
Регуляция метаболизма на уровне клеток
Регуляторные механизмы, работающие на уровне клеток можно условно разделить на механизмы специфической и неспецифической регуляции их метаболизма.
Механизмы неспецифической регуляции клеточного метаболизма основаны на действии факторов, изменяющих активность многих ферментов, и влияющих на уровень клеточного метаболизма в целом.
К механизмам неспецифической регуляции могут быть отнесены:
изменение концентрации соединений в среде, окружающей клетку. Так, концентрация глюкозы в крови, а соответственно, и в межклеточной жидкости, может изменяться в норме в 2 раза, содержание жирных кислот — в несколько раз, а содержание ацетоновых тел при голодании может возрастать в десятки раз. Увеличение концентрации субстратов приводит к повышению скорости ферментативных реакций и увеличению потока метаболитов по соответствующим метаболическим путям. Собственно именно на этот эффект рассчитывают медики, вводя парентерально раствор глюкозы ослабленным больным.
изменение температуры тела при различных заболеваниях может достигать 41°С, т.е. повышаться на 4–5°С, что увеличивает скорость ферментативного катализа минимум в 1,5 раза. Следствием этого будет интенсификация обменных процессов.
Снижение температуры, наоборот, приводит к замедлению метаболизма, что используется иногда в хирургии при проведении операций на органах с временным отключением в этих органах кровообращения. Замедление метаболизма в клетках, тканях или органах в условиях пониженных температур используется при консервации органов с целью их последующей пересадки.
Специфические механизмы регуляции метаболизма клеток.
К более специфическим механизмам регуляции клеточного метаболизма относятся:
Изменение активности ферментов;
Изменение количества ферментов;
Изменение проницаемости клеточных мембран.
Изменение активности ферментов
Изменение активности имеющихся в клетке ферментов относится к механизмам экстренной или срочной регуляции метаболизма, поскольку метаболический ответ клетки формируется на базе уже имеющихся в клетке ферментов.
Второй важной особенностью этих механизмов является обратимость их действия, что крайне важно для возврата клетки к исходному состоянию. Ферменты, активность которых может изменяться в зависимости от потребности клеток, получили название регуляторных ферментов.
Изменение активности регуляторных ферментов может происходить в клетке с участием трех основных механизмов:
Аллостерическая модуляция
При аллостерической модуляции регуляторный фермент имеет в своей структуре один или несколько аллостерических центров, способных высоко избирательно взаимодействовать с низкомолекулярными соединениями — аллостерическими модуляторами за счет слабых взаимодействий.
В результате этого взаимодействия изменяется конформация белка-фермента, в том числе несколько изменяется и структура активного центра, что сопровождается изменением эффективности катализа.
Связывание легко обратимо: при снижении концентрации модулятора в среде окружения,комплекс фермент-модулятор диссоциирует и фермент восстанавливает свою исходную конформацию, а, следовательно, и каталитическую активность.
В качестве аллостерических модуляторов в клетке выступают обычно промежуточные метаболиты или конечные продукты того или иного метаболического пути.
Наиболее часто встречается ингибирование по принципу отрицательной обратной связи или ретроингибирование. В этом случае конечный продукт метаболического пути ингибирует активность регуляторного фермента, катализирующего одну из начальных реакций того же метаболического пути:
 Второй вариант — активация предшественником. В этом случае один из промежуточных метаболитов, образующихся в начале метаболического пути, выступает в качестве активатора того или иного фермента, катализирующего одну из конечных реакции того же самого метаболического пути:
Второй вариант — активация предшественником. В этом случае один из промежуточных метаболитов, образующихся в начале метаболического пути, выступает в качестве активатора того или иного фермента, катализирующего одну из конечных реакции того же самого метаболического пути:
Источник
Регуляция метаболических процессов
Живая клетка – это открытая система, которая постоянно обменивается с внешней средой веществами и энергией. В клетку поступают питательные вещества, которые используются в качестве строительного и энергетического материала, из клетки выводятся конечные продукты метаболизма.
В клетке постоянно происходит большое количество разнообразных химических реакций, которые формируют метаболические пути – последовательность превращения одних соединений в другие. Метаболизм – совокупность всех метаболических путей, протекающих в организме.
Выделяют – катаболизм (распад сложных веществ до простых с высвобождением энергии) и анаболизм (синтез более сложных веществ из простых веществ).
