
- Триацилглицеролы нужны при работе
- Общая характеристика мобилизации ТАГ
- Медицинская биохимия, принципы измерительных технологий в биохимии, патохимия, диагностика, биохимия злокачественного роста. Часть 2.
- Метаболизм липидов в норме и при патологии. Дисфункция жировой ткани. Ожирение
- 1. Метаболизм липидов
- Синтез фосфолипидов и триацилглицеролов тесно связаны
- Синтез ТАГ — это запасание энергии
- Липогенез
- Хиломикроны и ЛПОНП переносят триацилглицеролы
- Характеристика хиломикронов
- Характеристика липопротеинов очень низкой плотности
Триацилглицеролы нужны при работе
Синтезируясь во время и сразу после приема пищи (липогенез) и запасаясь в жировой ткани, триацилглицеролы являются формой хранения насыщенных и мононенасыщенных жирных кислот. Распад триацилглицеролов (триглицеридов) по-другому называется липолиз или мобилизация жира. Он идет в жировых клетках постоянно и обычно существует равновесие между синтезом и распадом ТАГ.
Даже в состоянии покоя организма печень, сердце, скелетные мышцы и другие ткани (кроме эритроцитов и нейроцитов) более 50% энергии получают из окисления жирных кислот, поступающих из жировой ткани благодаря фоновому липолизу. По мере уменьшения резервов глюкозы клетки все больше энергии получают из окисления жирных кислот. Таким образом, насыщенные жирные кислоты выполняют роль своеобразного буфера энергии в организме.
Мобилизация триацилглицеролов и окисление жирных кислот активируется
- при нормальных физиологических стрессовых ситуациях – эмоциональный стресс, мышечная работа, голодание,
- при патологических состояниях – сахарный диабет I типа, другие гормональные заболевания (гиперкортицизм, гипертиреоз).
В результате стимулированного липолиза в адипоцитах образуются свободный глицерол и жирные кислоты .
Глицерол с кровью доставляется в печень и почки, здесь фосфорилируется и окисляется в метаболит гликолиза диоксиацетонфосфат. В зависимости от условий диоксиацетонфосфат может включаться в реакции глюконеогенеза (при голодании, мышечной нагрузке) или окисляться в реакциях гликолиза.
Жирные кислоты транспортируются в крови в комплексе с альбуминами плазмы:
- при физической нагрузке – в мышцы ,
- в обычных условиях и при голодании – в мышцы и большинство тканей , однако при этом около 30% жирных кислот захватывается печенью .
При голодании и физической нагрузке после проникновения в клетки жирные кислоты вступают на путь β-окисления .
Общая характеристика мобилизации ТАГ
В целом мобилизацию жира можно представить как последовательность следующих событий:
- Липолиз – гормонзависимый распад ТАГ в жировой ткани или резервных ТАГ в самой клетке.
- Транспорт жирных кислот из жировой ткани по крови в комплексе с альбумином.
- Проникновение жирной кислоты в цитозоль клетки-мишени.
- Активация жирной кислоты через присоединение HS-КоА.
- Карнитин-зависимое перемещение жирной кислоты в митохондрию.
- Окисление жирной кислоты с образованием ацетильных групп (в форме ацетил-SКоА).
- Сгорание ацетил-SКоА в цикле лимонной кислоты или синтез (только в печени) кетоновых тел.
Источник
Медицинская биохимия, принципы измерительных технологий в биохимии, патохимия, диагностика, биохимия злокачественного роста. Часть 2.
Метаболизм липидов в норме и при патологии. Дисфункция жировой ткани. Ожирение
%2C_de_Juan_Carre%C3%B1o_de_Miranda..jpg)
В 1680 году Juan Carreno de Miranda была изображена девочка-«монстр», страдающая синдромом Prader–Willi (1954)
1. Метаболизм липидов
Термин «липиды» объединяет вещества, обладающие общим физическим свойством — гидрофобностью, то есть нерастворимостью в воде. Однако такое определение в настоящее время является не совсем корректным ввиду, того, что некоторые группы (триацилглицерины, фосфолипиды, сфинголипиды и др.) проявляют себя как амфифильные или дифильные соединения, т.е. способные растворяться как в полярных веществах (гидрофильность), так и в неполярных (гидрофобность). По структуре липиды настолько разнообразны, что у них отсутствует общий признак химического строения. Липиды разделяют на классы, в которые объединяют молекулы, имеющие сходное химическое строение и общие биологические свойства.
