
- Категории статей
- Ученые ставят диагноз планете
- Природный регулятор температуры колибри
- Биоразлагаемые пакеты – вред или польза?
- Видео лекции на канале Temperatures.ru
- Теплэко – тепло из ниоткуда?
- Популярные статьи
- Польза и вред инфракрасного обогревателя (323421)
- Почему горячая вода замерзает быстрее, чем холодная? (209331)
- Вредно ли разогревать пищу в микроволновке? (199115)
- Контролируйте температуру приготовления мяса! (181235)
- 451 градус по Фаренгейту, температура возгорания бумаги? (165463)
- Основные разделы
- Парадокс теплообмена или почему у птиц не мерзнут лапы
- Особенности терморегуляции сельскохозяйственной птицы
- Лекция 11. Терморегуляция. Сезонные и суточные ритмы.
Категории статей
Ученые ставят диагноз планете
Cтолько углекислого газа, как сейчас, в атмосфере не было последние 2 млн лет, метана и закиси азота — 800 тыс. лет. Далее
Природный регулятор температуры колибри
Учитывая огромную скорость и частоту крыльев, птицы должны нагреваться до температур, несовместимых с жизнью. Далее
Биоразлагаемые пакеты – вред или польза?
Интересно разобраться, действительно ли такие пакеты не наносят вреда окружающей природе. Далее
Видео лекции на канале Temperatures.ru
Две видео лекции уже доступны для просмотра на канале Temperatures.ru Далее
Теплэко – тепло из ниоткуда?
К рекламе на телевидении нужно относиться очень критически, особенно когда её представляют умные люди. Далее
Популярные статьи
Польза и вред инфракрасного обогревателя (323421)
Среди электрических обогревателей, которые мы используем в быту, наиболее популярными сейчас становятся инфракрасные нагреватели. Они очень широко рекламируются в Интернете и в газетах. Говорят, что они намного эффективнее масляных радиаторов и тепловентиляторов. Меньше потребляют энергии, не сжигают кислород и т.д. Главное – они совершенно не вредные, никакого отрицательного воздействия на организм человека не оказывают. Далее
Почему горячая вода замерзает быстрее, чем холодная? (209331)
Это действительно так, хотя звучит невероятно, т.к в процессе замерзания предварительно нагретая вода должна пройти температуру холодной воды. Парадокс известен в мире, как «Эффект Мпембы». Далее
Вредно ли разогревать пищу в микроволновке? (199115)
Одна моя знакомая отказывается есть пищу, которую кто-то разогрел в микроволновой печи. Всему виной — страшилки в Интернете. Далее
Контролируйте температуру приготовления мяса! (181235)
При приготовлении сырого мяса, особенно, домашней птицы, рыбы и яиц необходимо помнить, что только нагревание до надлежащей температуры убивают вредные бактерии. Далее
451 градус по Фаренгейту, температура возгорания бумаги? (165463)
451 градус по Фаренгейту. Это название знаменитой книги Рэя Брэдбери. На языке оригинала звучит так: ‘Fahrenheit 451: The Temperature at which Book Paper Catches Fire, and Burns’. Действительно ли при этой температуре начинают гореть книги? Далее
Основные разделы
Парадокс теплообмена или почему у птиц не мерзнут лапы
На нашем сайте можно найти несколько статей о температурных и тепловых парадоксах. (см. ,например, фотометрический парадокс, парадокс Мпембы). Не можем обойти вниманием еще один тепловой парадокс, который вошел в «Золотой фонд» элементарной физики по мнению Московского физического центра. Это парадокс теплообмена. Также его называют «встречный теплообменник».
Классическая постановка задачи, приведенная в книге В. Н. Ланге «Физические парадоксы, софизмы и занимательные задачи» звучит следующим образом: Имеются три одинаковых изолированных от среды сосуда А, Б, В. В первых двух налито по одному литру воды при температуре 80 и 20 °С. Также имеется пустой сосуд Г меньшего размера с тонкими, абсолютно теплопроводными стенками. Можно ли, используя все эти сосуды, нагреть при помощи горячей воды холодную так, чтобы ее конечная температура стала выше конечной температуры горячей воды? Смешивать воду А и Б не разрешается.
