
- Способы транспорта веществ через мембрану клетки
- Биология. 11 класс
- § 11. Поверхностный аппарат клетки
- Транспорт веществ через мембрану клетки. Виды транспорта веществ и их механизмы
- Транспорт питательных веществ
- Схема процесса пассивной диффузии
- Пассивная диффузия
- Схема процесса облегченной диффузии
- Облегченная диффузия
- Схема активного транспорта
- Активный транспорт
Способы транспорта веществ через мембрану клетки
• Существует два основных типа мембранных транспортных белков: каналы и переносчики
• Ионные каналы катализируют быстрый и избирательный транспорт ионов по электрохимическому градиенту
• Транспортеры и насосы представляют собой белки-переносчики, которым необходима энергия для транспорта веществ против электрохимического градиента
• В каждой клетке несколько различных мембранных транспортных белков действуют как единое целое
Некоторые из транспортных белков находятся в плазматической мембране, в то время как другие входят в состав мембраны клеточных органелл. Для поддержания определенного состава внутриклеточной среды и клеточных компартментов необходимо, чтобы транспортные белки обладали избирательностью по отношению к определенным метаболитам. В зависимости от характера процесса транспорта мембранные транспортные белки подразделяются на две группы: каналы и переносчики.
Канальные белки содержат поры, через которые при открытом канале с высокой скоростью проходят метаболиты. Белок-переносчик связывает метаболит, находящийся по одну сторону мембраны, после чего конформация его меняется, и метаболит высвобождается с другой стороны мембраны.
Существует несколько типов белков, образующих каналы в мембранах. Порины, присутствующие в клетках некоторых прокариот, в митохондриях и в нексусах, где соединяется цитоплазма соседних клеток, обеспечивают прохождение метаболитов в соответствии с размером их молекул.
К более селективным каналам относятся комплексы ядерных пор и каналы, обеспечивающие перемещение белков через мембрану эндоплазматического ретикулума. В статьях на сайте будут рассмотрены ионные каналы и аквапорины, через которые осуществляется селективный трансмембранный перенос ионов и молекул воды соответственно. К настоящему времени описано более 100 различных типов канальных белков. Все они обладают следующими характеристиками:
• селективностью по отношению к переносимому метаболиту,
• высокой скоростью переноса метаболита и
• воротным механизмом, регулирующим транспорт метаболитов.
Участок канального белка, через который осуществляется трансмембранный транспорт, называется порой канала. Канал может находиться в нескольких различных конфигурациях. Некоторые каналы состоят из одного белка, трансмембранные сегменты которого образуют пору. Другие существуют в виде олигомеров, состоящих из идентичных или разных субъединиц, которые вместе образуют пору.
Наконец существуют каналы, состоящие из двух и более субъединиц, каждая из которых сама образует пору. Часто комплексы олигомеров каналов регулируются или позиционируются на определенных мембранах с участием дополнительных субъединиц.
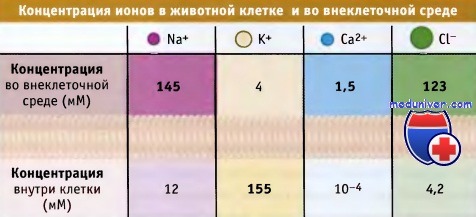
Большинство канальных белков обладают высокой селективностью в отношении определенных метаболитов, например ионов натрия (Na+), калия (К+), кальция (Са2+), хлорида (Сl-) или воды. Другие канальные белки являются неселективными катионными или анионными каналами. Как будет показано далее, при рассмотрении свойств специфических канальных белков, их поры характеризуются определенными структурными особенностями, т. е. являются селективным фильтром, который позволяет им проявлять избирательность по отношению к различным метаболитам.
![]() Каналы и переносчики представляют собой два основных типа белков мембранного транспорта.
Каналы и переносчики представляют собой два основных типа белков мембранного транспорта.
Растворенные метаболиты с максимальной скоростью диффундируют через пору каналов,
а переносчики связывают метаболит с одной стороны мембраны, затем претерпевают конформационные изменения и высвобождают его с другой стороны.
Этот процесс протекает значительно медленнее.
Направление потока ионов растворенных веществ через канал определяется электрохимическим градиентом. Иными словами, метаболиты движутся через канал в энергетически-выгодном направлении, т. е. по электрохимическому градиенту. Например, для покоящейся клетки, схематически изображенной на рисунке ниже, существует направленный в клетку ионный поток через Na+-, Са2+-, Cl—каналы, и противоположный, проходящий через К+-каналы.