Основную массу липидов в организме составляют жиры — триацилглицеролы, служащие формой депонирования энергии. Жиры располагаются преимущественно в подкожной жировой ткани и выполняют также функции теплоизоляционной и механической защиты. Триацилглицеролы практически не растворимы в воде, поэтому их депонирование не сопровождается появлением осмотических проблем.
Фосфолипиды — большой класс липидов, получивший своё название из-за остатка фосфорной кислоты, придающего им свойства амфифильности. Благодаря этому свойству фосфолипиды формируют бислойную структуру мембран, в которую погружены белки. Клетки или отделы клеток, окружённые мембранами, отличаются по составу и набору молекул от окружающей среды, поэтому химические процессы в клетке разделены и ориентированы в пространстве, что необходимо для регуляции метаболизма.
Стероиды, представленные в животном мире холестеролом и его производными, выполняют разнообразные функции. Холестерол — важный компонент мембран и регулятор свойств гидрофобного слоя. Производные холестерола (жёлчные кислоты) необходимы для переваривания жиров. Стероидные гормоны, синтезируемые из холестерола, участвуют в регуляции энергетического, водно-солевого обменов, половых функций. Кроме стероидных гормонов, многие производные липидов выполняют регуляторные функции и действуют, как и гормоны, в очень низких концентрациях. Например, тромбоцитактивирующий фактор — фосфолипид особой структуры — оказывает сильное влияние на агрегацию тромбоцитов в концентрации 10 -12 М; эйкозаноиды, производные полиеновых жирных кислот, вырабатываемые почти всеми типами клеток, вызывают разнообразные биологические эффекты в концентрациях не более 10 -9 М. Из приведённых примеров следует, что липиды обладают широким спектром биологических функций.
В тканях человека количество разных классов липидов существенно различается. В жировой ткани жиры составляют до 75 % сухого веса. В нервной ткани липидов содержится до 50 % сухого веса, основные из них фосфолипиды и сфингомиелины (30 %), холестерол (10 %), ганглиозиды и цереброзиды (7 %). В печени общее количество липидов в норме не превышает 10-13 %.
Нарушения обмена липидов приводят к развитию многих заболеваний, но среди людей наиболее распространены два из них — ожирение и атеросклероз.
Липидный обмен включает в себя следующие процессы:
- Расщепление, переваривание и всасывание липидов в пищеварительном тракте, поступающих вместе с пищей.
- Транспорт жиров из кишечника с помощью хиломикронов.
- Обмен триацилглицеролов.
- Обмен фосфолипидов.
- Обмен холестерола.
- Взаимопревращения жирных кислот и кетоновых тел.
- Липогенез.
- Катаболизм липидов — липолиз.
- Катаболизм жирных кислот.
Печень является главным местом синтеза жирных кислот, жиров, кетоновых тел и холестерина. Жиры могут также синтезироваться в жировой ткани, однако её основной функцией остаётся депонирование липидов.
Источник
Синтез фосфолипидов и триацилглицеролов тесно связаны
Начальные реакции синтеза триацилглицеролов и фосфолипидов совпадают и происходят при наличии глицерола и жирных кислот .
В реакциях биосинтеза можно выделить следующие события:
1. Образование глицерол-3-фосфата через диоксиацетонфосфат из глюкозы или при фосфорилировании свободного глицерола.
2.. Биосинтез фосфатидной кислоты – требует наличия глицерол-3-фосфата и жирных кислот. При связывании глицерол-3-фосфата с жирными кислотами синтезируется фосфатидная кислота.
Далее фосфатидная кислота может превращаться двумя путями – в ЦДФ-ДАГ или дефосфорилироваться до 1,2-ДАГ.