Все пути согласованы между собой во времени и пространстве. Эта согласованность протекания метаболических процессов обеспечивается сложными механизмами регуляции.
Организация химических реакций в метаболические пути
Оптимальная активность ферментов, регулирующих реакции метаболического пути, достигается благодаря определенной организации в клетке.
Пространственная локализация ферментов
Большинство ферментов локализовано внутри клетки, причем ферменты одного метаболического пути находятся в одном отделе клетки. Разделение метаболических путей важно для противоположно направленных процессов. Например, синтез жирных кислот происходит в цитоплазме, а их распад в митохондриях. Если бы такого разделения не существовало, то возникали бы бесполезные с физиологической точки зрения пути.
В метаболических путях продукт первой реакции служит субстратом второй и так далее до формирования конечного продукта. Промежуточные продукты одного пути могут высвобождаться из последовательных реакций и использоваться в других метаболических путях, т.е. все метаболические пути связаны между собой.
Пространственная организация ферментов может быть настолько выражена, что продукт реакции не может быть вычленен из метаболического пути и обязательно служит субстратом следующей реакции.
Такая организация метаболического пути называется мультиферментным комплексом. Эти комплексы связаны с мембранами. Пример такого комплекса – пируватдегидрогеназный комплекс, под действием которого происходит окислительное декарбоксилирование пировиноградной кислоты.
Структура метаболических путей
Метаболические пути подразделяются на 4 типа. Если субстрат превращается в один продукт, то такой путь называется линейным (гликолиз). Чаще встречаются разветвленные пути – когда синтезируются разные продукты в зависимости от потребности клетки (синтез нуклеотидов). Также существуют циклический (цикл трикарбоновых кислот) и спиральный (β-окисление жирных кислот) метаболические пути.
Ферменты находятся во всех клетках организма. В процессе дифференцировки клеток изменяется и их ферментный состав. Например, фермент аргиназа, участвующая в синтезе мочевины, находится в клетках печени. Это так называемый органоспецифичный фермент.
Функционирование клетки обеспечивается пространственной и временной регуляцией метаболических путей. Пространственная регуляция связана с локализацией определенных ферментов в различных органеллах. В ядре находятся ферменты, связанные с синтезом ДНК и РНК, в цитоплазме – ферменты гликолиза, в лизосомах – гидролитические ферменты, в митохондриях – ферменты цикла трикарбоновых кислот и цепи переноса электронов.
Принципы регуляции метаболических путей
Все химические реакции в клетке протекают при участии ферментов. Чтобы воздействовать на скорость протекания метаболического пути, достаточно регулировать количество или активность ферментов. В каждом метаболическом пути есть ключевые ферменты, которые регулируют скорость всего пути. Эти ферменты называются регуляторными.
Регуляция скорости ферментативных реакций осуществляется на 3 уровнях:
Изменение количества молекул фермента
Доступность субстрата и кофермента
Изменение каталитической активности фермента
Регуляция количества молекул фермента в клетке
В клетке постоянно происходит синтез и распад белковой молекулы фермента
синтез
Распад
Регуляция синтеза фермента может происходить на любой стадии формирования белковой молекулы. Наиболее изучена регуляция синтеза белковой молекулы на уровне транскрипции, которая осуществляется гормонами и биологически активными молекулами. Распад ферментов менее изучен.
Регуляция скорости ферментативной реакции доступностью субстрата и кофермента
Главным и необходимым параметром, регулирующим скорость метаболического пути, является наличие первого субстрата. Чем выше его концентрация, тем выше скорость метаболического пути.
Другим параметром является наличие регенерированных коферментов. В реакциях дегидрирования коферментом дегидрогеназ служат окисленные формы НАД+, ФАД, ФМН, которые восстанавливаются в ходе реакции. Чтобы коферменты вновь участвовали в реакции, необходимо, что они вновь превратились в окисленную форму.
Регуляция каталитической активности фермента
Регуляция с помощью белок-белковых взаимодействий
Регуляция путем фосфорилирования/дефосфорилирования фермента
Ферменты, имеющие такой механизм регуляции являются, как правило, олигомерными белками. Они состоят из нескольких (не менее 2 х ) субъединиц, имеют активный и аллостерический центры, которые находятся на разных субъединицах. Присоединение эффектора (клеточного метаболита) в аллостерический центр вызывает кооперативные конформационные изменения всех протомеров.
