
- Транспорт углекислого газа. Углекислый газ транспортируется от тканей к легким
- Транспорт углекислого газа
- Газообмен происходит по градиенту концентраций
- Обмен кислорода и углекислого газа в тканях
- Обмен кислорода и углекислого газа в легких
- 234. Транспорт углекислоты кровью, значение карбоангидразы, взаимосвязь транспорта о2и со2.
- 235. Иннервация дыхательных мышц.
- 236. Дыхательный центр. Современные представления о структуре и локализации. Автоматия дыхательного центра.
Транспорт углекислого газа. Углекислый газ транспортируется от тканей к легким
Углекислый газ транспортируется от тканей к легким. Осуществляется несколькими способами:
1. С помощью соединения углекислого газа с гемоглобином – карбгемоглобина.
Карбгемоглобин – это также нестойкое легко распадающееся соединение (как и оксигемоглобин).
Образуется карбгемоглобин в капиллярах большого круга кровообращения, когда кровь принимает от тканей углекислый газ. При этом кровь становится венозной.
Гемоглобин соединяется с 4 молекулами углекислого газа с помощью 4 атомов железа, содержащихся в геме одной молекулы гемоглобина.
А распадается карбгемоглобин в легких – когда кровь легочных капилляров отдает в альвеолярный воздух углекислый газ.
2. В виде угольной кислоты:
3. В виде солей карбонатов: NаНСО3, КНСО3, Са (НСО3)2 и др.
В виде раствора в крови.
Внутреннее (тканевое) дыхание (4 фаза дыхания)
Внутреннее (тканевое) дыхание включает:
– газообмен между кровью капилляров большого круга кровообращения и тканями;
— клеточное дыхание – потребление кислорода клетками и выделение ими углекислого газа.
Газообмен в тканях, как и в легких, происходит вследствие разности парциального давления (напряжения) кислорода и углекислого газа в крови капилляров и в тканях.
Ткани поглощают кислород и выделяют углекислый газ.
К тканям притекает артериальная кровь, насыщенная кислородом.
В этой крови парциальное давление кислорода больше, чем в тканях, поэтому кислород движется из области большего давления в область меньшего давления – из крови в ткани, и кровь отдает кислород.
В тканях при этом в результате обмена веществ уже накопился углекислый газ, поэтому парциальное давление углекислого газа в тканях больше, чем в крови, поэтому углекислый газ движется из области большего давления в область меньшего давления – из тканей в кровь, и ткани отдают углекислый газ, а кровь становится венозной.
Потребности в кислороде у различных органов различны.
Потребность в кислороде особенно высока у миокарда сердца, коры головного мозга, печени, почек.
При физической нагрузке потребление кислорода тканями увеличивается, например, у миокарда в 3-4 раза, в скелетных мышцах в 20 раз.
При нарушении газообмена в легких или при нарушении транспорта кислорода к тканям и в тканях возникает состояние гипоксии или аноксии.
При гипоксии нарушаются процессы жизнедеятельности, при аноксии наступает полная утрата функции, так, в нейронах необратимые изменения происходят уже через 10 мин. постоянной аноксии.
РЕГУЛЯЦИЯ ДЫХАНИЯ
Регуляция дыхания осуществляется нервным и гуморальным путем.
Источник
Транспорт углекислого газа




![]()
![]()
Транспорт углекислого газа, как и кислорода, осуществляется кровью в виде физического растворения и химической связи. Причем С02, как и 02, переносится и плазмой, и эритроцитами (И. М. Сеченов, 1859). Однако соотношение фракций С02, переносимых плазмой и эритроцитами, существенно отличается от таковых для 02. Ниже приведены усредненные показатели содержания С02 в крови.
Распределение С02 в плазме и эритроцитах. Большая часть С02 транспортируется плазмой крови, причем около 60 % всего С02 находится в виде бикарбоната натрия (МаНС03, 34 об%), т.е. в виде химической связи, 4,5 об% — в виде физически растворенного С02 и около 1,5% СО, находится в виде Н2С03. Всего в венозной крови содержится 58 об% С02. В эритроците С02 находится в форме химических соединений карбгемоглобина (ННЬС02, 5,5 об%) и бикарбоната калия (КНС03, 14 об%). Углекислый газ,
образуемый в организме, выделяется в основном через легкие (около 98%,) и только 0,5% — через почки, около 2% — через кожу в виде НС03-бикарбонатов.
Следует отметить, что некоторое увеличение содержания С02 в крови оказывает благоприятное влияние на организм: увеличивает кровоснабжение мозга и миокарда, стимулирует процессы биосинтеза и регенерацию поврежденных тканей. Увеличение содержания С02 в крови стимулирует также сосудодвигательный и дыхательный центры.