3. Синтез триацилглицерола (липогенез) – идет из 1,2-ДАГ после дефосфорилирования фосфатидной кислоты. Образованный 1,2-ДАГ ацилируется до ТАГ.
4. Синтез фосфолипидов . Сейчас рассматриваются два пути синтеза фосфолипидов.
- По одному пути 1,2-ДАГ не превращается в ТАГ, а связывается с этаноламином с образованием фосфатидилэтаноламина, либо с холином – образуется фосфатидилхолин.
- По другому пути, ЦДФ-ДАГ связывается либо с инозитолом, либо с серином с образованием соответственно фосфатидилинозитола или фосфатидилсерина. При декарбоксилировании фосфатидилсерина далее образуется фосфатидилэтаноламин, который может превратиться, в свою очередь, в фосфатидилхолин.
Синтезированный любым способом фосфатидилэтаноламин также способен взаимодействовать с серином и обратно образовывать фосфатидилсерин.
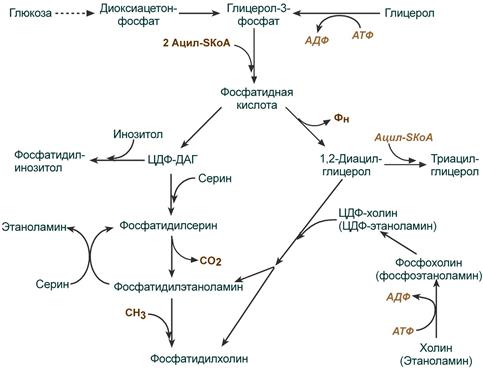
Общая схема реакций синтеза триацилглицеролов и фосфолипидов
ЦДФ-ДАГ, являясь активной формой фосфатидной кислоты, способен превращаться не только в фосфатидилинозитол, фосфатидилсерин, но и в другие фосфолипиды, например в кардиолипин .
Источник
Синтез ТАГ — это запасание энергии
Липогенез
Синтез триацилглицеролов (липогенез) заключается в дефосфорилировании фосфатидной кислоты, полученной из глицерол-3-фосфата, и присоединении ацильной группы.
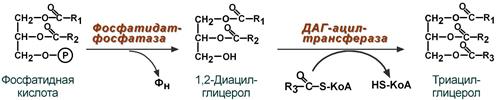
Реакции синтеза ТАГ из фосфатидной кислоты
Если синтез ТАГ происходил в печени, то они эвакуируются из нее в ткани, имеющие на эндотелии своих капилляров липопротеинлипазу (транспорт ТАГ в крови). Транспортной формой служат ЛПОНП. Строго говоря, клеткам организма нужны только жирные кислоты, все остальные компоненты ЛПОНП не являются необходимыми.
Синтез ТАГ увеличивается при соблюдении хотя бы одного из следующих условий, которые обеспечивают появление избытка ацетил-SКоА :
- наличие источника «дешевой» энергии . Например,
1) диета богатая простыми углеводами (глюкоза, сахароза). При этом концентрация глюкозы в печени и адипоцитах после еды резко повышается, она окисляется до ацетил-SКоА и под влиянием инсулина в этих органах активно происходит синтез жиров.
2) наличие этанола, высокоэнергетичного соединения, который окисляется до ацетил-SКоА. «Алкогольный» ацетил используется в печени для синтеза жира при условии нормального питания. Примером может служить «пивное ожирение». - повышение концентрации жирных кислот в крови . Например, при усиленном липолизе в жировых клетках под воздействием каких-либо веществ (фармпрепараты, кофеин и т.п.), при эмоциональном стрессе и отсутствии (!) мышечной активности увеличивается поток жирных кислот в гепатоциты. Здесь в результате происходит интенсивный синтез ТАГ.
- высокие концентрации инсулина и низкие концентрации глюкагона – после приема высокоуглеводной и жирной пищи.
Источник
Хиломикроны и ЛПОНП переносят триацилглицеролы
Транспорт триацилглицеролов от кишечника к тканям ( экзогенные ТАГ) осуществляется в виде хиломикронов (ХМ), от печени к тканям ( эндогенные ТАГ) – в виде липопротеинов очень низкой плотности .
