
Способы соединения нервных клеток
Взаимодействия между глиальными и нервными клетками отчетливо проявляются в процессах развития и структурной организации нервных волокон. Нервным волокном называется отросток нервной клетки, окруженный глиальной оболочкой.
Непосредственно сам отросток называют еще осевым цилиндром, а клетки глиальной оболочки — нейролеммоцитами. Различают миелиновые (мякотные) и безмиелиновые (безмякотные) нервные волокна.
В безмиелиновых нервных волокнах отростки нервных клеток погружены в углубления на поверхности нейролеммоцитов, имеющих вид желоба. Погруженный в тело глиальной клетки нервный отросток ограничен как собственной плазмолеммой, так и внешней мембраной нейролеммоцита. Он как бы подвешен на двухлистковой ее складке. Эти складки мембран (своеобразные ультраструктурные «брыжейки») называют мезаксонами. Безмиелиновые волокна могут включать несколько осевых цилиндров.
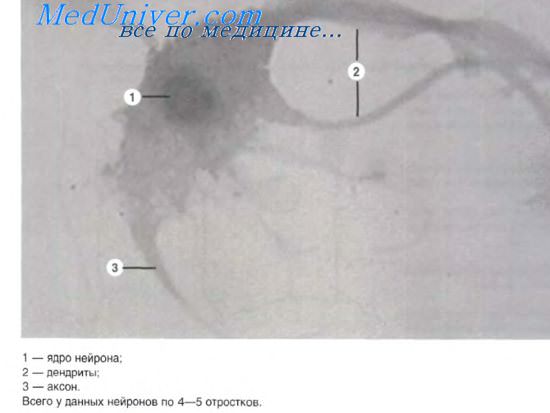
Миелиновое нервное волокно состоит из нервного отростка и нейролеммоцитов (шванновских клеток). Осевой цилиндр не просто погружен в цитоплазму нейролеммоцита, а окружен спиральной слоистой оболочкой (миелином), образованной наматыванием мезаксонов нейролеммоцитов при их вращении вокруг отростка нервной клетки. В миелиновой оболочке обнаружены липиды, щелочной белок миелина, маркерный белок S100 и др.
Высокое содержание липидов (почти 2/3 массы миелина) выявляется при обработке препаратов четырехокисью осмия, окрашивающей миелиновую оболочку в темно-коричневый цвет. По ходу миелинового волокна имеются сужения — узловые перехваты (перехваты Ранвье). Они соответствуют границе смежных нейролеммоцитов. Каждый межузловой сегмент оболочки волокна представлен одним нейролеммоцптом. Миелиновые волокна толще безмиелиновых. Скорость проведения нервного импульса по ним составляет 5-120 м/с, тогда как по безмиелиновым волокнам импульс проводится со скоростью 1-2 м/с.
Сложные взаимоотношения между нервными и глиальными клетками складываются при формировании чувствительных нервных окончаний (рецепторов) и двигательных нервных окончаний (эффекторов).
Нервные окончания — концевой аппарат нервных волокон, формирует межнейрональные контакты, или синапсы, рецепторные (чувствительные) окончания и двигательные (эффекторные) окончания.
Синапс (от synapsis — соединение) — специализированный для передачи нервных импульсов контакт между двумя нейронами или между нейроном и эффектором. Процессы возбуждения нейронов, возникновение импульсов и распространение их по отросткам связаны с изменениями в плазмолемме. Она является структурной основой возникновения и передачи потенциалов действия. Плазмолемма имеет существенные особенности строения и функции в участках, входящих в состав синапсов.
Межнейрональные синапсы бывают нескольких видов: аксосоматические (между аксоном одного нейрона и телом другого нейрона); аксодендритические (между аксоном одного нейрона и дендритом другого нейрона); аксоаксональные (между аксонами двух нейронов). Описаны также синапсы соматосоматические, дендродендритические и др.
Все синапсы по механизму передачи импульсов между нервными клетками подразделяются на 3 типа: синапсы с химической передачей, электротонические и смешанные синапсы. Типичный синапс с химической передачей состоит из пресинаптической и постсинаптической частей, а также синаптической щели. Пресинаптическая часть включает концевое расширение аксона, ограниченное пресинаптической мембраной. Специфическими структурами этой части являются синоптические пузырьки, содержащие нейромедиаторы. Пузырьки бывают со светлым и электронно-плотным содержимым и называются в связи с этим агранулярными и гранулярными.