Образование соединений углекислого газа. В результате окислительных процессов и образования С02 его напряжение в клетках и, естественно, в межклеточных пространствах значительно больше (достигает 60- 80 мм рт.ст.), чем в поступающей к тканям артериальной крови (40 мм рт.ст.). Поэтому С02, согласно градиенту напряжения, из интерстиция переходит через стенку капилляров в кровь. Небольшая его часть остается в плазме в виде физического растворения. В плазме образуется также небольшое количество Н2С03 (Н20+С02 -> Н2С03), но этот процесс идет очень медленно, так как в плазме крови нет фермента карбоангидразы, катализирующего образование Н2С03
Карбоангидраза имеется в различных клетках организма, в том числе в лейкоцитах и тромбоцитах. С02 поступает и в эти клетки, где также образуются угольная кислота и ионы НС03
. Однако роль этих клеток в транспорте С02 невелика, так как они не содержат гемоглобина, их число значительно меньше, нежели эритроцитов, их размеры очень маленькие (тромбоциты имеют диаметр 2-3 мкм, эритроциты — 8 мкм).
Гемоглобин транспортирует не только 02, но и С02. При этом образуется так называемая карбаминовая связь: ННЬ + С02 = = ННЬС02 (НЬ-ЫН-СООН-карбгемоглобин, точнее — карбамино-гемоглобин).
Небольшое количество С02 (1 -2 %) переносится белками плазмы крови также в виде карбаминовых соединений.
Диссоциация соединений углекислого газа. В легких происходят обратные процессы — выделение из организма С02 (за сутки выделяется около 850 г С02). В первую очередь начинается выход в альвеолы физически растворенного С02 из плазмы крови, поскольку парциальное давление Рсо2 в альвеолах (40 мм рт.ст.) ниже, чем в венозной крови (46 мм рт.ст.). Это ведет к уменьшению напряжения Рсо2 в крови. Причем, присоединение кислорода к гемоглобину ведет к уменьшению сродства углекислого газа к гемоглобину и расщеплению карбгемоглобина (эффект Холдена). Общая схема процессов образования и диссоциации всех соединений кислорода
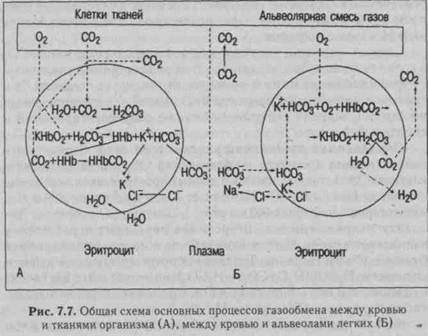
и углекислого газа, происходящих во время прохождения крови в капиллярах тканей и легких, представлена на рис. 7.7.
В процессе дыхания регулируется рН внутренней среды вследствие удаления С02 из организма, так как Н2С03 диссоциирует на Н20 и С02. При этом предотвращается закисление внутренней среды организма постоянно образующейся Н2С03.
Источник
Газообмен происходит по градиенту концентраций
Обмен кислорода и углекислого газа в тканях
В тканях диффундирующий в кровь из клеток СО2 большей частью (около 90%) по градиенту концентрации попадает в эритроциты. Движущей силой этого процесса является быстрая, постоянно идущая реакция превращения его в угольную кислоту при участии фермента карбоангидразы . Угольная кислота диссоциирует и подкисляет содержимое эритроцита, что улучшает отдачу оксигемоглобином кислорода (Эффект Бора).
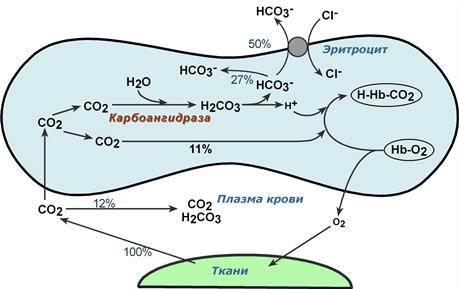
Реакции газообмена в капиллярах тканей
Одновременно с концевыми NH2-группами β-цепей гемоглобина связывается 10-12% двуокиси углерода (CO2) с образованием карбаминогемоглобина (H-HbCO2).
Остальные бикарбонаты выходят в плазму крови в обмен на ионы хлора (гипохлоремический сдвиг).
Таким образом, в составе карбаминогемоглобина транспортируется 10-12% CO2, в растворенном состоянии в плазме крови также до 12%, остальная часть находится в виде растворенного карбонат-иона (HCO3 – ) в плазме и эритроците, примерно поровну (соотношение 9 : 7).
