
Способы регуляции углеводного обмена



Регуляция углеводного обмена
А. Регуляция углеводного обмена
У высших организмов обмен углеводов подвержен сложным механизмам регуляции, в которых участвуют гормоны, метаболиты и коферменты. Представленная здесь схема относится к печени , которая занимает в углеводном метаболизме центральное место (см. с. 302). Некоторые из представленных механизмов не действуют в других тканях.
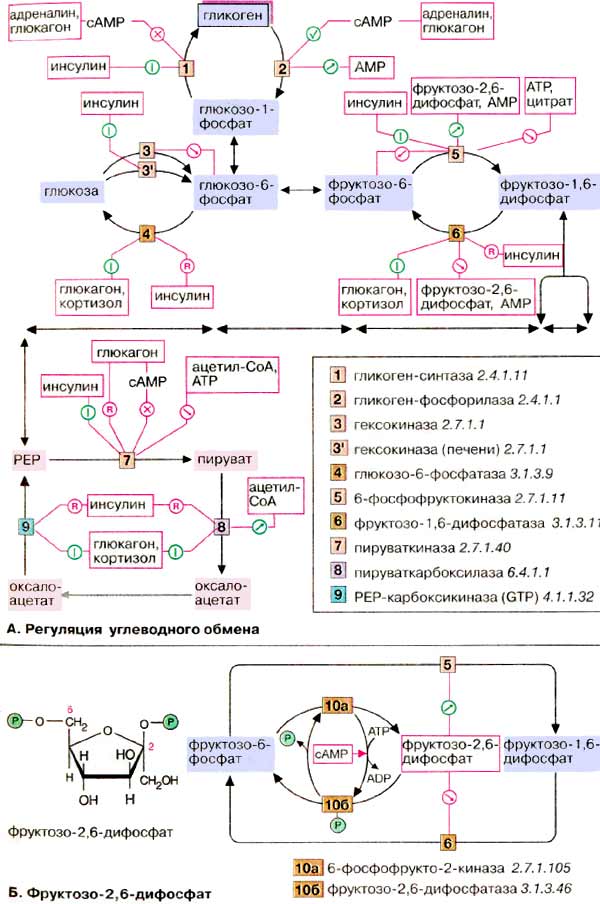
Одной из важнейших функций клеток печени является накопление избыточной глюкозы в виде гликогена и ее быстрое высвобождение по мере метаболической необходимости (буферная функция). После полной мобилизации запасов гликогена печень может поставлять глюкозу за счет синтеза de novo (глюконеогенез, см. сс. 156, 232). Кроме того, как и все ткани, она потребляет глюкозу путем гликолиза. Функции накопления (синтеза) глюкозы в виде гликогена и его распада должны быть взаимосогласованы. Таким образом, совершенно невозможно одновременное протекание гликолиза и глюконеогенеза, как и синтеза и деградации гликогена. Согласование процессов обеспечивается тем, что синтез (анаболизм) и распад (катаболизм) катализируются двумя различными ферментами и контролируются независимо. На схеме показаны только эти ключевые ферменты .
Гормоны. К гормонам, которые влияют на углеводный обмен, принадлежат пептиды инсулин и глюкагон , глюкокортикоид кортизол и катехоламин адреналин (см. сс. 362, 368). Инсулин индуцирует (см. с. 120) синтез de novo гликоген-синтазы [ 1 ], а также некоторых ферментов гликолиза [ 3, 5, 7 ]. Одновременно инсулин подавляет синтез ключевых ферментов глюконеогенеза (репрессия, [ 4, 6, 8, 9 ]). Глюкагон как антагонист инсулина действует в противоположном направлении: индуцирует ферменты глюконеогенеза [ 4, 6, 8, 9 ] и репрессирует пируваткиназу [ 7 ], ключевой фермент гликолиза. Другие аффекты глюкагона основаны на взаимопревращении ферментов и опосредованы вторичным мессенджером цАМФ (сАМР, см. с. 114). По этому механизму тормозится синтез гликогена [ 1 ] и активируется расщепление гликогена [ 2 ]. Подобным образом действует и адреналин. Торможение пируваткиназы [ 7 ] глюкагоном также обусловлено взаимопревращением ферментов.
Глюкокортикоиды, прежде всего кортизол (см. с. 362), индуцируют все ключевые ферменты глюконеогенеза [ 4, 6, 8, 9 ]. Одновременно они индуцируют ферменты деградации аминокислот и обеспечивают тем самым глюконеогенез исходными соединениями.