По форме они подразделяются на круглые и уплощенные. На внутренней поверхности пресинаптической мембраны расположены конусовидные электронно-плотные образования — пресинаптические уплотнения. В цитоплазме пресинаптической части имеются митохондрии. Синаптическая щель размером 20-30 нм содержит филаменты, связывающие наружные слои плазмолеммы контактирующих нейронов.
Постсинаптическая часть в составе плазмолеммы второго нейрона имеет рецепторы к медиатору, который выделяется в синаптическую щель при деполяризации мембраны первого нейрона. Внутренняя поверхность постсинаптической мембраны характеризуется наличием электронно-плотного слоя цитоплазмы — постсинаптические уплотнения.
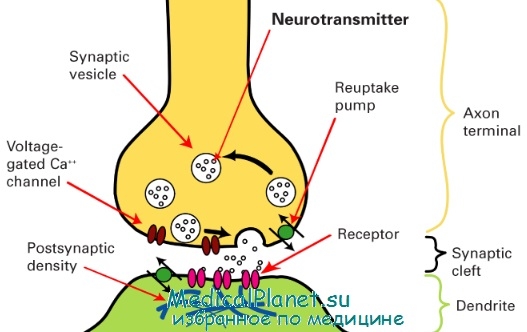 Схема строения синапса
Схема строения синапса
Источник
ВВЕДЕНИЕ В КОГНИТИВНЫЕ НЕЙРОНАУКИ. Глава 3. Нейроны и связи между ними.

Глава 3. Нейроны и связи между ними.
Что мы знаем о процессах, происходящих на уровне нейронов? Можем ли мы сейчас построить непротиворечивую теорию относительно событий на этом уровне?
Основными клетками мозга являются нейроны, высококонсервативные с эволюционной точки зрения. Они сохранялись в относительно неизменном виде в течение многих сотен миллионов лет, и даже очень разные виды животных имеют одинаковые типы нейронов. Со многих точек зрения нейроны не отличаются от остальных клеток, но есть то, что выделяет их среди остальных: специализация на электрохимической сигнализации, благодаря которой они способны принимать входящий сигнал на дендритах и посылать электрохимический сигнал вдоль аксона. Весь мозг можно рассматривать как сверхсложную структуру, состоящую из связанных между собой нейронов.
Дендритами и аксонами называют выросты тела нейрона; один нейрон может иметь до десяти тысяч дендритов и один аксон.
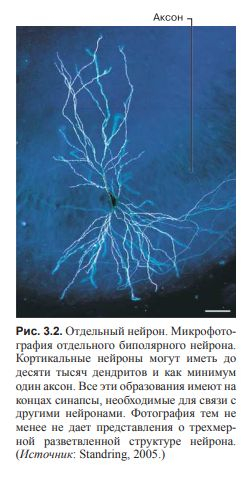
(рис. 3.2 и 3.3). 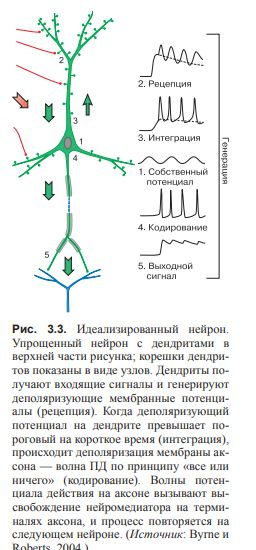
Потенциал действия (ПД) проходит по аксону значительно медленнее, чем электрический ток в компьютере, однако многие задачи наш мозг выполняет гораздо лучше современных компьютеров. В настоящее время компьютеры далеко отстоят от человека в задачах восприятия, языковой коммуникации, семантической памяти, контроля движения и творчества.
Нейробиология концентрирует внимание на связи и взаимодействии нейронов. Рассмотрение таких связей удобно начинать как раз с генерализованного нейрона.
Классические нейроны соединяются при помощи синапсов, которые могут быть возбуждающими и тормозящими.