Обмен кислорода и углекислого газа в легких
В легких в альвеолярном воздухе имеется высокая концентрация кислорода и относительно низкая концентрация углекислого газа. Поэтому происходит быстрая диффузия СО2 из плазмы через альвеолярные мембраны и его удаление с выдыхаемым воздухом,
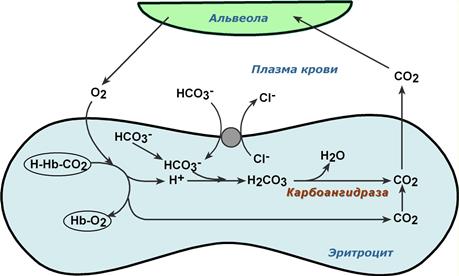
Реакции газообмена в легочных капиллярах
- уменьшение концентрации СО2 в плазме стимулирует его образование в карбоангидразной реакции внутри эритроцита и снижает здесь концентрацию иона HCO3 – ,
- одновременно высокая концентрация кислорода вытесняет СО2 из комплекса с гемоглобином с образованием оксигемоглобина – более сильной кислоты, чем угольная,
- диссоциирующие от оксигемоглобина ионы Н + нейтрализуют поступающий из-вне ион HCO3 – с образованием угольной кислоты. После карбоангидразной реакции образуется СО2, который выводится наружу.
Источник
234. Транспорт углекислоты кровью, значение карбоангидразы, взаимосвязь транспорта о2и со2.
Углекислый газ транспортируется следующими путями:
Растворенный в плазме крови — около 25 мл / л.
Связанный с гемоглобином (карбгемоглобин) — 45 мл / л.
В виде солей угольной кислоты — букарбонаты калия и натрия в плазме крови — 510 мл / л.
Таким образом, в состоянии покоя кровь транспортирует 580 мл углекислого газа в 1 л. Итак, основной формой транспорта СО2 является бикорбонаты плазмы, образующихся благодаря активному протеканию карбоангидразнои реакции.
В эритроцитах содержится фермент карбоангидраза (КГ), который катализирует взаимодействие углекислого газа с водой с образованием угольной кислоты, распадается с образованием бикарбонатного иона и протона. Бикарбонат внутри эритроцита взаимодействует с ионами калия, выделяемых из калиевой соли гемоглобина при восстановлении последнего. Так внутри эритроцита образуется бикарбонат калия. Но бикарбонатно ионы образуются в значительной концентрации и поэтому по градиенту концентрации (в обмен на ионы хлора) поступают в плазму крови. Так в плазме образуется бикарбонат натрия. Протон, образовавшегося при диссоциации угольной кислоты, реагирует с гемоглобином с образованием слабой кислоты ННb.
В капиллярах легких эти процессы идут в обратном направлении. С ионов водорода и бикарбонатных ионов образуется угольная кислота, которая быстро распадается на углекислый газ и воду. Углекислый газ удаляется наружу.
Итак, роль эритроцитов в транспорте углекислоты такова:
образование солей угольной кислоты;
Диффузия газов в тканях подчиняется общим законам (объем диффузии прямо пропорционален площади диффузии, градиента напряжения газов в крови и тканях). Площадь диффузии увеличивается, а толщина диффузного слоя уменьшается при увеличении количества функционирующих капилляров, что имеет место при повышении уровня функциональной активности тканей. В этих же условиях возрастает градиент напряжения газов за счет снижения в активно работающих органах Ро2 и повышения Рсо2 (газовый состав артериальной крови, как и альвеолярного воздуха остается неизменным!). Все эти изменения в активно работающих тканях способствуют увеличению объема диффузии О2 и СО2 в них. Потребление О2 (СО2) по спирограмму определяют по изменению (сдвигу) кривой вверх за единицу времени (1 минуту).
235. Иннервация дыхательных мышц.
Дыхательный центр, расположенный в продолговатом мозге, посылает импульсы к мотонейронам спинного мозга, иннервирующим дыхательные мышцы. Диафрагма иннервируется аксонами мотонейронов, расположенных на уровнеIII—IV шейныхсегментовспинного мозга. Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположеныв передних рогах (III—XII) грудных сегментовспинного мозга.
236. Дыхательный центр. Современные представления о структуре и локализации. Автоматия дыхательного центра.
Информация о состоянии кислородно-углекислого баланса в организме поступает в дыхательный центр, который представляет нейронную организацию центральной нервной системы, определяющую функцию дыхания.
В анатомическомсмысле дыхательный центр– это совокупность нейронов в локальной зоне центральной нервной системы, без которой дыхание становится невозможным.
Такой центр находится в ретикулярной формации продолговатого мозга в областидна IV желудочка.
Он состоит из двух отделов:
1) центр вдоха(инспираторный отдел);
2) центр выдоха(экспираторный отдел).
Нейроны бульбарного центра обладают автоматией и находятся в реципрокных взаимоотношениях между собой.