Ответ, разумеется, можно! На то он и парадокс. Хотя, на первый взгляд, возникают подозрения о нарушении второго закона термодинамики. Решение задачи – в подогреве воды «по частям», для этого и даны дополнительные сосуды.
Половину холодной воды отольем в сосуд Г и поместим его внутрь сосуда А с горячей водой. В отсутствии тепловых потерь в сосудах А и Г наступит равновесие при температуре примерно 60 °С. Перельем подогретую воду из сосуда Г в пустой изолированный сосуд В и повторим процедуру. Выльем остаток холодной воды из Б в Г и опять погрузим Г в А. Вода в А и Г примет температуру около 47 °С. Если теперь перелить воду из Г в В, температура в В понизится до 53 °С, но при этом окажется большей, чем температура в сосуде А! Таким образом задача выполнена. Холодная вода оказалась нагретой до 53 °С за счет охлаждения горячей воды до 47 °С. При этом никакого нарушения законов теплообмена нет, т.к. температура жидкости, которой передается тепло, всегда меньше, чем температура жидкости, отдающей тепло.
Если разделить воду не на две, а на большее число частей, то разница конечных температур будет больше. На этом физическом явлении основана работа встречного теплообменника, в котором по соосно расположенным трубкам текут навстречу друг другу два потока жидкости – холодный и горячий. При достаточно длинных трубках происходит фактически «обмен температурами» между холодной и горячей водой. Если бы потоки двигались в одном направлении, то их температуры, в лучшем случае, только бы выравнивались. КПД встречных теплообменников очень высокий.

Явление встречного теплообмена через тонкостенную трубку и встречного обмена концентрации жидкостей через мембрану широко используется в науке и индустрии. Оно также очень распространено в природе.
В холодную погоду поток крови к конечностям птиц и млекопитающих снижается, кровь возвращается к внутренним органам через глубокие вены, при этом подкожные вены конечности плотно сжаты. Поскольку глубокие вены у птиц тесно переплетены с артериями, образуется встречная система теплообмена, которая переносит тепло от артериальной крови в венозную кровь, возвращающуюся в организм подогретой, что приводит к минимальным потерям тепла через конечности в холодную погоду. Птицы и млекопитающие, которые регулярно погружают свои конечности в холодную или ледяную воду, особенно хорошо развивают встречные системы кровотока, что позволяет переносить длительное воздействие холода без значительной потери тепла тела, даже когда конечности являются такими тонкими как, например, нижние ноги или лапки птицы.

Еще один интересный факт. Поскольку явление встречного теплообмена универсально и очень полезно, есть попытки связать его с константами «золотого сечения», которые, как известно, управляют гармонией природы. В статье С.Л. Василенко «Водные парадоксы» приводится следующее наблюдение. Так, если принять, что первоначально холодная вода имеет температуру 0 °С, а горячая 100 °С, разделить холодную воду на 12 частей, и рассчитать до какой температуры может охладиться горячая вода, то получим 38,2 °С. Как известно, 0,382 – это константа «золотого сечения».

Надо сказать, что привязка к законам гармонии довольно искусственная, поскольку, как мы знаем, значение температуры 100 °С для кипения воды было выбрано Цельсием произвольно, в шкале Фаренгейта оно составляет 212 градусов. Но наблюдение довольно интересное.Самое интересное в любом физическом парадоксе – это как люди используют открытое явление природы для своих технических и научных целей, а также для объяснения функционирования систем теплообмена в живых организмах.
Источник
Особенности терморегуляции сельскохозяйственной птицы

Важной особенностью сельскохозяйственной птицы является более высокая (41-42°С), чем у животных, нормальная температура тела. Это связано с тем, что обмен веществ в организме птицы происходит более интенсивно, чем у животных. Это также значит, что организм птицы производит большее количество тепла
Чем больше живая масса птицы, тем больше тепла она производит.
Зимой это помогает сократить расходы на обогрев, а летом при высокой внешней температуре это становится большой проблемой.
Нормальное функционирование всех систем организма птицы возможно только в том случае, когда количество отведенного тепла равно количеству произведенного.
В отличие от человека, птица не имеет потовых желез, поэтому не может отводить тепло через пот. Через кожу отводится лишь незначительная часть тепла, а основной способ теплоотдачи происходит путем испарения воды из организма в процессе дыхания.