Активность нейрона опосредована десятками факторов — циклом сна и бодрствования, доступностью предшественников нейромедиаторов и многими другими. Все эти факторы влияют на вероятность прохождения сигнала между двумя нейронами и могут быть представлены в виде синаптических весов. Таким образом, все разнообразие нейронов можно с успехом представить в виде интегративного нейрона, а все способы межнейронной коммуникации — в форме вероятности прохождения сигнала между нейронами.
Существует по меньшей мере шесть основных нейромедиаторов и не менее тридцати «менее важных», в основном являющихся нейропептидами.
Даже дендриты отдельной клетки, по всей видимости, способны к обработке информации. Имеются также данные, что способна принимать участие в обработке информации и нейроглия — поддерживающая ткань нервной системы.
На настоящий момент известно о существовании в некоторых частях взрослого головного мозга стволовых клеток. Образование новых синапсов идет в течение всей жизни; для образования новых синапсов отростки дендритов способны образовываться за несколько минут.
1.3. Обработка информации нейронами.
Искусственные нейронные сети использовались для моделирования многих функций мозга — распознавания элементов изображений, управления роботами, обучения и улучшения функционирования на основе опыта.
Во многих случаях такие сети выполняли задачи лучше, чем компьютерные программы, основанные на логике и математике.
Так же, они помогают нам понять принципы работы реальных нейронных сетей в мозге.
Нейросети помогают нам понять работу нервной системы.
Так искусственные нейросети могут служить моделями для изучения реальных структур в мозге.
Мы ограничимся рассмотрением синапсов только двух типов — возбуждающего (повышающего вероятность прохождения ПД (Потенциал действия) на постсинаптическом нейроне) и тормозящего (понижающего такую вероятность).
Глутамат — наиболее распространенный медиатор в ЦНС — является возбуждающим.
ГАМК (гаммаАминоМасляная кислота) является наиболее распространенным тормозным медиатором.
В нервной системе распространены массивы нейронов, часто именуемые картами.
2.1. Упрощенный случай: рецепторы, пути и контуры.
Каждый сенсорный нерв может содержать несколько параллельных каналов, каждый из которых проводит несколько различающуюся информацию. Так, зрительный тракт имеет канал передачи цвета, называемый мелкоклеточным, и канал передачи формы и размеров объекта, называемый крупноклеточным.
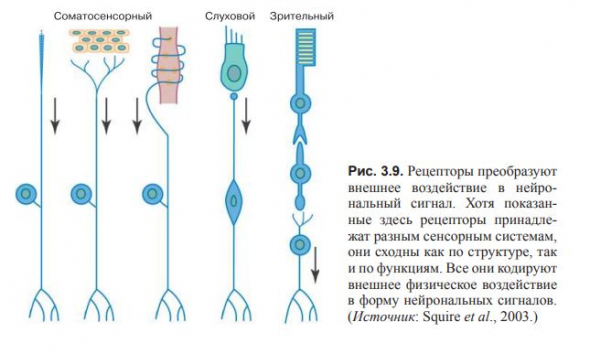
Точно так же соматосенсорные пути сочетают каналы передачи прикосновения, давления, боли и некоторые другие.
Большинство сенсорных волокон оканчиваются в таламусе, где они передают сигнал нейронам, оканчивающимся в коре.