В транспорте ТАГ к тканям можно перечислить последовательность следующих событий:
- Образование незрелых первичных ХМ в кишечнике.
- Движение первичных ХМ через лимфатические протоки в кровь.
- Созревание ХМ в плазме крови – получение белков апоС-II и апоЕ от ЛПВП.
- Взаимодействие ХМ с липопротеинлипазой (ЛПЛ) эндотелия кровеносных сосудов, которая отщепляет жирные кислоты от ТАГ. Далее жирные кислоты проникают непосредственно в клетки данной ткани или, связываясь с альбумином, разносятся по организму. В результате количество ТАГ в хиломикроне резко снижается, образуются остаточные ХМ.
- Переход остаточных ХМ в гепатоциты и полный распад их структуры.
- Синтез ТАГ в печени из пищевой глюкозы. Использование липидов, пришедших в составе остаточных ХМ.
- Образование первичных ЛПОНП в печени.
- Созревание ЛПОНП в плазме крови – получение белков апоС-II и апоЕ от ЛПВП.
- Взаимодействие с липопротеинлипазой эндотелия и потеря большей части ТАГ. Образование остаточных ЛПОНП (по-другому липопротеины промежуточной плотности, ЛППП).
- Далее остаточные ЛПОНП
- переходят в гепатоциты и полностью распадаются,
- либо остаются в плазме крови и превращаются в ЛПНП.
![]()
Схема транспорта экзогенных и эндогенных триацилглицеролов
(цифрами обозначены события соответственно тексту)
Характеристика хиломикронов
Общая характеристика
- формируются в кишечнике из ресинтезированных жиров,
- яявляются самыми крупными липопротеинами, их размер от 100 до 1200 нм (0,1-1,2 мкм),
- в их составе преобладают ТАГ, мало белка, фосфолипидов и холестерола (2% белка, 87% ТАГ, 2% ХС, 5% эфиров ХС, 4% фосфолипидов),
- основным апобелком является апоВ-48, это структурный липопротеин, в плазме крови получают от ЛПВП белки апоС-II и апоЕ,
- в норме натощак не обнаруживаются, в крови появляются после приема пищи, поступая из лимфы через грудной лимфатический проток, и полностью исчезают через 10-12 часов,
- не атерогенны.
Функция
Транспорт экзогенных ТАГ из кишечника в ткани, запасающие или использующие жирные кислоты, в основном в жировую ткань, миокард, скелетные мышцы и лактирующую молочную железу, в меньшей степени в легкие, костный мозг, почки, селезенку. На эндотелии капилляров этих тканей имеется фермент липопротеинлипаза .
Количество липопротеинлипазы в жировой ткани увеличивается при действии инсулина и прогестерона.
Активность фермента здесь также возрастает при регулярном потреблении алкоголя (в эксперименте ежедневно по две условные единицы спирта), и как побочный эффект происходит одновременное повышение в крови антиатерогенных ЛПВП.
При действии адреналина в жировой ткани активность липопротеинлипазы снижается. У мужчин относительное количество фермента в жировой ткани по сравнению с мышечной ниже, чем у женщин (влияние тестостерона).
Частые физические нагрузки, даже низкой интенсивности (умеренная ходьба), быстро повышают активность фермента в скелетных мышцах (влияние СТГ и адреналина) и сжигание жиров в мышцах. И, наоборот, даже относительно недолгая гиподинамия в разы снижает здесь его активность, что открывает путь к эффективному усвоению хиломикронов и ЛПОНП в жировой ткани и ускоряет ожирение.
Для поддержания высокой активности липопротеинлипазы в мышечной ткани намного важнее меньше сидеть, чем интенсивно тренироваться.
Метаболизм
1. После ресинтеза жиров в эпителиоцитах кишечника формируются первичные хиломикроны, имеющие только апоВ-48.
2. Из-за большого размера они не проникают напрямую в кровеносное русло и эвакуируются через лимфатическую систему, попадая в кровь через грудной лимфатический проток.