Поэтому в жару наблюдается учащенное дыхание у птицы (с 20-24 до 250 циклов в мин.) Частое дыхание в течение нескольких часов ведет к гипервентиляции легких, при этом снижается уровень углекислого газа (СО2) в крови птицы. Недостаток СО2 способствует изменению кислотно-щелочного равновесия организма (pH>7), из-за чего наступает состояние респираторного алкалоза и, как следствие, массовый падеж.
При высокой внешней температуре потребление воды птицей увеличивается более чем в 3 раза. Вследствие этого часть съеденного корма транзитом выводится из организма, не успев усвоиться. Более того, в жару птица уменьшает потребление корма, т.к. в процессе переваривания организмом будет выделяться еще больше теплоты.
В жару при увеличении потребление воды птицей и уменьшении поедания корма снижается снабжение организма питательными и минеральными веществами, уменьшаются привесы.
Чтобы этого избежать, следует соблюдать «зону температурного комфорта», когда птица чувствует себя комфортно и не испытывает негативные факторы окружающей среды: слишком жарко, слишком сухо или слишком холодно. В «зоне температурного комфорта» энергия птицы не растрачивается на терморегуляцию тела.
При создании комфортной температуры для содержания птицы следует учитывать немаловажный фактор влажность воздуха в помещении, так как при разной влажности должна быть разная температура воздуха.
Ниже представлена таблица комфортной температуры внутри птичника при различных значениях относительной влажности воздуха из руководства по выращиванию бройлеров кросса «КОББ».
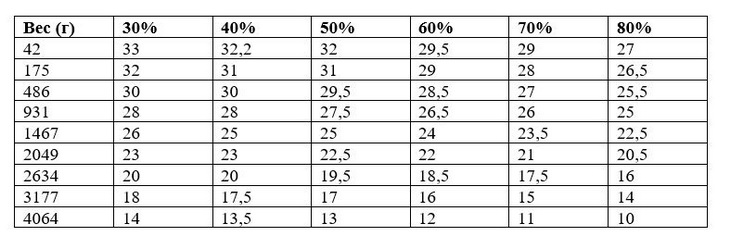
Из таблицы видно, что для молодой птицы весом 486 гр. при относительной влажности воздуха 30% «зона температурного комфорта» составляет 30°С, а при влажности 80% – 25,5°С. В обоих случаях птица будет чувствовать себя одинаково.
Чем выше относительная влажность воздуха, тем меньше должна быть температура внутри птичника!
В первые дни после посадки, чтобы у цыплят не возник тепловой стресс, рекомендуется поддерживать влажность воздуха на уровне 60-70%, близкую к уровню влажности в инкубаторе (около 80%), например, с помощью системы орошения в птичнике.
Комфортная температура в птичнике позволит снизить затраты на обогрев здания, а также поможет избежать падежа стада, что положительно скажется на себестоимости продукции.
Источник
Лекция 11. Терморегуляция. Сезонные и суточные ритмы.
Будучи гомойотермными животными, птицы характеризуются высокой и устойчивой температурой тела. Температура тела у различных видов птиц в норме составляет около 41 °С с колебаниями от 38 до 43,5 °С; такие колебания намного меньше, чем у млекопитающих. Кроме того, очень высока устойчивость температуры тела. Даже у арктических видов при температуре среды до 20-50 °С ниже нуля колебания температуры тела не превышают 2°С. То же можно сказать и о видах, обитающих в жарких, засушливых условиях.
Регуляция теплообмена может осуществляться путем адаптивных изменений уровня теплопродукции и (или) теплоотдачи.
У птицвысок уровень окислительно-восстановительных реакций, сопровождающихся выделением тепла, поэтому у них высокая и стабильная температура тела. Процесс рефлекторного усиления теплопродукции в ответ на снижение температуры окружающей среды носит название химической терморегуляции.
Выделение энергии в виде тепла при работе различных органов свойственно всем животным, в том числе и пойкилотермным. Специфика гомойотермных животных состоит в том, что они способны переносить значительные колебания температуры внешней среды без существенных изменений уровня их жизнедеятельности и активная работа внутренних органов или мышц также не ведет к значительному перегреванию организма. Терморегуляторное образование идет преимущественно в скелетной мускулатуре. Здесь следует различать тепло, выделяемое при работе мышцы и специфические терморегуляторные изменения теплопродукции. Повышение теплообразования при охлаждении может происходить в покоящейся, не проявляющей видимых признаков деятельности мышце.