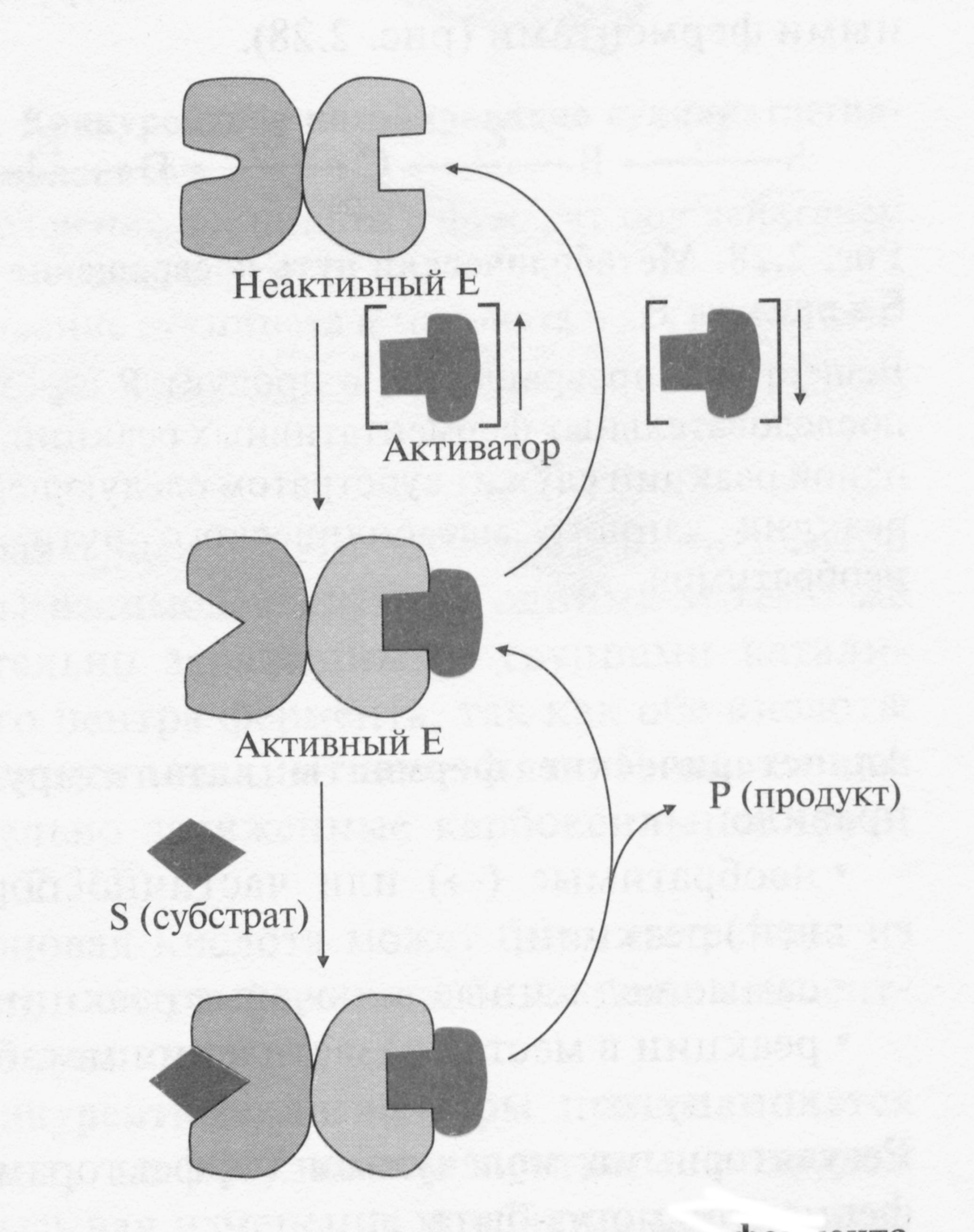
Если в аллостерическом центре связывается эффектор (активатор), повышается связывание субстрата в активном центре и возрастает скорость реакции, которую катализирует этот фермент. Конформационные перестройки в активном центре фермента повышают или понижают его сродство к субстрату.
