоме, которая называется кавеосома. Наконец, некоторые метаболиты попадают в клетку вместе с мелкими пиноцитозными везикулами, которые образуются в отсутствие известных типов оболочек (окаймления).
Интересно, что некоторые вирусы животных используют кислые значения pH, характерные для эндоцитоза, для проникновения в цитоплазму клетки, где они проходят цикл репликации. Так, вирусы везикулярного стоматита и леса Семлики проникают в клетку через кислое окружение люмена эндосом; при этом активируются гликопротеины «шипов» на поверхности вирусной частицы и запускается процесс слияния мембран вируса и эндосомы. При этом происходит попадание в цитоплазму клетки вирусного генома и начинается распространение инфекции.
В клетках почкующихся дрожжей S. cerevisiae происходит пиноцитоз, но не фагоцитоз. Это не вызывает удивления, поскольку дрожжевая клетка обладает толстой клеточной стенкой. У этих клеток плохо различимы органеллы, принимающие участие в эндоцитозе, поскольку их очень мало. Однако вакуоли, которые представляют собой аналоги лизосом, хорошо видны в электронном микроскопе. До недавних пор генетические исследования эндоцитоза на дрожжах отставали от исследований экзоцитоза, и лишь в последнее время они стали особенно важны для понимания роли убиквитинирования белков в процессе их деградации.
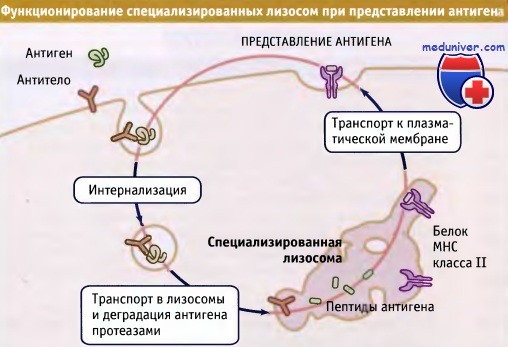 В клетках иммунной системы специализированные лизосомы разрушают захваченные белковые антигены,
В клетках иммунной системы специализированные лизосомы разрушают захваченные белковые антигены,
образуя пептиды, которые связываются с молекулами МНС класса II.
Комплексы пептид-МНС класса II транспортируются в плазматическую мембрану для представления антигена эффекторным клеткам. ![]() В поляризованной клетке процесс трансцитоза протекает таким образом,
В поляризованной клетке процесс трансцитоза протекает таким образом,
что материал, транспортируемый по механизму эндоцитоза из одного мембранного домена,
проходит через клетку и путем экзоцитоза выходит через другой мембранный домен.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в содержание раздела «генетика» на нашем сайте
Источник
Библиотека
С.Т. Метельский доктор биологических наук, главный научный сотрудник ГУ НИИ Общей патологии и патофизиологии РАМН; контактная информация для переписки — S.T.Metelsky@gmail.com ; Москва, 125315, Балтийская 8.
Цель лекции. Рассмотреть физиологические механизмы всасывания в желудочнокишечном тракте (ЖКТ).
Основные положения. В литературе данные вопросы освещаются с трех сторон: 1) топография всасывания веществ в различных отделах ЖКТ – желудок, двенадцатиперстная кишка, тощая, подвздошная и толстая кишка; 2) основные функции энтероцитов; 3) основные механизмы всасывания в кишечнике. Рассмотрено 7 основных механизмов всасывания веществ в кишечнике.
Заключение. Из всего ЖКТ тощая и подвздошная кишка характеризуются самым широким спектром всасывания различных соединений. Понимание физиологических механизмов всасывания в тонкой кишке имеет большое значение в практической гастроэнтерологии.
Ключевые слова:
Всасывание, ионы, натрий, нутриенты, желудочнокишечный тракт, простая диффузия, облегченная диффузия, осмос, фильтрация, околоклеточный транспорт, активный транспорт, сопряженный транспорт, вторично-энергизованный транспорт, эндоцитоз, трансцитоз, Р-гликопротеин.