Наиболее обычные физиологические механизмы усиления теплопродукции — «терморегуляторный тонус» и дрожь. Терморегуляторный тонус представляет собой микросокращения во внешне покоящейся мускулатуре, при этом повышается электрическая активность мышцы, потребление кислорода повышается на 150%. При более сильном охлаждении наряду с резким повышением терморегуляционного тонуса в процесс теплообразования включается видимая сократительная деятельность мышц в форме холодовой дрожи. Газообмен мышц при этом возрастает до 300-400%.
При длительном воздействии холода этот (сократительный) тип термогенеза может быть замещен (или дополнен) включением «свободного» (нефосфорилирующего) тканевого дыхания, при котором выпадает фаза образования и последующего разложения АТФ. Биологическое преимущество нефосфорилирующего окисления заключается в том, что большая часть энергии сразу расходуется в виде тепла, окислительные процессы не могут быть заторможены недостатком АТФ и неорганического фосфата.
При изменении температуры среды меняется и уровень теплообразования. В общей форме эти изменения могут быть представлены кривой. В определенном интервале температур внешней среды теплопродукция, соответствующая основному обмену организма, полностью скомпенсирована его «нормальной» теплоотдачей. Эта температурная зона носит название «термонейтральной зоны»; уровень обмена минимален. Нередко говорят о «критической точке», подразумевая конкретную температуру, при которой достигается тепловой баланс со средой.
Понижение температуры за пределы термонейтральной зоны вызывает рефлекторное повышение уровня обмена веществ, а соответственно и теплопродукции. Этот процесс соответствует левой части кривой.
Повышение температуры среды за пределы термонейтральной зоны также вызывает увеличение уровня обмена веществ и теплопродукции, на этот раз вызванное затратами энергии на усиление теплоотдачи (физической терморегуляции). Эти механизмы будут рассмотрены ниже. Обе системы терморегуляции действенны в определенном для каждого вида диапазоне температур, после чего наступают патологические изменения, вызванные перегревом (или переохлаждением) организма, и гибель.
Межвидовые отличия химической терморегуляции в значительной степени связаны с разницей в размерах тела, определяющих общий уровень метаболизма, но отражают и экологическую специфику вида. Так, зерноядные птицы характеризуются несколько меньшей интенсивностью химической терморегуляции по сравнению с насекомоядными; оседлые птицы существенно более реактивны, чем перелетные, не подвергающиеся действию суровых зимних температур.
Сезонные изменения уровня теплопродукции. Более это характерно для оседлых видов, живущих при широком годовом колебании температур. В годовом цикле изменяется положение термонейтральной зоны. Для оседлых видов зимой характерно расширение диапазона термонейтральной зоны и сдвиг критической точки в сторону более низких температур, тогда как у перелетных птиц эти явления выражены менее отчетливо или отсутствуют совсем.
Такая «настройка» организма на поддержание минимального обмена при более низких температурах среды приводит к тому, что терморегуляторное повышение теплопродукции начинается при более низких, чем летом, температурах, что определяет более экономный расход энергии.
Более высокий уровень химической терморегуляции в зимнее время требует адаптаций, обеспечивающих энергетику этого процесса. Такие адаптации выражены, к примеру, в сезонном накоплении жира у зимующих в холодных зонах птиц.
Физическая терморегуляция.Химическая терморегуляция может обеспечить только кратковременное пребывание в дискомфортной температуре. Стойкие приспособления к длительному пребыванию в дискомфортных температурных условиях невозможны без сочетания адаптивных изменений теплопродукции с устойчивыми морфофизиологическими механизмами, определяющими уровень теплоотдачи организма.
Изменения уровня теплоотдачи могут быть приспособлением как к понижению температуры среды по сравнению с оптимумом, так и к ее повышению. Различны и механизмы терморегуляции.