При увеличении в клетке концентрации активатора возрастает скорость его связывания в аллостерическом центре. Изменяется конформация регуляторной субъединицы фермента, происходят кооперативные конформационные изменения в ферменте, изменяется конформация активного центра фермента, повышается сродство фермента к субстрату и скорость ферментативной реакции. При понижении концентрации аллостерического активатора снижается скорость связывания регуляторного лиганда в аллостерическом центре. Изменяется конформация регуляторной субъединицы, происходят кооперативные конформационные изменения в ферменте, изменяется конформация активного центра, снижается сродство к субстрату и понижается скорость реакции.
Если же эффектором является ингибитор, то сродство фермента к субстрату и скорость превращения его в продукт снижаются.


Аллостерические ферменты регулируют скорость метаболических путей, которые представляют собой последовательность взаимосвязанных реакций, катализируемых разными ферментами
S B C D P 



Вещество S превращается в продукт Р в результате 4 последовательных ферментативных реакций. Продукт одной реакции служит субстратом следующей.
Аллостерические ферменты катализируют:
Необратимые или частично обратимые реакции
Самые медленные, ключевые реакции
Реакции в местах разветвления метаболического пути
Конечные продукты метаболических путей
Субстраты метаболических путей
Промежуточные метаболиты или специфические молекулы
Например, катаболизм глюкозы до СО2 и Н2О регулируется аллостерически.
глюкоза В С М N ….. ……. СО2, Н2О, АТФ




Значение данного процесса состоит в синтезе АТФ в клетке за счет катаболизма глюкозы. При увеличении отношения АТФ/АДФ скорость реакций данного метаболического пути снижается. Из представленной выше последовательности ферментативных реакций аллостерическим является Е3, так как он катализирует необратимую самую медленную реакцию.
При повышении уровня АТФ в клетке
АТФ взаимодействует с аллостерическим центром фермента Е3
Происходят кооперативные конформационные изменения фермента Е3
Снижается сродство Е3 к субстрату
Понижается активность и замедляется реакция катализируемая ферментом Е3
Понижается скорость метаболического пути


Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Источник
РЕГУЛЯЦИЯ МЕТАБОЛИЗМА
Живая клетка является открытой динамической саморегулирующейся системой, метаболизм которой зависит как от внутренних потребностей, так и от факторов окружающей среды. Поэтому все существующие в клетке гены экспрессируются не одновременно, а по потребности. Кроме того и активность ферментов меняется в зависимости от нужд клетки. В этом и состоит сущность регуляции метаболизма.
Регуляция метаболизма осуществляется на двух основных уровнях – генетическом и биохимическом. На генетическом уровне обмен веществ регулируется путем регуляции экспрессии генов, а именно усилением или подавлением транскрипции и трансляции. Второй уровень регуляции – биохимический осуществляется за счёт регуляции активности ферментов. Генетическая регуляция – грубый способ настройки метаболизма, биохимическая регуляция – более тонкая настройка.
Молекулярной основой обоих уровней регуляции являются аллостерические ферменты и белки, имеющие обычно два типа активных центров. Один из них служит для присоединения низкомолекулярных эффекторов, которые могут влиять на проявление активности второго активного центра, путем изменения пространственной структуры белка.
При регуляции ферментативной активности (биохимический уровень)– сами ключевые ферменты того или иного метаболического цикла являются аллостерическими. Они имеют два типа активных центров –каталитический(для связывания с субстратом) иэффекторный(для связывания с эффектором – активатором или ингибитором). Если фермент связывается с активатором, изменяется его конформация и, в том числе, пространственная структура каталитического центра. Это способствует облегчению связывания фермента с субстратом и усиливает ферментативную активность. Если эффектор является ингибитором, то его присоединение к эффекторному центру фермента ослабляет или делает невозможным взаимодействие субстрата с каталитическим центром и ведет к понижению или полному угнетению ферментативной активности.
В регуляции экспрессии генов(генетической уровень) также участвуют аллостерические белки. Они выступают в роли белков-регуляторов, которые связываются с ДНК в промоторной зоне гена (или оперона) в области оператора и могут либо усиливать, либо подавлять транскрипцию. Один центр белка-регулятора служит для присоединения к ДНК, второй центр – для связывания эффектора. Аллостерические белки-регуляторы выступают в роли посредников между ДНК и эффектором.