Несовершенность координации дыхательного акта центрами продолговатого мозга была доказана методом перерезок. Так после отделения продолговатого мозга от вышележащих отделов чередование вдохов и выдохов сохраняется, но длительность и глубина дыханий становится нерегулярной.
В физиологическомсмысле дыхательный центр– это совокупность нейронов, расположенных на различных уровнях центральной нервной системы (от спинного мозга до коры головного мозга), которые обеспечивают координированное ритмическое дыхание, то есть делают функцию дыхания более совершенной.
В целом, регуляция активности дыхательного центра может быть представлена тремя уровнями:
1) на уровне спинного мозгарасполагаются центры диафрагмальных и межрёберныхнервов, обусловливающие сокращение дыхательных мышц.Однако этот уровень регуляции дыхания не может обеспечить ритмическую смену фаз дыхательного цикла, так как большое количество афферентных импульсов от дыхательного аппарата непосредственно направляются в продолговатый мозг, то есть минуя спинной мозг.
2) на уровне продолговатого мозга и варолиевого мостанаходится основной дыхательный центр, который перерабатывает разнообразные афферентные импульсы, идущие от дыхательного аппарата, а также от основных сосудистых рефлексогенных зон. Этот уровень регуляции обеспечивает ритмическую смену фаз дыхания и активность спиномозговых мотонейронов, аксоны которых иннервируют дыхательную мускулатуру;
3) на уровне верхних отделов головного мозга, включая кору головного мозга, осуществляются адекватные приспособительные реакции системы дыхания к изменяющимся условиям окружающей среды.
Ритмические импульсы от дыхательного центра продолговатого мозга поступают по нисходящим двигательным путям к мотонейронам дыхательных мышц спинного мозга.
Мотонейроны диафрагмальных нервовнаходятся в передних рогах серого веществаIII—IV шейных сегментов.
Мотонейроны межрёберных нервоврасположены в передних рогахгрудного отделаспинного мозга.
Отсюда возбуждение поступает к дыхательной мускулатуре (к диафрагме и межрёберным мышцам).
Мотонейроны спинного мозгаполучают от проприорецепторов мышц грудной клетки сигналы о степени их растяжения при вдохе.
Эти сигналы могут изменять число вовлечённых в активность мотонейронов и, таким образом, определяют особенности дыхания, осуществляя регуляцию дыхания на уровне спинного мозга
Бульбарный дыхательный центрполучает афферентные импульсы от механорецепторов лёгких, дыхательных путей и дыхательных мышц, от хемо- и прессорецепторов сосудистых рефлексогенных зон.
Мотонейроны спинного мозгаполучают от проприорецепторов мышц грудной клетки сигналы о степени их растяжения при вдохе.
Эти сигналы могут изменять число вовлечённых в активность мотонейронов и, таким образом, определяют особенности дыхания, осуществляя регуляцию дыхания на уровне спинного мозга
Бульбарный дыхательный центрполучает афферентные импульсы от механорецепторов лёгких, дыхательных путей и дыхательных мышц, от хемо- и прессорецепторов сосудистых рефлексогенных зон.
Для нормальной деятельности бульбо-понтинногодыхательного центра необходима постоянная информация о состоянии внутренней среды организма и самих органов дыхания.
Нисходящие нервные влияния на дыхательный центр оказывают верхние отделы головного мозга, включая корковые нейроны. Так, эмоциональные возбуждения, охватывающие структуры,лимбико-ретикулярного комплексаи в первую очередьгипоталамическую область, распространяются в нисходящем направлении и вызывают изменение деятельности дыхательного центра.
Гипоталамустакже оказывает влияния при изменениях во внешней среде, изменении метаболизма, а также как высший центр вегетативных регуляций.
Речь, относящаяся к высшим мозговым функциям корычеловека, возможна на основе дыхательных движений, вызывающих прохождение воздуха через голосовой аппарат.
Поэтому во время речи к дыхательному центру приходят влияния, подстраивающие его деятельность для необходимых речевых реакций.
Одновременно дыхательный центр управляет тем объёмом лёгочной вентиляции, который необходим для поддержания дыхательного гомеостаза. Поэтому дыхание в условиях речи становится апериодическим.
На роль корыв регуляции дыхания указывает возможность произвольного контроля дыхания, когда человек может сознательно изменить дыхание: сделать его более глубоким или поверхностным, частым или редким, произвести задержку дыхания на определённое время.
Таким образом, на примере особенностей дыхательного центра наблюдаются общие принципы организации любых нервных центров, в частности:
1) принцип изоморфизма(принципиально однотипная структурная организация);
2) принцип иерархичности(многоуровневое расположение центрального представительства);
3) принцип субординации(соподчинение нервных центров, когда высшие центры модулируют работу низших и, чем выше уровень центра, тем более сложную регуляцию он обеспечивает).
Источник