Птицы приспособлены к низким температурам благодаря в первую очередь развитию перьевого покрова. Перья удерживают вокруг тела птицы некоторое количество воздуха, препятствующего оттоку тепла и таким образом снижающего уровень энергетических затрат на поддержание температуры тела. Структура оперения соответствует этой функции. У птиц, зимующих в умеренных и высоких широтах, имеют место сезонные изменения густоты и структуры оперения, определяющие повышение его теплоизоляционных свойств в зимнее время.
Все вышесказанное – это устойчивые свойства перьевого покрова, определяющие общий уровень теплоизоляции организма. Подвижность перьев открывает возможность и для лабильной регуляции теплоотдачи при быстрых и иногда кратковременных изменениях. Изменение положения перьев по отношению к поверхности тела меняет толщину теплоизолирующей воздушной прослойки. Это называется «птиломоторная» реакция. Степень распушенности оперения меняется в зависимости от активности птицы и температуры окружающей среды. Спокойно сидящая птица взъерошивает оперение тем сильнее, чем ниже температура воздуха, но при активных движениях и кормлении мускульные сокращения и действие пищи дают дополнительное тепло, и оперение приглаживается, давая путь к увеличению теплоотдачи. Увеличение теплоотдачи может достигаться и противоположным путем – максимальным распущенней перьев, при котором вершины их не прилегают друг к другу, и устойчивость воздушной прослойки нарушается. Такое положение перьев чаще всего наблюдается при перегреве, особенно – при патологическом повышении температуры тела.
Терморегуляция при действии высоких температур направлена в основном на усиление теплоотдачи. Наиболее важный механизм при этом – усиление теплоотдачи путем интенсификации испарения влаги. Хотя у птиц не имеется потовых желез, у ряда видов отмечено испарение воды с поверхности кожи. Однако кожное испарение, видимо, не подвержено активной регуляции, поэтому терморегуляторная влагоотдача связана почти исключительно с усилением вентиляции ротовой полости и верхних дыхательных путей. Увеличение испарения влаги может достигаться при этом либо путем учащения дыхания (терморегуляторное полипноэ), либо особыми колебательными движениями кожи нижней стороны шеи (горловая дрожь), производимыми с участием гиоидного аппарата. Чаще оба механизма действуют вместе.
В комплекс механизмов физической терморегуляции входят и сосудистые реакции. Сосудистая регуляция может служить приспособлением и к повышению, и к понижению температуры среды. В первом случае адаптивный ответ организма заключается в расширении кровеносных сосудов, расположенных близко к поверхности кожи, что ведет к усилению отдачи тепла во внешнюю среду. Во втором – происходит сжатие поверхностных и расширение глубоколежащих сосудов, что вызывает большую консервацию тепла в организме. Большее значение в терморегуляции имеют неоперенные участки тела, с поверхности которых может осуществляться интенсивная отдача тепла.
Специальная функция регуляции теплоотдачи присуща лишенным оперения задним конечностям птиц. Расширение расположенных здесь сосудов приводит к усилению теплоотдачи. Сосудистые реакции в конечностях птиц используются и как механизм сохранения тепла. Установлено, что артериальные и венозные сосуды, подходящие к конечностям, образуют сплетения, действующие по принципу противоточного теплообменника. Протекающая по этим сплетениям артериальная кровь отдает часть своего тепла возвращающейся из охлажденных конечностей венозной крови, при этом в свою очередь охлаждаясь. Таким образом, конечности снабжаются охлажденной кровью, что уменьшает их температурный градиент со средой и резко ограничивает рассеивание тепла. Этот тип сосудистых реакций особенно характерен для арктических птиц.
Терморегуляция в активном полете.Все сказанное о терморегуляторных реакциях у птиц справедливо для животных, находящихся в состоянии покоя или относительно невысокой активности. Как осуществляется регуляция теплообмена при активном полете, выяснено недостаточно.
Машущий полет связан с интенсивной деятельностью мускулатуры и, соответственно, с резким повышением уровня обмена веществ и теплопродукции. Уровень метаболизма летящих птиц в 6-8 раз превосходит обмен покоя. Исходя из этих данных, можно предположить, что химическая терморегуляция при активном полете не играет существенной роли. Показано, что в течение активного полета температура тела птиц стойко или спонтанно повышается на 1,5-4°С.