Эффекторами, способными «включать» или «выключать» гены, являются:
в катаболических генах (оперонах)– самисубстраты(например, углеводы), которые подлежат расщеплению, они выступают активаторами для белка-регулятора, т.е. выполняют функцию «включателей» гена;
в анаболических оперонах – конечные продукты синтеза(например, аминокислоты, нуклеотиды), они выступают в роли корепрессоров для белка-регулятора и, связываясь с ним, «выключают» транскрибирование ферментов, необходимых для их собственного синтеза.
Механизмы регуляции метаболизма на генетическом уровне впервые были изучены на прокариотах (в оперонах кишечной палочки) в работах Жакоба и Моно еще в 40х-60х годах ХХ столетия. В настоящее время установлено, что регуляция экспрессии генов осуществляется на уровне транскрипции – при синтезе и-РНК ина уровне трансляции– при синтезе белка на рибосомах.
На уровне транскрипциивыявлены такие механизмы регуляции:
положительный и отрицательный контроль;
индукция и репрессия;
смешанные механизмы регуляции;
регуляция посредством взаимодействия с энхансерами и сайленсерами (у эукариот)
На уровне трансляциивыявлены следующие механизмы:
аттенуация путем образования альтернативных шпилек на и-ДНК (изучена у прокариот),
регуляция трансляции на уровне сборки рибосом;
регуляция трансляции с помощью факторов инициации, элонгации и терминации.
Регуляция на уровне транскрипции
Положительный и отрицательный контрольрегуляции работы генов (или оперонов) реализуется с помощью белков-регуляторов (R)и базируется на их природе.При положительном контролебелок-регулятор, связываясь с оператором, ускоряет сборку РНК-полимеразы и, следовательно, усиливает транскрипцию.При отрицательном контролебелок-регулятор (R) блокирует оператор, который часто перекрывается с промотором, и препятствует сборке РНК-полимеразы либо её продвижению по матричной цепи ДНК. Белки-регуляторы кодируются отдельными генами, расположенными вблизи оперона, внутри оперона среди структурных генов (при аутогенном контроле), либо далеко за пределами оперона.
Индукция и репрессия. Осуществление таких механизмов регуляции экспрессии генов, как индукция и репрессия, реализуется благодаря эффекторам. Генетические системы чутко реагируют на присутствие в клетке и окружающей среде питательных субстратов, тех или иных важных метаболитов, поэтому экспрессия «нужных» генов начинается только после появления в среде или в клетке соответствующих эффекторов – субстратов или конечных продуктов синтеза. Белки-регуляторы являются посредниками между молекулой ДНК и эффекторами, запускающими тот или иной метаболический путь.
Для катаболических оперонов и генов характерна индукция. При этом эффекторами белков R являются сами субстраты (глюкоза, лактоза, глицерин и др.). Их называют индукторами и они являются эффекторами, усиливающими транскрипцию. В этом есть глубокий биологический смысл. Так, например, лактозный оперон, кодирующий три фермента катаболизма лактозы (бета-галактозидазу, пермеазу и трансацетилазу) включается только тогда, когда в среде появляется эффектор – субстрат лактоза, выступающий в качестве индуктора.
Для анаболических оперонов и генов характерна репрессия. При этом эффекторами для белков-регуляторов являются конечные продукты синтеза (например, аминокислоты, нуклеотиды), которые выступают в роли корепрессоров и способны угнетать транскрипцию. Например, уE. Coli аминокислота триптофан, накопленная клеткой в результате синтеза в избыточном количестве, как корепрессор белка-регулятора (репрессора) блокирует синтез ферментов, закодированных в 5 генах триптофанового оперона. Т.е. эффектором (корепрессором) триптофанового оперона является сам триптофан.
Обобщая сказанное выше, еще раз подчеркнем:
индукция характерна для катаболических оперонов, эффекторами являются субстраты, выступающие в роли индукторов;
репрессия характерна для анаболических оперонов, эффекторами являются конечные продукты синтеза, выступающие в роли корепрессоров.
В условиях «invivo» положительный и отрицательный контроль почти всегда сочетаются с индукцией и репрессией, т.е. в регуляции принимают участие и белки-регуляторы и эффекторы. Поэтому выделяют4 типа классических оперонов: 2 индуцибельных оперона – соответственно с положительным и отрицательным контролем и 2 репрессибельных оперона – также с положительным и отрицательным контролем. Рассмотрим их.