 Основные механизмы всасывания
Основные механизмы всасывания
 Основные механизмы всасывания
Основные механизмы всасыванияВсасывание в различных отделах желудочно-кишечного тракта
В желудке всасывается 20% потребленного алкоголя, а также короткоцепочечные жирные кислоты. В двенадцатиперстной кишке – витамины A и B1, железо, кальций, глицерин, жирные кислоты, моноглицериды, аминокислоты, моно- и дисахариды. В тощей кишке – глюкоза, галактоза, аминокислоты и дипептиды, глицерин и жирные кислоты, моно- и диглицериды, медь, цинк, калий, кальций, магний, фосфор, йод, железо, жирорастворимые витамины D, E и K, значительная часть комплекса витаминов В, витамин С и остатки алкоголя. В подвздошной кишке – дисахариды, натрий, калий, хлорид, кальций, магний, фосфор, йод, витамины C, D, E, K, B1, B2, B6, B12 и большая часть воды. В толстой кишке – натрий, калий, вода, газы, некоторые жирные кислоты, образовавшиеся при метаболизме растительных волокон и непереваренного крахмала, витамины, синтезированные бактериями, – биотин (витамин Н) и витамин К.
Основные функции энтероцитов
Основные механизмы всасывания соединений в кишечнике
На рис. 2 представлены основные механизмы всасывания веществ. Рассмотрим указанные механизмы более подробно.
Пресистемный метаболизм, или метаболизм (эффект) первого прохождения кишечной стенки. Явление, при котором концентрация вещества перед попаданием в кровеносное русло резко снижается. При этом если введенное вещество является субстратом P-гликопротеина (см. ниже), его молекулы могут неоднократно поступать в энтероциты и выводиться из него, в результате чего вероятность метаболизма данного соединения в энтероцитах возрастает.
P-гликопротеин в большом количестве экспрессирован в нормальных клетках, выстилающих кишечник, проксимальные канальцы почек, капилляры гематоэнцефалического барьера, и в клетках печени. Транспортеры типа P-гликопротеина являются членами надсемейства самого большого и наиболее древнего семейства транспортеров, представленного в организмах от прокариотов до человека. Это трансмембранные белки, функцией которых является транспорт широкого спектра
| |
веществ через вне- и внутриклеточные мембраны, включая продукты метаболизма, липиды и лекарственные вещества. Такие белки классифицируются как АТФ-связывающие кассетные транспортеры (АВС-транспортеры) на основании их последовательности и устройства АТФ-связывающего домена. АВС-транспортеры влияют на невосприимчивость к лекарственным средствам опухолей, кистозного фиброза, устойчивость бактерий ко многим лекарственным препаратам и некоторые другие явления.
Пассивный перенос веществ через эпителиальный пласт. Пассивный транспорт веществ через монослой энтероцитов протекает без затрат свободной энергии и может осуществляться или трансклеточным, или околоклеточным путем. К этому виду транспорта относятся простая диффузия (рис. 3), осмос (рис. 4) и фильтрация (рис. 5). Движущей силой диффузии молекул растворенного вещества является его концентрационный градиент.
Зависимость скорости диффузии вещества от его концентрации линейна.Диффузия – это наименее специфичный и самый, по-видимому, медленный процесс транспорта. При осмосе, представляющем собой разновидность диффузионного переноса, происходит перемещение в соответствии с концентрационным градиентом свободных (не связанных с веществом) молекул растворителя (воды).
| |
Процесс фильтрации заключается в переносе раствора через пористую К пассивному переносу веществ через мембраны относится также облегченная диффузия – перенос веществ с помощью транспортеров, т. е. специальных каналов или пор (рис. 6). Облеченная диффузия обладает специфичностью к субстрату. Зависимость скорости процесса при достаточно высоких концентрациях переносимого вещества выходит на насыщение, поскольку перенос очередной молекулы тормозится ожиданием, когда транспортер освободится от переноса предыдущей.
Околоклеточный транспорт – это транспорт соединений между клетками через область плотных контактов (рис. 7), он не требует затрат энергии. Структура и проницаемость плотных контактов тонкой кишки в настоящее время активно исследуются и дискутируются. Например, известно, что за селективность плотных контактов для натрия отвечает клаудин-2.
Другая возможность состоит в том, что межклеточный перенос осуществляется благодаря некоторым дефектам в эпителиальном пласте. Такое движение может происходить по межклеточным областям в тех местах, где происходит слущивание отдельных клеток.  Такой путь может оказаться воротами для проникновения чужеродных макромолекул прямо в кровь или в тканевые жидкости.
Такой путь может оказаться воротами для проникновения чужеродных макромолекул прямо в кровь или в тканевые жидкости.
Эндоцитоз, экзоцитоз, рецепторопосредованный транспорт (рис. 8) и трансцитоз. Эндоцитоз – это везикулярный захват жидкости, макромолекул или небольших частиц в клетку. Существуют три механизма эндоцитоза: пиноцитоз (от греческих слов «пить» и «клетка»), фагоцитоз (от греческих слов «поедать» и «клетка») и рецепторопосредованный эндоцитоз или клатрин-зависимый эндоцитоз. Нарушения указанного механизма приводят к развитию определенных заболеваний. Многие кишечные токсины, в частности холерный, попадают в энтероциты именно по этому механизму.