Метаболиты. Высокие концентрации АТФ (АТР) и цитрата тормозят гликолиз путем аллостерической регуляции фосфофруктокиназы. Кроме того, АТФ тормозит пируваткиназу. Ингибитором пируваткиназы является ацетил-КоА. Все эти метаболиты образуются при распаде глюкозы (торможение конечным продуктом). АМФ (AMP), сигнал дефицита АТФ, активирует расщепление гликогена и тормозит глюконеогенез.
Важную роль в обмене веществ в печени играет фруктозо-2,6-дифосфат . Это сигнальное вещество образуется в незначительных количествах из фруктозо-6-фосфата и выполняет чисто регуляторную функцию: стимулирует гликолиз путем активации фосфофруктокиназы и подавляет глюконеогенез с помощью торможения фруктозо-1,5-дифосфатазы.
Образование и распад фруктозо-2,6-дифосфата катализируются одним и тем же белком [ 10а и б ]. В нефосфорилированной форме этот белок вызывает образование фруктозо-2,6-дифосфата [ 10а ]. После фосфорилирования цАМФ-зависимой киназой он действует как фосфатаза [ 10б ] и катализирует превращение фруктозо-2,6-дифосфата в фруктозо-6-фосфат. В присутствии адреналина и глюкагона в клетках печени повышается уровень цАМФ (см. с. 122), т.е. оба гормона воздействуют как на гликолиз, так и на глюконеогенез. Суммарным результатом является быстрое повышение уровня глюкозы в крови.
Источник
Регуляция углеводного обмена
Регуляция углеводного обмена осуществляется на всех его этапах нервной системой и гормонами. Помимо этого, активность ферментов отдельных путей метаболизма углеводов регулируется по принципу “обратной связи”, в основе которого лежит аллостерический механизм взаимодействия фермента с эффектором. К аллостерическим эффекторам можно отнести конечные продукты реакции, субстраты, некоторые метаболиты, адениловые мононуклеотиды. Важнейшую роль в выборе направленности углеводного обмена (синтез или распад углеводов) играет соотношение коферментов НАД + /НАДН∙Н + и энергетический потенциал клетки.
Постоянство уровня глюкозы в крови – важнейшее условие поддержания нормальной жизнедеятельности организма. Нормогликемия является результатом слаженной работы нервной системы, гормонов и печени.
Печень– единственный орган, депонирующий глюкозу (в виде гликогена) для нужд всего организма. Благодаря активной фосфатазе глюкозо-6-фосфата гепатоциты способны образовыватьсвободнуюглюкозу, которая, в отличие от ее фосфорилированных форм, может проникать через мембрану клеток в общий круг кровообращения.
Выдающуюся роль среди гормонов играет инсулин. Он оказывает действие только на инсулинзависимые ткани, прежде всего, на мышечную и жировую. Мозг, лимфатическая ткань, эритроциты относятся к инсулиннезависимым. Поступление глюкозы в печеночные клетки и выход глюкозы в кровь из печени также являются независимыми от инсулина процессами.
Наиболее существенным действием инсулина на организм является снижение нормального или повышенного уровня глюкозы в крови – вплоть до развития гипогликемического шока при введении высоких доз инсулина. Уровень глюкозы в крови снижается в результате: 1) ускорения поступления глюкозы в клетки; 2)повышения использования глюкозы клетками.
Инсулин ускоряет поступление моносахаридов в инсулинзависимые ткани, особенно глюкозы (а также сахаров схожей конфигурации в положении С-1– С-3), но не фруктозы. Связывание инсулина со своим рецептором на плазматической мембране приводит к перемещению запасных белков-переносчиков глюкозы (ГЛУТ 4) из внутриклеточных депо и включению их в мембрану.
Инсулин активирует использование клетками глюкозы путем:
активирования и индукции синтеза ключевых ферментов гликолиза (глюкокиназы, фосфофруктокиназы, пируваткиназы);
увеличения включения глюкозы в пентозофосфатный путь (активирование дегидрогеназ глюкозо-6-фосфата и 6-фосфоглюконата);
повышения синтеза гликогена за счет стимуляции образования глюкозо-6-фосфата и активирования гликогенсинтазы (одновременно инсулин ингибирует гликогенфосфорилазу);
торможения активности ключевых ферментов глюконеогенеза (пируваткарбоксилазы, фосфоенолпируваткарбоксикиназы, дифосфатазы, глюкозо-6-фосфатазы) и репрессии их синтеза (уставлен факт репрессии гена фосфоенолпируваткарбоксикиназы).
Другие гормоны, как правило, способствуют увеличению содержания глюкозы в крови.