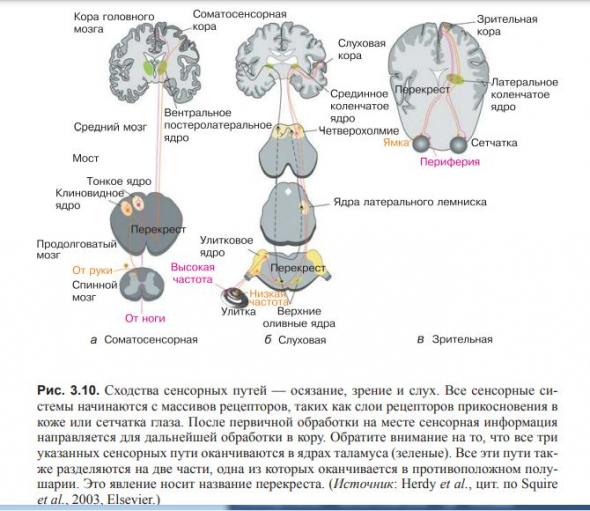
(рис. 3.10 и 3.11) 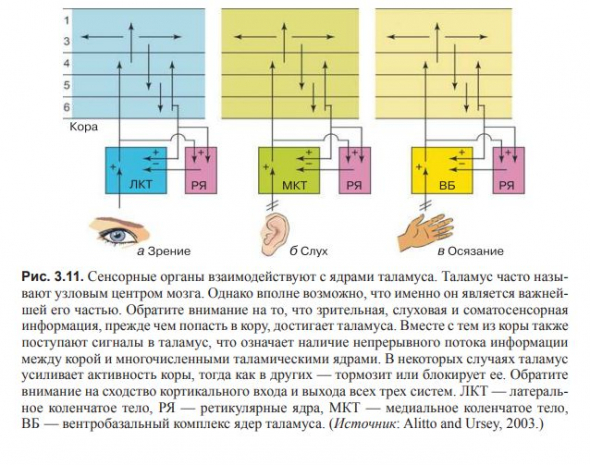
Таким образом, в большинстве сигнальных путей существуют петли обратной связи — такие, как в нейросети с двумя или более слоями.
С этой точки зрения мозг представляется системой воздействующих друг на друга массивов и сетей.
Массивы нейронов представляет собой двумерную сеть нейронов.
Когда массивы соответствуют пространственной организации той или иной структуры, их называют картами.
В мозге имеет место как временное, так и пространственное кодирование, наряду со многими другими способами кодирования и обработки информации.
Пространственные карты являются наиболее наглядной формой пространственного кодирования.
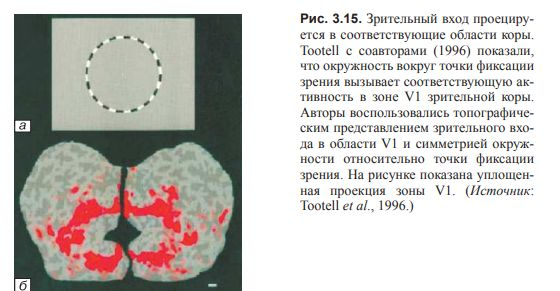
(рис. 3.15 и 3.16). 
Такая соматосенсорная информация , как информация о прикосновении и боли, также обрабатывается кортикальными картами. Другие чувства, такие как слух, вкус и обоняние, гораздо меньше ассоциированы с положением рецепторов, однако в слуховой коре имеется картоподобный клиновидный регион.
Таким образом, даже информация от не ассоциированных с пространством органов чувств обрабатывается массивами и картами нейронов.
Наш мозг организует огромные количества входящей информации так, чтобы отражать положение окружающих объектов. Моторная кора, как вы можете догадаться, также выглядит как непропорциональная карта скелетных мышц тела.
Главным вопросом относительно сенсорики на сегодняшний день яляется вопрос о том, как осуществляется высокоуровневая обработка воспринятой информации. И модель нейросетей предоставляет один из возможных ответов.
Мозг постоянно корректирует работу моторных систем на основании сенсорной информации и адаптирует сенсорные системы при помощи моторной активности.
Сенсорные системы можно представить в виде иерархических систем, состоящих из иерархических систем низшего порядка, начиная с рецепторов и постепенно переходя ко все более сложным объектам.
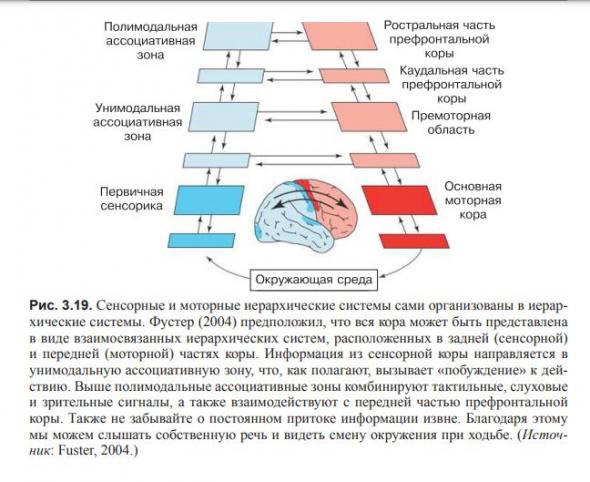
Моторные системы могут быть представлены в виде иерархической структуры противоположного направления, заканчивающиеся на мотонейронах .