При пиноцитозе гибкая плазматическая мембрана образует впячивание (инвагинация) в виде ямки. Такая ямка заполняется жидкостью из внешней среды. Затем она отшнуровывается от мембраны и в виде везикулы продвигается в цитоплазму, где ее мембранные стенки перевариваются, а содержимое высвобождается. Благодаря такому процессу клетки могут поглощать как крупные молекулы, так и различные ионы, не способные проникнуть через мембрану самостоятельно. Пиноцитоз часто наблюдается в клетках, функция которых связана со всасыванием. Это чрезвычайно интенсивный процесс: в некоторых клетках 100% плазматической мембраны поглощается и восстанавливается всего за час.
При фагоцитозе (явление открыто русским ученым И.И. Мечниковым в 1882 г.) выросты цитоплазмы захватывают капельки жидкости, содержащие какие-либо плотные (живые или неживые) частицы (до 0,5 мкм), и втягивают их в толщу цитоплазмы, где гидролизующие ферменты переваривают поглощенный материал, разрушая его до таких фрагментов, которые могут быть усвоены клеткой. Фагоцитоз осуществляется с помощью клатрин-независимого актин-зависимого механизма; это – основной механизм защиты организма хозяина от микроорганизмов. Фагоцитоз поврежденных или постаревших клеток необходим для обновления тканей и заживления ран.
При рецепторопосредованном эндоцитозе (см. рис. 8) для переноса молекул используются специфические поверхностные рецепторы. Этот механизм обладает следующими свойствами – специфичность, способность к концентрированию лиганда на поверхности клетки, рефрактерность. Если специфический рецептор после связывания лиганда и его поглощения не возвращается на мембрану, клетка становится рефрактерной к данному лиганду.  С помощью эндоцитозного везикулярного механизма всасываются как высокомолекулярные соединения типа витамина В12, ферритина и гемоглобина, так и низкомолекулярные – кальций, железо и др. Роль эндоцитоза особенно велика в раннем постнатальном периоде. У взрослого человека пиноцитозный тип всасывания существенного значения в обеспечении организма питательными веществами, по-видимому, не имеет.
С помощью эндоцитозного везикулярного механизма всасываются как высокомолекулярные соединения типа витамина В12, ферритина и гемоглобина, так и низкомолекулярные – кальций, железо и др. Роль эндоцитоза особенно велика в раннем постнатальном периоде. У взрослого человека пиноцитозный тип всасывания существенного значения в обеспечении организма питательными веществами, по-видимому, не имеет.
Трансцитоз – это механизм, посредством которого молекулы, пришедшие в клетку извне, могут доставляться к различным компартментам внутри клетки или даже перемещаться от одного слоя клеток к другому. Одним из хорошо изученных примеров трансцитоза является проникновение некоторых материнских иммуноглобулинов через клетки кишечного эпителия новорожденного. Материнские антитела с молоком попадают в организм ребенка. Антитела, связанные с соответствующими рецепторами, сортируются в ранние эндосомы клеток пищеварительного тракта, затем с помощью других пузырьков проходят сквозь эпителиальную клетку и сливаются с плазматической мембраной на базолатеральной поверхности. Здесь лиганды освобождаются от рецепторов. Затем иммуноглобулины собираются в лимфатические сосуды и попадают в кровоток новорожденного.
Рассмотрение механизмов всасывания с точки зрения отдельных групп веществ и соединений будут представлены в одном из следующих номеров журнала.
Работа поддержана грантом РФФИ 09-04-01698
Список литературы:
1. Метельский С.Т. Транспортные процессы и мембранное пищеварение в слизистой оболочке тонкой кишки. Электрофизиологическая модель. – М.: Анахарсис, 2007. – 272 с.
2. Общий курс физиологии человека и животных. – Кн. 2. Физиология висцеральных систем / Под ред. А.Д. Ноздрачева. – М.: Высшая школа, 1991. – С. 356–404.
3. Membrane digestion. New facts and concepts / Ed. A.M. Ugolev. – M.: MIR Publishers, 1989. – 288 p.
4. Tansey T., Christie D.A., Tansey E.M. Intestinal absorption. – London: Wellcome Trust, 2000. – 81 p
статья взята с сайта Русского журнала Гастроэнтерологии, Гепатологии, Колопроктологии
Источник