Глюкагони адреналинприводят к росту гликемии путем активации гликогенолиза в печени (активирование гликогенфосфорилазы), однако в отличие от адреналина глюкагон не влияет на гликогенфосфорилазу мышц. Кроме того, глюкагон активирует глюконеогенез в печени, следствием чего также является увеличение концентрации глюкозы в крови.
Глюкокортикоиды способствуют повышению уровня глюкозы в крови за счет стимуляции глюконеогенеза (ускоряя катаболизм белков в мышечной и лимфоидной тканях, эти гормоны увеличивают содержание в крови аминокислот, которые, поступая в печень, становятся субстратами глюконеогенеза). Кроме того, глюкокортикоиды препятствуют утилизации глюкозы клетками организма.
Гормон роста вызывает увеличение гликемии опосредованно: стимулируя распад липидов, он приводит к увеличению уровня жирных кислот в крови и клетках, снижая тем самым потребность последних в глюкозе (жирные кислоты – ингибиторы использования глюкозы клетками).
Тироксин, особенно вырабатываемый в избыточных количествах при гиперфункции щитовидной железы, также способствует повышению уровня глюкозы в крови (за счет увеличения гликогенолиза).
При нормальном уровне глюкозы в крови почки полностью ее реабсорбируют и сахар в моче не определяется. Однако если гликемия превышает 9–10 ммоль/л (почечный порог), то появляетсяглюкозурия. При некоторых поражениях почек глюкоза может обнаруживаться в моче и при нормогликемии.
В норме содержание глюкозы в крови натощак обычно ниже 6 ммоль/л, уровень в пределах 6–8 ммоль/л должен рассматриваться как пограничное состояние, а равный или превышающий 8 ммоль/л может служить диагнозом сахарного диабета.
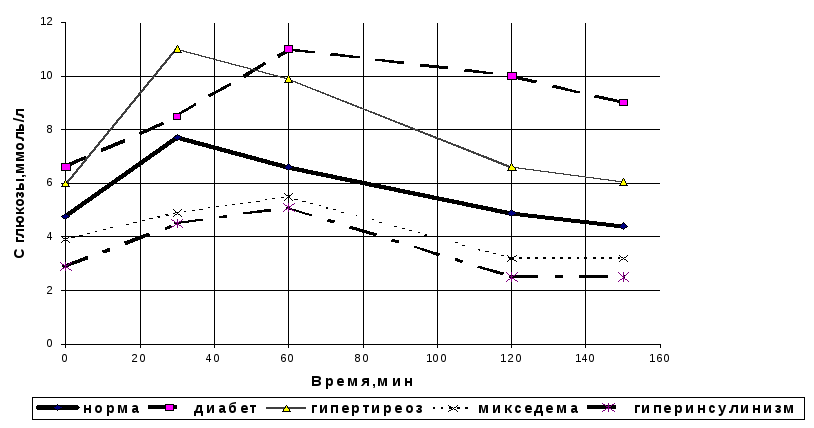 Проверка способности организма регулировать содержание глюкозы в крови (толерантность к глюкозе) используется для диагностики сахарного диабета при постановке перорального глюкозо-толерантного теста. Первая проба крови берется натощак после ночного голодания. Затем больному в течение 5 мин. дают выпить раствор глюкозы из расчета 1 г глюкозы на кг массы тела (1,5 г сахарозы) в 200 мл воды. После этого каждые 30 мин на протяжении 2 ч определяют содержание глюкозы в крови и полученные результаты используют для построения “сахарных кривых”. Некоторые варианты таких кривых в сравнении с данными, полученными у здорового человека, приводятся на рис.5.21. Гликемические кривые у детей имеют такой же характер, что и у взрослых, с тем лишь отличием, что повышение концентрации глюкозы в крови у детей после нагрузки меньшее.
Проверка способности организма регулировать содержание глюкозы в крови (толерантность к глюкозе) используется для диагностики сахарного диабета при постановке перорального глюкозо-толерантного теста. Первая проба крови берется натощак после ночного голодания. Затем больному в течение 5 мин. дают выпить раствор глюкозы из расчета 1 г глюкозы на кг массы тела (1,5 г сахарозы) в 200 мл воды. После этого каждые 30 мин на протяжении 2 ч определяют содержание глюкозы в крови и полученные результаты используют для построения “сахарных кривых”. Некоторые варианты таких кривых в сравнении с данными, полученными у здорового человека, приводятся на рис.5.21. Гликемические кривые у детей имеют такой же характер, что и у взрослых, с тем лишь отличием, что повышение концентрации глюкозы в крови у детей после нагрузки меньшее.