Главная биологическая задача терморегуляции во время активного полета заключается в усилении теплоотдачи. Во время полета учащается дыхание, при этом увеличивается испарительная влагоотдача. Однако чрезмерный расход воды при длительном (например, миграционном) полете биологически невыгоден. Видимо, поэтому испарительная влагоотдача в полете у большинства исследованных видов относительно невелика и повышается лишь при высокой температуре воздуха.
В гораздо большей степени отдача тепла в полете осуществляется конвекцией и радиацией с поверхности тела. Плотно прижатое к телу оперение облегчает этот процесс. Такое положение перьев вызвано необходимостью создания обтекаемой поверхности, что выгодно и для терморегуляторной теплоотдачи. Увеличению теплоотдачи в полете может способствовать и то, что при расправленном положении крыльев оказываются открытыми менее защищенные перьями участки тела.
Активная терморегуляторная роль сосудистых реакций у птиц отмечена для неоперенной части задних конечностей. Опыты с чайками в аэродинамическом приборе показали, что температура поверхности ног уменьшалась при снижении температуры воздуха и увеличивалась при ее повышении.
Нервный контроль. Поведение.Терморегуляторные реакции находятся под непосредственным контролем центральной нервной системы по принципу обратных связей. Исходным звеном этих связей служит информация о температуре внешней среды и температуре тела. Холодовые рецепторы расположены на языке и на клюве и представляют собой свободные нервные окончания. Вообще терморецепторы птиц изучены недостаточно. Информация с терморецепторов направляется в центральную нервную систему, где она, видимо, дополняется информацией об изменениях внутренней температуры, а именно температуры крови в сосудах спинного и головного мозга.
Повышение или понижение температуры крови (и, соответственно, спинного мозга) вызывает отчетливые адекватные терморегуляторные реакции. В эксперименте при повышенной температуре спинного мозга у птиц развивалась тепловая одышка, местное охлаждение спинного мозга вызывало снижение частоты дыхания до нормы.
Интегрированный терморегуляторный ответ целого организма осуществляется на уровне гипоталамической области головного мозга. В эксперименте охлаждение гипоталамической области вызывало повышение потребления кислорода и подъем температуры тела; при нагревании наблюдался противоположный эффект. Показано, что птиломоторные и поведенческие терморегуляционные механизмы находятся под преимущественным контролем гипоталамуса, а холодовая дрожь (и, видимо, другие формы термогенеза) – спинного мозга. Тепловая одышка подчинена контролю со стороны блуждающего нерва.
На уровне высшей нервной деятельности приспособления птиц к температурным условиям среды осуществляются в виде особых форм адаптивного поведения; это явление нередко называют «экологической терморегуляцией».
Один из видов такой терморегуляции заключается в использовании особенностей микроклимата при размещении по стациям и ярусам, выборе места для ночевки и пр. Например, в лесных биотопах при дождливой и ветреной погоде птицы ищут убежища в участках хвойного леса, в густых кронах, кустарниках. В Антарктике распределение колоний пингвинов, глупышей, буревестников и некоторых других птиц приурочено к понижениям рельефа или местам, находящимся под прикрытием скалистых уступов.
Большое значение для энергообмена птиц имеет выбор мест для ночевки, особенно в холодную погоду. Одним из примеров могут служить ночевки тетеревиных птиц под снегом. Помимо тетеревиных птиц подснежные ночевки свойственны пуночкам, чечеткам, ряду видов жаворонков, седому и черному дятлам и некоторым другим видам. Использование для ночевки густых крон, зарослей тростника, дупел и других убежищ дает тот же биологический эффект. Для защиты от холода многие птицы образуют скопления. Например, пищухи в холодную погоду собираются в укрытых местах группами до 20 особей. Длиннохвостые синицы зимой с наступлением сумерек скапливаются в густых кустарниках; здесь птицы, взъерошивая оперение, устраиваются на ветвях, тесно прижимаясь друг к другу. Возможны и групповые ночевки под снегом. Это показано, например, для серых куропаток. Один из самых ярких примеров роли скоплений – императорские пингвины. Показано, что самцы императорского пингвина, столпившиеся в большие группы, имеют среднюю температуру тела 35,7 °С, птицы в небольших группах – 36,9, тогда как температура тела у одиночных особей равна в среднем 37,9°С.