Так как других источников энергии, кроме электрохимического градиента, в данном случае не используется, то такой тип транспорта называется пассивным. Канальные белки обеспечивают высокую скорость транспорта. Так, для ионных каналов скорость составляет 108 в 1 с, что близко к максимальной скорости диффузии ионов в водной среде.
Транспортные мембранные белки контролируют различные клеточные функции при участии воротного механизма. В основе этого механизма лежит индукция конформационных изменений в белке в ответ на воздействие определенного стимула. Эти изменения приводят к открытию или закрытию канала. Например, открытие и закрытие ионных каналов могут регулироваться лигандами, величиной электрического потенциала, механическими или температурными воздействиями. Активация каналов может происходить очень быстро, что делает их идеальным инструментом обработки поступающих в клетку сигналов.
Например, передача нервного импульса зависит от электрического заряда, который создается ионным каналом в мембране. Каналы также играют важную роль в регуляции объема клетки и внутриклеточного pH, в транспорте солей и воды через эпителиальные клетки, в закислении внутриклеточных органелл и во внеклеточной передаче сигналов.
 Транспортеры и насосы представляют собой два основных типа белков-переносчиков.
Транспортеры и насосы представляют собой два основных типа белков-переносчиков.
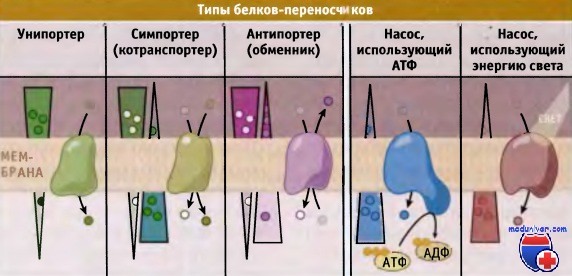
Транспортеры делятся на три типа, каждый из которых осуществляет один из типов переноса: унипорт, симпорт, или антипорт.
Показано направление электрохимического градиента через мембрану.
В зависимости от типа белка-переносчика транспорт может происходить или в направлении градиента (от высокой концентрации к низкой), или против него.
При транспорте субстратов против градиента концентрации, белки-переносчики используют свободную энергию электрохимического градиента, АТФ или других источников. Поскольку при этом происходит потребление энергии, процесс носит название активный транспорт. Белки-переносчики подразделяются на две группы, транспортеры и насосы.
Для транспортировки субстратов через мембрану транспортеры используют энергию электрохимического градиента. Они подразделяются на унипортеры, симпортеры (или котранс-портеры) и антипортеры (обменники). Для энергозависимого транспорта метаболитов мембранные насосы используют энергию, например высвобождающуюся при гидролизе АТФ. По сравнению с канальными белками, переносчики характеризуются более медленной скоростью транспорта, порядка 1000 молекул в 1 с.
Различают два типа активного транспорта, первичный и вторичный. Белки-переносчики, осуществляющие первичный активный транспорт, в качестве источника энергии используют АТФ и транспортируют метаболиты против электрохимического градиента. Таким образом, они помогают поддерживать градиент концентрации метаболитов по обеим сторонам мембран клетки. Са2+-АТФаза и Na+/К+-АТФаза являются характерными примерами транспортных насосов, которые управляют процессом первичного активного транспорта.
Белки-переносчики, которые участвуют во вторичном активном транспорте, непосредственно не используют энергию АТФ. Вместо этого они используют свободную энергию электрохимического градиента, который возникает за счет первичного активного транспорта. Вторичный активный транспорт происходит с участием симпортеров и антипортеров. Механизмы транспорта метаболитов представлены на рисунке ниже.
В клетке все типы мембранных транспортных белков-каналы, транспортеры и насосы-функционируют совместно. Таким образом, работа какого-либо одного типа транспортного белка зависит от работы других. В данной главе мы рассмотрим несколько примеров такого взаимодействия транспортных белков. Например, ионный градиент по обеим сторонам мембраны поддерживается за счет сложных взаимодействий между различными типами транспортных белков. Для правильного функционирования эпителиальных клеток, например почек, слизистой кишечника и легкого, необходимо протекание транспортных процессов с участием различных ионов и метаболитов. Мы также рассмотрим роль нарушений процессов транспорта в развитии различных заболеваний.
![]() Транспорт растворенных метаболитов через мембрану может быть пассивным или активным.