Идет непрерывный обмен информацией между двумя системами в процессе цикла от восприятия до действия, начиная с наинизшего и заканчивая высшими уровнями планирования, мышления и анализа возможного развития событий

(рис. 3.20 Иерархическая система из области архитектуры).
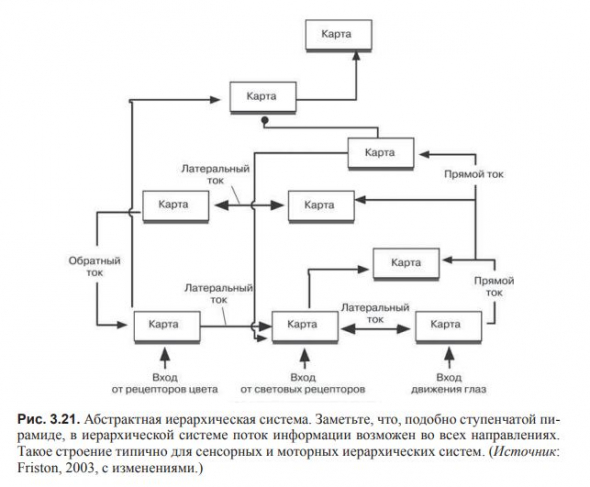
В схеме иерархической системы обработки информации, каждый массив нейронов назван картой; карты существуют на разных уровнях, и сигнал может идти вверх, вниз и к другой карте того же уровня.
При рассмотрении электрической активности десятков миллиардов нейронов мозг поневоле начинает казаться огромным оркестром, а не одним инструментом. За сотни миллионов лет эволюции в мозге появились нейроны с самыми разными видами временного и пространственного кодирования (блок 3.1).
В путях следования информации имеется множество точек выбора, с которых она может быть направлена по нескольким различным путям или быть передана на уровень выше или ниже.
Если вернуться к ступенчатой пирамиде, то такой разветвленный путь похож на путь человека к вершине: он может достигнуть ее прямым или окружным путем.
Зрительная картина мира подвержена постоянным изменениям. Однако мозг тем не менее ведет обработку таких изменений. Животное не может позволить себе не заметить хищника, прячущегося в траве, только потому, что сейчас закат, или потому, что на него падает тень.
Для того чтобы выжить, мы должны были иметь превосходную зрительную систему.
Вот, например, кошка, выслеживающая жертву, может осматривать дерево только одним глазом, тогда как другой глаз остается пассивным. Это приводит к явлению бинокулярной конкуренции — конкуренции между зрительными входами от разных глаз.
Многие животные получают от разных глаз совершенно разные входы — такие животные, как кролики и олени, вообще не имеют участков перекрытия полей зрения, поэтому для них явление бинокулярной конкуренции невозможно.
Мозг постоянно имеет некоторые ожидания относительно встречаемых им внешних условий. Спускаясь по лестнице в темноте, мы ожидаем, что под ногой будет ступенька.
При анализе неоднозначно трактуемых объектов ожидание обусловливает выбор наиболее приемлемого варианта трактовки. Многие слова в языке имеют больше одного значения, поэтому, даже читая это, вы вынуждены иметь дело с неоднозначностями. Мозг опирается не только на входящую информацию — он имеет множество причин выбора того или иного варианта, основанных на предсказании результата и ожидании.
Селективное внимание позволяет нам динамически изменять свои сенсорные предпочтения, а долговременная память увеличивает силу синапсов, ответственных за точное восприятие.
Многие ученые считают, что всю кору целиком, наряду с сопутствующими областями, такими как таламус, следует рассматривать как одну функциональную единицу. Ее часто называют таламокортикальной системой.
Одним из основных свойств поведения животных является способность адаптироваться.
Основным свойством мозга, таким образом, является приспособляемость. Однако какие изменения в структуре самого мозга приводят к такой приспособляемости?
Для этих целей гораздо лучше подходят методы визуализации структур мозга, получившие интенсивное развитие в два прошедших десятилетия.
Хотя большая часть методов визуализации ориентирована на конкретную область, подчеркивая тем самым функциональное разделение, а не интеграцию, были предприняты попытки изучения обучения как системного процесса, включающего глобальные изменения структуры и функций мозга.