Перечисленные формы приспособительного поведения способствуют переживанию неблагоприятных температур с меньшей напряженностью энергетического обмена. Однако затраты на терморегуляцию остаются довольно высокими, что вызывает необходимость интенсивного питания. В условиях короткого зимнего дня птицы заняты сбором корма практически все светлое время суток, часто расширяя период активности за счет сумеречного времени. Многим птицам в этот период свойственно групповое питание, при котором расширяются возможности отыскания корма. Для ряда видов, зимующих в районах холодного климата, свойственны различные формы запасания кормов.
В зависимости от питания изменяются и поведенческие температурные адаптации. Например, в Якутии врановые птицы, питаясь высококалорийными кормами, постоянно поддерживают высокий уровень метаболизма и температуры тела. Тетеревиные птицы, питание которых менее калорийно и уровень метаболизма гораздо ниже, используют другие экологические механизмы (теплые убежища в снегу) для переживания периодов с низкими температурами внешней среды.
Птицы, обитающие в жарком, засушливом климате, тесно связаны с необходимостью пополнения водных запасов, поскольку испарение влаги представляет собой эффективный путь приспособления к высокой температуре. Наличие водопоев – важное условие существования большинства птиц, особенно обитающих в аридных условиях. Для последних возможность регулярного утоления жажды расширяет период активности и на жаркие часы суток.
Общие принципы адаптации теплообмена.У птиц существуют два принципиальных пути приспособления к температурным условиям окружающей среды. Первый из них представляет собой систему стабильных температурных адаптаций организма к среднему уровню температуры среды, сохраняющемуся в течение достаточно длительного времени. Таковы, например, сезонные особенности теплообмена. У птиц ведущее значение в сезонных изменениях теплообмена имеет смена уровней теплоотдачи, определяемая изменениями теплоизолирующих свойств перьевого покрова. Эти изменения теплоотдачи служат основой сезонных сдвигов теплопродукции (положение критической точки, диапазон термонейтральной зоны, уровень основного обмена).
Второй путь адаптивных изменений теплообмена представлен лабильными реакциями терморегуляции, обеспечивающими приспособление организма к быстрым, относительно кратковременным отклонениям температуры среды от ее среднего уровня, свойственного данному сезону. У птиц преимущественную роль в такого рода приспособлениях играет химическая терморегуляция при воздействии низких температур и активная регуляция респираторной влагоотдачи при действии высоких температур. Как уже указывалось, морфофизиологические адаптации птиц дополняются сложными и многообразными формами приспособительного поведения.
Принципиальная возможность изменения «настройки» терморегуляторных механизмов на поддержание обмена веществ на разном уровне лежит в основе весьма интересного явления обратимой гипотермии, обнаруженного у некоторых видов птиц.
Обратимая гипотермия у птиц может быть выражена в трех формах. У ряда видов наблюдается нерегулярное оцепенение, вызванное резкими похолоданиями, дождями, снегопадами и другими снижающими успешность питания факторами, и прекращающееся по мере восстановления благоприятных условий. Такой характер обратимой гипотермии присущ стрижам и ласточкам. Эти птицы при наступлении неблагоприятных условий скапливаются группами (иногда очень многочисленными) в различного рода убежищах, снижают активность и впадают в глубокое оцепенение, не проявляя никаких внешне видимых признаков жизни.
Вторая форма обратимой гипотермии представлена регулярными суточными циклами нормо- и гипотермии. Это явление наиболее характерно для колибри, которые впадают в оцепенение в ночное время, возвращаясь к активному состоянию утром. В состоянии оцепенения птицы совершенно неподвижны, глаза их закрыты, перья часто бывают взъерошены, дыхание неравномерно, дыхательные движения часто внешне совсем незаметны.
Третья форма естественной обратимой гипотермии представлена сезонным впадением в оцепенение, вполне аналогичным им ней спячке млекопитающих. Это явление достоверно описано лишь для одного вида козодоев – гуахаро. Сходство сезонного оцепенения гуахаро со спячкой млекопитающих подтверждается тем, что для него показано накопление жировых запасов, предшествующее сезонной гипотермии.