Транспорт растворенных метаболитов через мембрану может быть пассивным или активным.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
Биология. 11 класс
§ 11. Поверхностный аппарат клетки
Обязательным компонентом поверхностного аппарата любой клетки является цитоплазматическая мембрана (плазмалемма). Она отделяет и защищает внутреннее содержимое клетки от внешней среды. Важнейшим свойством цитоплазматической мембраны является избирательная проницаемость. Различные вещества проходят через плазмалемму с разной скоростью, а для некоторых она практически непроницаема. Таким образом, цитоплазматическая мембрана обеспечивает обмен веществ между клеткой и внеклеточной средой и постоянство химического состава клетки.
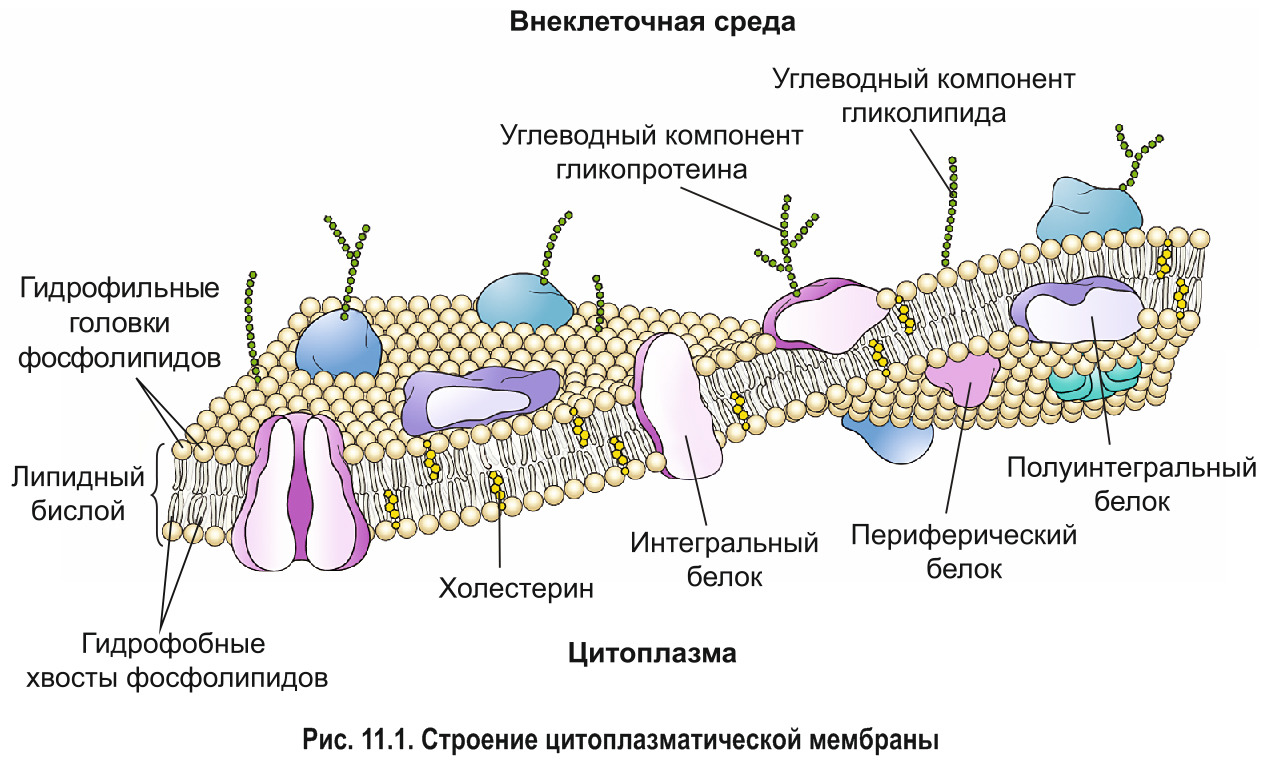
Химический состав и строение плазмалеммы. Цитоплазматическая мембрана состоит преимущественно из липидов и белков. В состав плазмалеммы также входят углеводы, которые являются компонентами сложных липидов и белков — гликолипидов и гликопротеинов. Толщина цитоплазматической мембраны составляет около 7,5 нм.
Около половины массы плазмалеммы составляют липиды, представленные в основном фосфолипидами. Они формируют основу строения мембраны — липидный бислой (двойной слой), в котором гидрофобные хвосты молекул погружены внутрь, а гидрофильные головки располагаются снаружи (см. рис. 6.3). Кроме фосфолипидов, в состав двойного слоя входят и другие липиды например стероиды.