Индуцибельный оперон с отрицательным контролем – сочетание отрицательного контроля и индукции. Такой тип регуляции был обнаружен в лактозном оперонеE. coli. Регуляция осуществляется следующим образом. Если в среде нет лактозы, то белок- регулятор, выступая в качестве репрессора (отрицательный контроль), блокирует операторную зону в данном опероне и препятствует транскрибированию генов. Как только лактоза появляется в среде, она как эффектор-индуктор связывается с белком-регулятором, изменяет его конформацию и уменьшает сродство к ДНК. В результате белок-регулятор (репрессор) отпадает от оператора, что «открывает путь» для РНК-полимеразы. Последняя осуществляет транскрипцию генов, кодирующих ферменты для катаболизма (утилизации) лактозы. Таким образом, лактоза путем индукции включает собственный оперон.
Индуцибельный оперон с положительным контролем – сочетание положительного контроля и индукции. Этот тип регуляции был обнаружен также уE. coliв мальтозном, рамнозном и арабинозном оперонах. Рассмотрим этот механизм на примере арабинозного оперона, который состоит из трех геновara B, ara A, ara D, кодирующих ферменты для превращенияL-арабинозы вD-ксилулозо-5-фосфат. В данном опероне белок-регулятор выполняет двоякую функцию. При отсутствии арабинозы в среде белок-регулятор выступает в качестве репрессора (отрицательный контроль), присоединяется к оператору и препятствует транскрипции. При появлении субстрата арабинозы в среде, она как индуктор связывается с белком-регулятором и, меняя его конформацию, превращает в белок-активатор (положительный контроль). Этот изменённый белок скользит в область промотора и, как активатор, усиливает сборку РНК-полимеразы и, следовательно, стимулирует транскрипцию. Следует отметить, что несмотря на название — «индуцибельный оперон с пложительным контролем» — на самом деле здесь имеет место более сложный тип регуляции, а именно, при отсутствии индуктора оперон «закрыт» белком-регулятором (отрицательный контроль), а появление индуктора «открывает» оперон для считывания (положительный контроль).
Репрессибельный оперон с отрицательным контролем – сочетание отрицательного контроля и репрессии. Обнаружено в триптофановом оперонеE. coli,содержащем 5 генов ферментов, участвующих в синтезе триптофана из хоризмовой кислоты (хоризмата). Это типичный анаболический оперон, подверженный регуляции по типу репрессии. Регуляция его транскрипции осуществляется следующим образом. Если в клетке недостаточно триптофана, то оперон беспрепятственно считывается РНК-полимеразой и обеспечивает продукцию ферментов для синтеза триптофана. При этом белок-регулятор имеется в клетке, но в неактивной форме – в виде апорепрессора (неполноценного репрессора). Как только в клетке накапливается избыток триптофана, он как корепрессор соединяется с апорепрессором и делает его полноценным репрессором. Присоединение репрессора к оператору вызывает блокирование транскрипции (отрицательный контроль), в результате чего прекращается синтез ферментов и самого триптофана.
Репрессибельный оперон с положительным контролем – сочетание положительного контроля и репрессии. Такой тип регуляции пока не обнаружен, но предполагается, что белок-регулятор при недостатке конечного продукта в клетке выступает в роли белка-активатора (положительный контроль) и активирует транскрипцию. При накоплении конечного продукта в избытке он сам выступает в качестве корепрессора, связывается с белком-регулятором и превращает его в репрессор. Это приводит к блокированию синтеза ферментов и, как следствие, самого конечного продукта.
Катаболитная репрессия является примером смешанного механизма регуляции и первоначально была обнаружена в описанном ранее лактозном оперонеE. coliпри явлении, получившем названиедиауксия. Это явление наблюдается в том случае, если в среде имеется несколько углеводов (например, глюкоза и лактоза) и сводится к тому, что сначала утилизируется глюкоза, как легко усвояемый субстрат, за счет вовлечения в гликолиз. При этом лактозный оперон будет заблокирован до тех пор, пока глюкоза не будет исчерпана, т.е. синтез ферментов, расщепляющих второй субстрат, репрессируется. Отсюда и название – «катаболитная репрессия». По мере утилизации глюкозы происходит дерепрессия лактозного оперона и начинается транскрипция генов катаболизма лактозы. Рассмотрим, как реализуется этот механизм.