Рис.5.21. Гликемические кривые при однократной нагрузке глюкозой в норме и при некоторых патологических состояниях
Таблица 5.3.Критерии ВОЗ, используемые при диагностике сахарного диабета и нарушения толерантности к глюкозе
Источник
Регуляция обмена углеводов
Энергетический гомеостаз обеспечивает энергетические потребности тканей с использованием различных субстратов. Т.к. углеводы являются основным источником энергии для многих тканей и единственным для анаэробных, регуляция углеводного обмена является важной составляющей энергетического гомеостаза организма.
Регуляция углеводного обмена осуществляется на 3 уровнях:
1. Центральный уровень регуляции углеводного обмена
Центральный уровень регуляции осуществляется с участием нейроэндокринной системы и регулирует гомеостаз глюкозы в крови и интенсивность метаболизма углеводов в тканях. К основным гормонам, поддерживающим нормальный уровень глюкозы в крови 3,3-5,5 мМоль/л, относят инсулин и глюкагон. На уровень глюкозы влияют также гормоны адаптации – адреналин, глюкокортикоиды и другие гормоны: тиреоидные, СДГ, АКТГ и т.д.
2. Межорганный уровень регуляции углеводного обмена
Глюкозо-лактатный цикл (цикл Кори) Глюкозо-аланиновый цикл

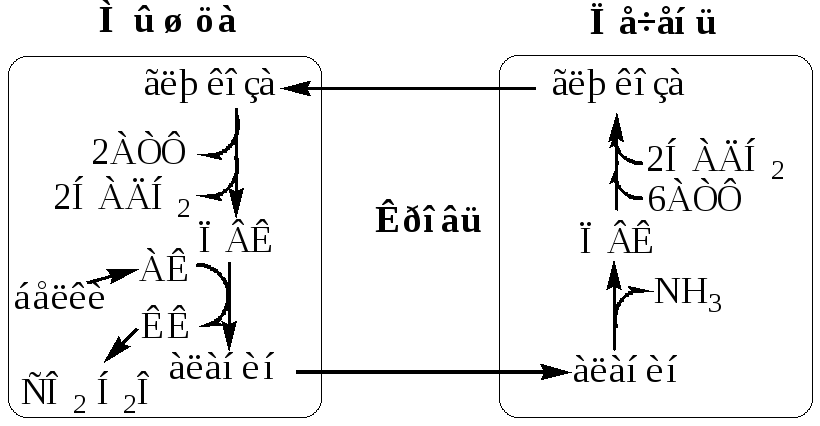
Глюкозо-лактатный цикл не требует наличие кислорода, функционирует всегда, обеспечивает: 1) утилизацию лактата, образующегося в анаэробных условиях (скелетные мышцы, эритроциты), что предотвращает лактоацидоз; 2) синтез глюкозы (печень).
Глюкозо-аланиновый циклфункционирует в мышцах при голодании. При дефиците глюкозы, АТФ синтезируется за счет распад белков и катаболизма аминокислот в аэробных условиях, при этом глюкозо-аланиновый цикл обеспечивает: 1) удаление азота из мышц в нетоксичной форме; 2) синтез глюкозы (печень).
3. Клеточный (метаболический) уровень регуляции углеводного обмена
Метаболический уровень регуляции углеводного обмена осуществляется с участием метаболитов и поддерживает гомеостаз углеводов внутри клетки. Избыток субстратов стимулирует их использование, а продукты ингибируют свое образование. Например, избыток глюкозы стимулирует гликогенез, липогенез и синтез аминокислот, дефицит глюкозы — глюконеогенез. Дефицит АТФ стимулирует катаболизм глюкозы, а избыток – наоборот ингибирует.
IV. Педфак. Возрастные особенности ПФШ и ГНГ, значение.
ГОСУДАРСТВЕННАЯ МЕДИЦИНСКАЯ АКАДЕМИЯ
Зав. каф. проф., д.м.н.
Тема: Структура и обмен инсулина, его рецепторов, транспорт глюкозы.
Механизм действия и метаболические эффекты инсулина.
Факультеты: лечебно-профилактический, медико-профилактический, педиатрический. 2 курс.
Гормоны поджелудочной железы
Поджелудочная железа выполняет в организме две важнейшие функции: экзокринную и эндокринную. Экзокринную функцию выполняет ацинарная часть поджелудочной железы, она синтезирует и секретирует панкреатический сок. Эндокринную функцию выполняют клетки островкового аппарата поджелудочной железы, которые секретируют пептидные гормоны, участвующие в регуляции многих процессов в организме.1-2 млн. островков Лангерганса составляют 1-2% массы поджелудочной железы.