*С липидным бислоем связаны белки. Молекулы интегральных белков пронизывают двойной слой липидов насквозь. Полуинтегральными называют белки, погруженные лишь в один из двух липидных слоев (внешний либо внутренний). Периферические белки примыкают к поверхности бислоя с наружной или внутренней стороны (рис. 11.1).*
Мембранные липиды и белки связаны между собой не ковалентными связями, а за счет гидрофобных и электростатических взаимодействий. В связи с этим молекулы данных веществ могут двигаться вдоль плоскости мембраны. Таким образом, плазмалемма является динамичной структурой — молекулы белков перемещаются в подвижном, текучем липидном бислое, как в жидкости. *Данная модель организации мембраны получила название жидкостно-мозаичной. Она была предложена американскими биологами Дж. Сингером и Г. Николсоном в 1972 г.* Свойство текучести обусловливает пластичность плазмалеммы, благодаря чему она способна, например, быстро восстанавливать свою целостность после незначительных повреждений.
*Движение в плоскости плазмалеммы происходит достаточно легко. Однако переход белков с одной стороны мембраны на другую (так называемый флип-флоп переход) практически невозможен. Подобное перемещение липидов происходит, но очень редко, при участии специальных белков флиппаз. Поэтому состав наружного и внутреннего слоев плазмалеммы различается.*
*Текучесть мембраны зависит от ее липидного состава и температуры. С увеличением содержания остатков ненасыщенных жирных кислот или температуры подвижность молекул возрастает. Стероиды , например холестерин в организме животных, регулируют текучесть плазмалеммы и придают ей жесткость. Стероиды располагаются в свободном пространстве между гидрофобными хвостами фосфолипидов, ограничивая подвижность их молекул друг относительно друга (см. рис. 11.1). Таким образом, стероиды обеспечивают стабильность мембраны и поддержание ее структуры. Кроме того, они регулируют проницаемость плазмалеммы.*
В клетках эукариот, кроме плазмалеммы, имеются внутренние мембраны, ограничивающие ядро и мембранные органоиды. Для всех биологических мембран характерен общий план строения — их основу составляет липидный бислой, с которым связаны молекулы белков.
Источник
Транспорт веществ через мембрану клетки. Виды транспорта веществ и их механизмы

Транспорт веществ через мембрану.
Если вещество движется через мембрану из области с высокой концентрацией в сторону низкой концентрации (т. е. по градиенту концентрации этого вещества) без затраты клеткой энергии, то такой транспорт называется пассивным, или диффузией. Различают два типа диффузии: простую и облегченную.
Характерна для небольших нейтральных молекул (H2O, CO2, O2), а также гидрофобных низкомолекулярных органических веществ. Эти молекулы могут проходить без какого-либо взаимодействия с мембранными белками через поры или каналы мембраны до тех пор, пока будет сохраняться градиент концентрации.
Характерна для гидрофильных молекул, которые переносятся через мембрану также по градиенту концентрации, но с помощью специальных мембранных белков — переносчиков. Для облегченной диффузии, в отличие от простой, характерна высокая избирательность, так как белок переносчик имеет центр связывания комплементарный транспортируемому веществу, и перенос сопровождается конформационными изменениями белка. Один из возможных механизмов облегченной диффузии может быть следующим: транспортный белок (транслоказа) связывает вещество, затем сближается с противоположной стороной мембраны, освобождает это вещество, принимает исходную конформацию и вновь готов выполнять транспортную функцию. Мало известно о том, как осуществляется передвижение самого белка. Другой возможный механизм переноса предполагает участие нескольких белков-переносчиков. В этом случае первоначально связанное соединение само переходит от одного белка к другому, последовательно связываясь то с одним, то с другим белком, пока не окажется на противоположной стороне мембраны.
Имеет место в том случае, когда перенос осуществляется против градиента концентрации. Такой перенос требует затраты энергии клеткой. Активный транспорт служит для накопления веществ внутри клетки. Источником энергии часто является АТФ. Для активного транспорта кроме источника энергии необходимо участие мембранных белков. Одна из активных транспортных систем в клетке животных отвечает за перенос ионов Na+ и K+ через клеточную мембрану. Эта система называется Na+ — K+ — насос. Она отвечает за поддержание состава внутриклеточной среды, в которой концентрация К+ выше, чем Na+.
Источник
Транспорт питательных веществ

Транспорт питательных веществ – это процесс прохождения веществ из окружающей среды через цитоплазматическую мембрану (ЦПМ) в бактериальную клетку [1] .