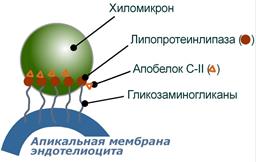
Взаимодействие хиломикрона и липопротеинлипазы
3. В крови хиломикроны взаимодействуют с ЛПВП и приобретают от них апоС-II и апоЕ, образуя зрелые формы. Белок апоС-II является активатором фермента липопротеинлипазы, белок апоЕ понадобится для удаления из крови остаточных хиломикронов.
4. На эндотелии капилляров вышеперечисленных тканей находится фермент липопротеинлипаза (ЛПЛ), отщепляющая жирные кислоты от ТАГ в положении 1 и 3, в результате накапливаются моно- и диацилглицеролы.
5. Находясь в плазме крови, хиломикроны также взаимодействуют с ЛПВП, отдавая им часть своих МАГ и ДАГ и получая взамен эфиры ХС.
6. После взаимодействия хиломикрона с липопротеинлипазой полученные свободные жирные кислоты проникают в клетки органа, либо остаются в плазме крови и в комплексе с альбумином разносятся с кровью в другие ткани. Липопротеинлипаза способна расщепиить до 90% всех ТАГ, находящихся в хиломикроне.
7. Остаточные (ремнантные) хиломикроны, сохранившие в своем составе МАГ и ДАГ, попадают в гепатоциты посредством апоЕ-рецепторного эндоцитоза и разрушаются до составных частей.
8. Часть фосфолипидной оболочки ремнантных хиломикронов, содержащая апоС-II и апоЕ, способна «отрываться» от основной частицы и формировать первичные ЛПВП.
Характеристика липопротеинов очень низкой плотности
Общая характеристика
Липопротеины очень низкой плотности:
- синтезируются в печени из эндогенных и экзогенных липидов,
- в их составе преобладают ТАГ, около 40% от массы составляют белок, фосфолипиды и холестерол (8% белка, 60% ТАГ, 6% ХС, 12% эфиров ХС, 14% фосфолипидов),
- основным белком является апоВ-100, выполняющий структурную функцию,
- в норме концентрация 1,3-2,0 г/л,
- слабо атерогенны.
Функция
Транспорт эндогенных и экзогенных ТАГ от печени в ткани, запасающие и использующие жирные кислоты, т.е. в те же ткани, что и хиломикроны.
Метаболизм
1. Первичные ЛПОНП образуются в печени и содержат только апоВ-100. Липидный компонент синтезируется
- из пищевых липидов, МАГ и ДАГ, пришедших в гепатоциты с остаточными хиломикронами,
- из липидов, синтезируемых из глюкозы.
2. В крови первичные ЛПОНП взаимодействуют с ЛПВП и приобретают от них апоС-II и апоЕ, образуя зрелые формы.
3. Аналогично хиломикронам, на эндотелии капилляров ряда тканей зрелые ЛПОНП подвергаются воздействию липопротеинлипазы с образованием свободных жирных кислот. Жирные кислоты перемещаются в клетки органа, либо остаются в плазме крови и в комплексе с альбумином разносятся с кровью в другие ткани.
4. При действии липопротеинлипазы в составе ЛПОНП снижается количество ТАГ и нарастает доля МАГ и ДАГ. Зрелый ЛПОНП превращается в остаточный (ремнантный) ЛПОНП.
5. Находясь в плазме крови, ЛПОНП взаимодействуют с ЛПВП, отдавая им часть своих МАГ и ДАГ и получая взамен эфиры ХС.
6. Остаточные ЛПОНП (также называемые липопротеины промежуточной плотности, ЛППП) в соотношении примерно 50 на 50
- либо попадают в гепатоциты посредством эндоцитоза, связанного со смешанным рецептором к апоЕ и апоВ-100-белкам,
- либо после воздействия на них печеночной липазы , находящейся в синусоидах печени, превращаются в следующий класс липопротеинов – липопротеины низкой плотности (ЛПНП).
Источник