Развитие технологии магнитно-резонансной томографии позволило начать изучение изменений структурных компонентов связи — трактов белого вещества — под влиянием обучения.
Как было показано, обучение жонглированию вызывает изменения как в сером, так и в белом веществе мозга.
Данные результаты стали поистине революционными, поскольку долгие годы считалось, что структура мозга неизменна.
Подобные открытия, позволяющие рассматривать мозг как функционально и структурно лабильный орган, вне всякого сомнения являются шагом вперед в нашем понимании процесса обучения.
(4.0. Адаптация и обучение массивов нейронов).
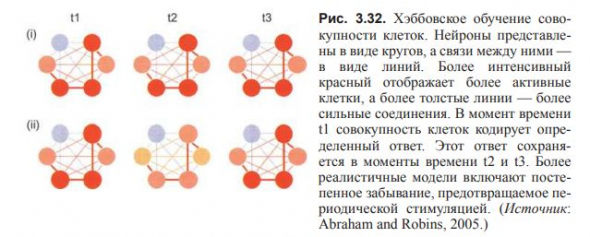
Наиболее известное правило обучения нейросетей, выражаемое в лозунге «neurons that fire together, wire together».
Нейроны, которые срабатывают вместе, соединяются вместе.
(Обучение по Хэббу).
Дональд Хэбб в 1949 г. постулировал, что ассамблеи нейронов способны обучаться благодаря усилению связей между нейронами, активирующимися при стимуляции одновременно.

В основе обучения и памяти лежит эффективность синаптической связи.
Существует множество способов воздействия на эффективность синаптической передачи. Так, два нейрона могут образовать больше синапсов, в самих синапсах может вырабатываться больше нейромедиатора, рецепторы постсинаптического нейрона могут стать эффективнее.
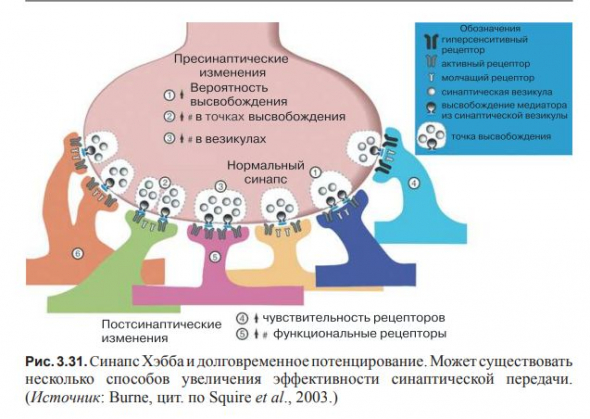
В обучении задействовано два типа изменений; их можно рассматривать как усиленное возбуждение и усиленное торможение.
Долговременное усиление возбудимости одного нейрона называют долговременой потенциацией.
Долговременное понижение же — долговременной депрессией. Оба события имеют место в гиппокампе.
Визуально обучение по Хэббу можно представить в виде утолщения линий между узлами сети, как в простой совокупности клеток.
Модели с третьим, скрытым слоем позволяют нейросети изменять силу соединений.
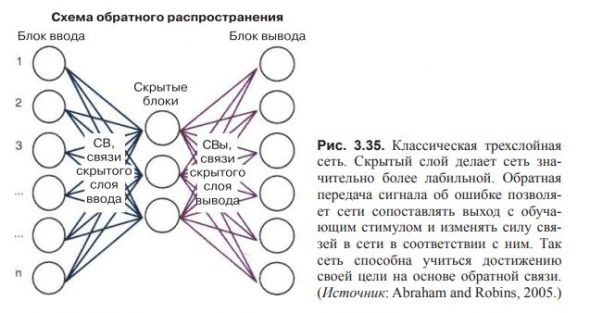
Классическая трехслойная прямая сеть со скрытым слоем и настраиваемой силой взаимодействий может эффективно обучаться путем сопоставления выхода нейросети с желаемым выходом и подстройки силы соединений для достижения желаемого результата.
Процесс носит название обратного распространения ошибки обучения и во многом подобен отрицательной обратной связи.
Сети такого типа на сегодняшний день наиболее распространены.