В лактозном опероне существует две системы регуляции. Первый регуляторный механизмуже был описан ранее как пример индуцибельного оперона с отрицательным контролем. Он релизуется в области оператора за счет белка-репрессора, действие которого убирается за счет связывания с индуктором лактозой. Однако этого недостаточно, чтобы началось транскрибирование генов оперона, поскольку оперон не может «включиться» до тех пор, пока с соседней областью промотора не свяжется другой специфический белок-регулятор САР (cataboliteactivatorprotein), т.е. пока не сработаетвторая система регуляции. Связывание САР с промотором является необходимым условием присоединения РНК-полимеразы к ДНК. Однако эффектором САР является циклический АМФ (цАМФ), который в достаточном количестве появляется только после исчерпания глюкозы. Пока в среде есть глюкоза, она вовлекается в гликолиз с образованием АТФ. Как только глюкоза исчерпается, то количесто АТФ в клетке резко уменьшается, но возрастает концентрация цАМФ, который является сигналом голода. Под воздействием цАМФ белок САР модифицируется и как активатор присоединяется к промотору, ускоряя присоединение и сборку РНК-полимеразы и, тем самым, делая возможной транскрипцию генов лактозного оперона, то есть клетка переходит на питание лактозой.
Таким образом, в лактозном опероне функционирует две системы регуляции– однана промоторе (индукция с положительным контролем), другая –на операторе(индукция с отрицательным контролем). На операторе контроль осуществляется с помощью белка-репрессора и индуктора лактозы, на промоторе «включение» оперона обеспечивается благодаря присоединению белка-регулятора САР и его эффектора цАМФ.
Аутогенный контроль(или аутогенная регуляция) осуществляется в тех оперонах, где один из структурных генов кодирует белок с двойными фунциями – и фермента и белка-регулятора. Принцип аутогенной регуляции состоит в том, что белок-регулятор управляет транскрипцией оперона и тем самым влияет на собственный синтез. То есть происходит саморегуляция оперона. Причем такой тип регуляции встречается в оперонах и с положительным и с отрицательным контролем. Это достаточно тонкая система регуляции, позволяющая приостановить или же наоборот активировать транскрипцию в соответствии с потребностями клетки на данный момент. Наиболее изучен подобный механизм на примере катаболического гистидиновогооперона hutу сальмонелл, в котором один из генов кодирует синтез фермента, одновременно выступающего в роли белка-регулятора, притормаживающего транскрипцию.
Регуляция экспрессии генов на уровне трансляции
Аттенуацияосуществляется на уровне трансляции путем образования альтернативных шпилек на и-РНК, проходящей через рибосому. Этот тип регуляции обнаружен уE. coliЧ.Яновским в 1983 г. при трансляции полицистронной и-РНК, считанной соперона trpи содержащей гены ферментов, обеспечивающих синтез триптофана.
Образуемая в оперона trpи-РНК содержит на 5/-конце лидерную последовательность, включающую повышенную долю кодонов для той аминокислоты (триптофана), синтез которой определяется данным опероном. Если в клетке недостаточно триптофана, то и-РНК своей лидерной последовательностью проходит через пептидильные и аминоацильные центры рибосомы «вхолостую», так как к месту синтеза не подходят нагруженные триптофаном т-РНК. В этом случае в лидерной последовательности, прокатанной через рибосому, за счет палиндрома формируется «шпилька», однако в области структурных генов альтернативная шпилька не образуется. Поэтому основная структурная часть и-РНК беспрепятственно транслируется с образованием ферментов, необходимых для синтеза триптофана.
Если же в клетке имеется избыток триптофана и к рибосомам подходят нагруженные триптофаном т-РНК, то лидерная последовательность транслируется, зато шпилька образуется в зоне структурных генов, что приводит к преждевременной терминации трансляции. В результате этого ферменты синтеза триптофана не образуются и синтез самого триптофана прекращается.
Механизм аттенуации обнаружен не только в оперонах биосинтеза аминокислот, но также при синтезе бета-лактамазы у E. coli.
Источник