В островковой части поджелудочной железы выделяют 4 типа клеток, секретирующих разные гормоны: А- (или α-) клетки (25%) секретируют глюкагон, В- (или β-) клетки (70%) — инсулин, D- (или δ-) клетки ( + или Са 2+ в крови; 6. лекарства, производные сульфонилмочевины (глибенкламид).
Под влиянием соматостатина секреция инсулина понижается. β-клетки также находятся под влиянием автономной нервной системы. Парасимпатическая часть (холинергические окончания блуждающего нерва) стимулирует выделение инсулина. Симпатическая часть (адреналин через α2-адренорецепторы) подавляет выделение инсулина.
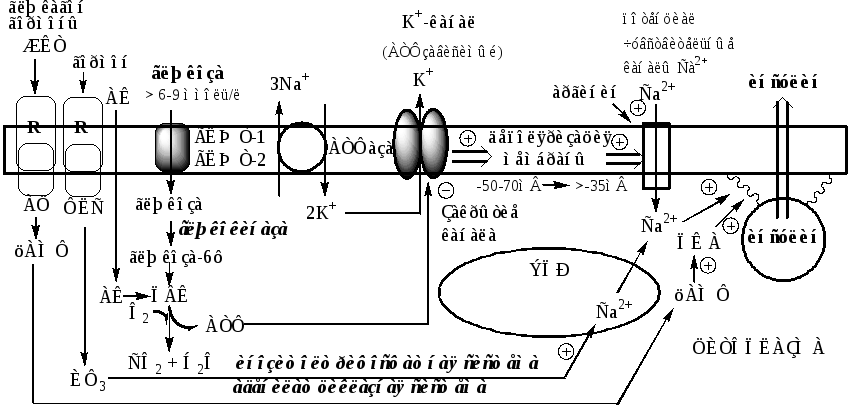
Секреция инсулина осуществляется с участием нескольких систем, в которых основная роль принадлежит Са 2+ и цАМФ.
Поступление Са 2+ в цитоплазму контролируется несколькими механизмами:
1). При повышении концентрации глюкозы в крови выше 6-9 ммоль/л, она при участии ГЛЮТ-1 и ГЛЮТ-2 поступает в β-клетки и фосфорилируется глюкокиназой. При этом концентрация глюкозо-6ф в клетке прямо пропорциональна концентрации глюкозы в крови. Глюкозо-6ф окисляется с образованием АТФ. АТФ образуется также при окислении аминокислот и жирных кислот. Чем больше в β-клетке глюкозы, аминокислот, жирных кислот тем больше из них образуется АТФ. АТФ ингибирует на мембране АТФ-зависимые калиевые каналы, калий накапливается в цитоплазме и вызывает деполяризацию клеточной мембраны, что стимулирует открытие потенциалзависимых Са 2+ -каналов и поступление Са 2+ в цитоплазму.
2). Гормоны, активирующие инозитолтрифосфатную систему (ТТГ), выпускают Са 2+ из митохондрий и ЭПР.
цАМФобразуется из АТФ с участием АЦ, которая активируется гормонами ЖКТ, ТТГ, АКТГ, глюкагоном и Са 2+ -кальмодулиновым комплексом.
цАМФ и Са 2+ стимулируют полимеризацию субъединиц в микротубулы (микроканальцы). Влияние цАМФ на микроканальцевую систему опосредуется через фосфорилирование ПК А микроканальцевых белков. Микроканальцы способны сокращаться и расслабляться, перемещая гранулы по направлению к плазматической мембране обеспечивая экзоцитоз.
Секреция инсулина в ответ на стимуляцию глюкозой представляет собой двухфазную реакцию, состоящую из стадии быстрого, раннего высвобождения инсулина, называемую первой фазой секреции (начинается через 1 мин, продолжается 5-10 мин), и второй фазы (продолжительность ее до 25-30 мин).
Транспорт инсулина. Инсулин водорастворим и не имеет белка-переносчика в плазме. Т1/2 инсулина в плазме крови составляет 3—10 мин, С-пептида — около 30 мин, проинсулина 20-23 мин.
Разрушение инсулина происходит под действием инсулинзависимой протеиназы и глутатион-инсулин-трансгидрогеназы в тканях мишенях: в основном в печени (за 1 проход через печень разрушается около 50% инсулина), в меньшей степени в почках и плаценте.
Источник