В самоорганизующейся аутоассоциативной сети выход ставится в соответствие входу.
Такая стратегия полезна при распознавании паттернов, таких как звук знакомого голоса.
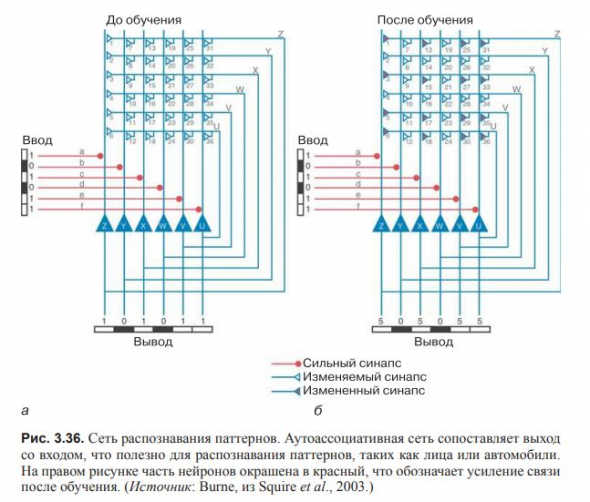
Самоорганизующиеся системы используются в природе для решения многих задач.
Сами организмы и их нервные системы можно рассматривать как самоорганизующиеся системы.
Самоорганизующаяся сеть способна справляться с фундаментальой проблемой распознавания человеческих лиц.
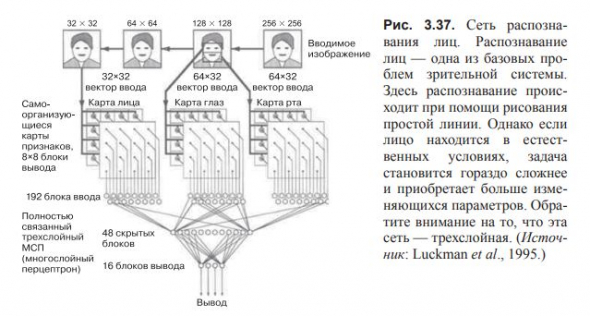
Человек учится реагировать на нормальные, недеформированные лица в очень ранний период жизни и вскоре становится способен отличать знакомые лица от незнакомых.
Задача, решаемая сетью, гораздо проще решаемой человеком, поскольку в модели происходит только формирование цепи.
Сеть способна обучаться предугадывать расположение рта в нижней части рисунка и двух глаз — в верхней.
4.2. Дарвинистский подход в нервной системе: выживают клетки и синапсы, наиболее приспособленные к данной задаче.
Нейральный дарвинизм предполагает, что нейроны развиваются и соединяются друг с другом в соответствии с дарвинистскими принципами.
Селекционизм — эффективный способ адаптации.
Отбор нейронов приводит к образованию долгоживущих нейронных совокупностей, выполняющих задачи адаптации, обучения, разпознавания паттернов и им подобные.
Нейросети отличаются высоким уровнем параллельности (что означает способность производить много разных вычислений одновременно) и распределенности (способности обрабатывать информацию в разных местах с использованием разных механизмов).
Это говорит о большей близости нейросетей к биологическим способам обработки информации.
Нейронные сети довольно просто перевести на язык математических выражений.
Нейросети способны обрабатывать символьную информацию, а символы могут быть переведены в нейросети.