Выделяют несколько типов транспортных систем, которые позволяют различным веществам преодолевать преграду цитоплазматической мембраны (ЦПМ) и попадать внутрь клетки микроорганизма. Это пассивная диффузия, облегченная диффузия, активный транспорт. Отмечается, что только активный транспорт способствует аккумуляции (накоплению) веществ внутри клетки [1] .

Схема процесса пассивной диффузии

Пассивная диффузия
Пассивная или простая диффузия – неспецифический процесс. Он происходит за счет разницы концентраций. Передвижение молекул осуществляется из более концентрированного раствора в менее концентрированный (по градиенту их концентрации).Этот процесс не связан с затратой энергии. Таким путем в клетку попадают низкомолекулярные вещества: кислород, липофильные соединения (спирты, жирные кислоты), вода, яды и другие, чужеродные для клетки вещества. Таким же образом происходит удаление продуктов обмена. Скорость перемещения веществ путем пассивной диффузии невелика и зависит от размеров транспортирующихся молекул [4] [3] .

Схема процесса облегченной диффузии

Облегченная диффузия
Облегченная диффузия – перенос веществ через цитоплазматическую мембрану по градиенту их концентрации с участием пермеаз (транслоказ) – специфических мембранных белков, способствующих прохождению веществ через цитоплазматическую мембрану [3] [1] .
Пермеаза фиксирует на себе молекулу переносимого вещества, вместе с ней преодолевает цитоплазматичекую мембрану. После этого комплекс «вещество-пермеаза» диссоциирует. Освободившаяся пермеаза, диффундирует к наружной поверхности, присоединяет новую молекулу вещества и транспортирует ее внутрь клетки [1] .
Облегченная диффузия не требует расхода энергии, если наружная концентрация вещества выше внутренней, поскольку в таком случае вещество перемещается «вниз» по химическому градиенту. Скорость процесса зависит от концентрации вещества в наружном растворе. Предполагается, что выход продуктов обмена веществ из микробной клетки может также происходит по методу облегченной диффузии с помощью переносчиков [1] .
Параллельно отмечается, что облегченная диффузия более характерна для эукариотических организмов [4] .

Схема активного транспорта

Активный транспорт
Активный транспорт является основным механизмом избирательного переноса вещества через цитоплазматическую мембрану в клетку против градиента концентрации. Этот процесс протекает при участии локализованных в цитоплазматической мембране переносчиков – пермеаз. Это вещества белковой природы, высокочувствительные к субстрату [3] .
Активным транспортом в цитоплазму бактериальной клетку поступает подавляющее большинство разнообразных веществ (ионы, углеводы, аминокислоты, липиды) [4] .
Для активного транспорта необходимы затраты энергии. Ее получают в виде АТФ, либо за счет протондвижущей силы энергизованной мембраны [3] .
У многих микробов, чаще у грамотрицательных бактерий, в активном транспорте принимают участие связующие белки. Эти вещества не входят в структуру мембраны, не идентичны пермеазам. Они локализованы в периплазматическом пространстве. Связующие белки не имеют каталитической активности, но обладают высоким сродством к определенным питательным веществам, аминокислотам, углеводам, неорганическим ионам. Выделено и изучено более 100 различных связующих белков [3] .
Активный транспорт осуществляется двумя путями:
- Без химической модификации переносимого вещества [4] .
- С химической модификацией переносимого вещества [4] .
В первом случае молекула питательного вещества образует комплекс с белком периплазматического пространства. Белок взаимодействует со специфической пермеазой цитоплазматической мембраны. После энергозависимого проникновения через цитоплазматическую мембрану комплекс «субстрат – белок периплазмы – пермеаза» диссоциирует и молекула субстрата освобождается [4] .
Во втором случае наблюдается следующие последовательные процессы:
- Фосфорилирование мембранного фермента-2 со стороны цитоплазмы фосфоенолпируватом [4] .
- Молекула субстрата связывается на поверхности цитоплазматической мембраны фосфорилированным ферментом-2 [4] .
- Энергозависимый транспорт молекулы субстрата через мембрану в цитоплазму[4] .
- Перенос фосфатной группы на молекулу сбстрата [4] .
- Диссоциация в цитоплазме комплекса «субстрат-фермент» [4] .
Отмечается, что молекулы субстрата аккумулируются в цитоплазме клеток и теряют способность выйти из них именно за счет фосфорилирования [4] .
Отдельные авторы второй путь активного транспорта (с химической модификацией переносимого вещества) выделяют в отдельный (четвертый) способ транспорта питательных веществ – транслокацию (перенос) групп (радикалов) [1] [3] .
Источник