Биологическое значение естественной обратимой гипотермии у птиц и разнообразие ее форм определяются экологией рассматриваемых видов: для ласточек и стрижей длительное понижение температуры воздуха, дожди, ветер резко ухудшают условия питания. Снижение уровня метаболизма в этом случае биологически оправдано. У перелетных видов такие условия в природной обстановке встречаются нерегулярно; у них отсутствует и регулярность впадения в гипотермию. Гуахаро сталкивается с сезонным периодом неблагоприятных условий, поэтому, видимо, и возникла сезонность впадения в гипотермию. У колибри неблагоприятная погода также ухудшает условия питания. Кроме того, мелкие размеры этих птиц ведут к тому, что расход энергии в течение ночи может привести к серьезным нарушениям теплового баланса.
Что касается физиологической характеристики состояния, обратимой гипотермии, то установлено, что это состояние существенно отличается от холодового оцепенения пойкилотермных животных. У рассмотренных видов птиц не отмечается прямой зависимости перехода в гипотермическое состояние от температуры среды. У видов с ритмическим впадением в гипотермию снижение обмена веществ и температуры тела регулируется временем суток на основе фотопериодической реакции и происходит подчас на фоне неизменной и довольно высокой температуры воздуха. У птиц с нерегулярной гипотермией важным условием впадения в оцепенение являются определенная степень истощения и потеря веса, вызванные недостатком пищи. При сезонной гипотермии, как уже говорилось, необходима физиологическая подготовка организма в виде определенной степени ожирения.
Процесс выхода из гипотермии также не имеет прямой связи с изменениями температуры среды. Повышение обмена при суточных циклах регулируется ритмикой освещения или эндогенными циклами и может начинаться при низкой (не отличающейся от ночной) температуре воздуха. Скорость повышения температуры тела (порядка 0,2-0,5 у стрижей и 1-1,5° в минуту у колибри) несопоставима с масштабами изменения температуры воздуха. У выходящих из состояния гипотермии птиц наблюдается мышечная дрожь, активные движения, регистрируется быстрое повышение интенсивности потребления кислорода и взъерошивание оперения. Все это свидетельствует об активном характере выхода из гипотермии.
Онтогенез терморегуляции.Морфофизиологические механизмы регуляции теплообмена возникают в онтогенезе птиц не сразу. Их формирование связано с общим развитием организма, поэтому сроки появления эффективной терморегуляции сильно отличаются у видов с разным типом онтогенеза.
У видов с птенцовым (имматуронатным) типом развития птенцы в течение некоторого периода после вылупления не обладают способностью к регуляции теплообмена. Интенсивность обмена веществ в их организме, а соответственно и температура тела находятся в прямой зависимости от температуры внешней среды, что уподобляет их пойкилотермным животным. Это сходство дополняется высокой устойчивостью к пониженной температуре, способностью обратимо снижать уровень обмена при понижении температуры тела вплоть до полного исчезновения внешних признаков жизни.
Появление химической терморегуляции, т. е. переход к гомойотермному типу реакции, происходит в различные сроки у разных видов и сопровождается общим подъемом уровня обмена веществ. Переход птенцов к гомойотермному типу реакции совершается «скачкообразно» в очень сжатые сроки. Такой быстрый переход из одного качественного состояния в другое может произойти только при условии предварительной подготовки всех морфофизиологических механизмов, обеспечивающих общий высокий уровень метаболизма и возможность его активной регуляции. Действительно, еще до появления терморегуляции в организме идут процессы наращивания энергетических резервов в виде гликогена, увеличения количества эритроцитов, гемоглобина и сахара в крови, происходит интенсивный рост сердца. Все эти показатели достигают полного выражения к включению химической терморегуляции.
Появление физической терморегуляции в виде птиломоторных реакций на холод зависит от формирования оперения и у незреловылупляющихся птенцов начинает функционировать лишь к концу гнездового периода.
Выводковые (матуронатные) птенцы приобретают способность к терморегуляции практически сразу после выклева; сроки включения гомойотермных реакций колеблются в пределах от нескольких часов до 2 суток и более. Как и у имматуронатных, быстрый переход к гомойотермии у птенцов выводковых видов подготовлен постепенным повышением дыхательной функции крови и увеличением энергетических ресурсов организма.
Птицы, занимающие промежуточное положение между двумя крайними по степени матуронатности типами развития (например, чайки), характеризуются соответствующими сдвигами в сроках развития терморегуляции.
Источник