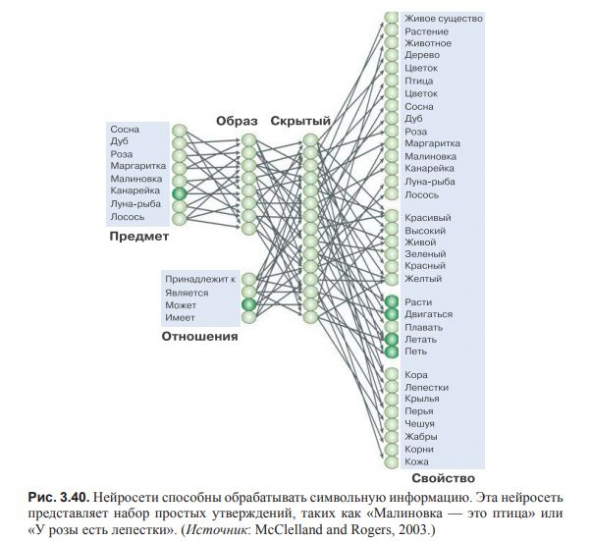
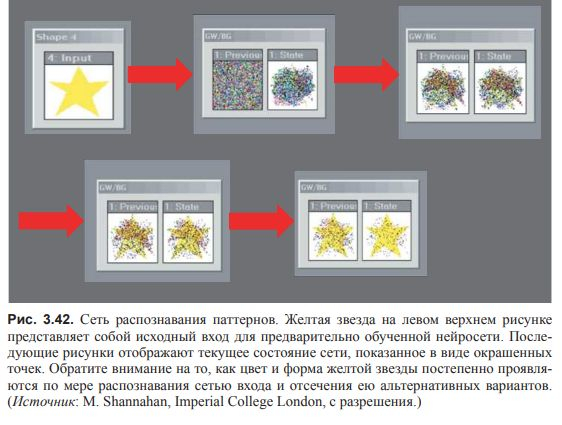
Обучение нейросети проявляются по мере распознавания сетью входа и отсечения ею альтернативных вариантов.
Существует масса способов координации работы нейронов. Одним из них являются масштабные ритмы, координирующие работу больших групп нейронов так же, как дирижер координирует игру симфонического оркестра. Если большая масса нейронов активируется одновременно, то их активность, как правило, суммируется.
Современные данные говорят в пользу гораздо более быстрых гамма- и тета-корреляций на тех частотах, на которых мозг выполняет большую часть этой работы.
Ритмы энцефалограммы на сегодняшний день считаются сигнализирующими о разных, но скоординированных процессах.
К примеру, гамма-ритмы высокой плотности считаются ассоциированными с осознанным зрительным восприятием и процессом решения простой проблемы эквивалентности.
Альфа-ритмы традиционно ассоциируются с отсутствием задач, требующих фокусировки внимания, тогда как тета-ритмы, как полагают на настоящий момент, контролируют гиппокампальную область и фронтальную кору в процессе обращения к долговременной памяти. Дельта-ритмы — сигналы глубокого сна — группируют быструю нейрональную активность с целью консолидации полученных данных.
При проектировании самолета инженеры закладывают в его конструкцию некоторую функциональную избыточность на случай выхода из строя важнейших систем. Так, если откажет один двигатель, то большая часть самолетов будет способна дотянуть до взлетно-посадочной полосы на оставшихся.
Человек и животные также обладают определенной функциональной избыточностью.
Мозга это правило тоже касается. Мозг способен работать даже после получения весьма значительных повреждений.
Латеральное торможение является распространенной стратегией для выделения различий между двумя однородными областями сигнала, такими как темные пятна на светлом фоне.
Клетки сенсорных систем имеют так называемые рецептивные поля, настроенные на определенные параметры входа, такие как ориентация линии, цвет, движение, форма и тип объекта. При повышении уровня визуальных карт их разрешение падает, в то время как способность к интеграции информации растет.
Поскольку сенсорные и моторные системы изучают отдельно друг от друга, мозг представляется нам огромным сенсомоторным органом, делающим возможным непрерывные высокоуровневые взаимодействия между входом и выходом.
Пространственные массивы нейронов делают возможным пространственное кодирование, однако не стоит забывать о том, что в нервной системе имеется еще и кодирование временное. Основные ритмы энцефалограммы, как полагают, отвечают за временную координацию активности больших групп нейронов.
Последние исследования позволяют предположить, что гамма-ритм ответственен за интеграцию сенсорной информации осознанные ощущения, а тета-ритм — за извлечение информации из долговременной памяти.
Контрольные задания к этой главе.
1. Опишите основные функции интегративного нейрона.
2. Что такое латеральное торможение и какую роль оно играет в сенсорных системах?
3. Каким образом сенсорные и моторные системы можно рассматривать в форме иерархических структур?
4. Опишите роль двусторонних взаимодействий в функционировании мозга.
5. Что такое дарвинистский подход к нервной системе и какие аспекты процессов, происходящих в мозге, он затрагивает?
6. Назовите три наиболее общих свойства сенсорных систем.
Источник



