
Регуляция экспрессии генов
Экспрессия генов — это реализация заложенной в них информации, то есть синтез РНК и белков. Другими словами, под экспрессией генов понимают их активность.
В клетках живых организмов экспрессия генов регулируется: одни гены могут быть реализованы, другие — нет. Причем регуляция может осуществляться на разных этапах: может выполняться или нет транскрипция, из пре-мРНК в результате альтернативного сплайсинга могут образовываться разные мРНК, может блокироваться трансляция и др.
У эукариот, обладающих отграниченным от цитоплазмы ядерным содержимым и более сложным геномом, регуляция экспрессии генов намного разнообразнее и сложнее, чем у прокариот.
Регуляция экспрессии генов у прокариот
У прокариот пока молекула РНК синтезируется на участке ДНК, она тут же может транслироваться (начиная с уже синтезированного конца). Поэтому у них регуляция экспрессии (активности) генов осуществляется почти исключительно на уровне ДНК, так как в РНК часто невозможно внести какие-нибудь изменения до ее трансляции.
В 1961 г. Жакобом и Моно была предложена модель оперона как системы регуляции генов у бактерий. Оперон состоит из промотора, оператора, структурных генов оперона (их может быть разное количество) и терминатора. В области промотора прикрепляется фермент РНК-полимераза. В области оператора присоединяется белок-репрессор, который кодируется отдельно отстоящим от оперона геном-регулятором (может быть сцеплен со своим опероном, а может находиться на расстоянии).
Если белок-репрессор соединяется с оператором, то транскрипция всех структурных генов оперона становится невозможной, так как РНК-полимераза не может перемещаться по цепи ДНК.
В свою очередь активность белка-репрессора может блокироваться определенным для него низкомолекулярным соединением — индуктором (тем или иным питательным веществом бактерий). В результате взаимодействия с индуктором белок-репрессор видоизменяется и уже не может присоединиться к оператору своего оперона. В этом случае гены оперона экспрессируются (т. е. на них идет синтез).
Бывает обратная ситуация, когда индуктор активирует белок-репрессор.
Таким образом, в зависимости от того, какие индукторы находятся в цитоплазме, у прокариот экспрессируются те или иные генные группы.
Вышеописанный механизм экспрессии генов относится к негативной регуляции, так как гены транскрибируются, если они не выключены репрессором. И наоборот: не транскрибируются, если выключены.
Кроме негативной регуляции у бактерий существует также позитивная. В этом случае вместо белка-репрессора действие оказывает белок-активатор. На эти белки также действуют индукторы, активируя или инактивируя их.
Также у прокариот были выявлены опероны, которые актируются двумя регуляторными белками, соединенными друг с другом.
Регуляция экспрессии генов у эукариот
У многоклеточных организмов в клетках разных тканей экспрессируются разные гены, т. е. для эукариот характерна дифференциальная экспрессия.
У эукариот, также как и у прокариот, существуют регуляторные белки с похожим механизмом действия. При этом для эукариот не характерна регуляция по типу оперона. Цистроны (транскрибируемые участки) эукариот обычно содержат по одному гену. (Это не касается геномов хлоропластов и митохондрий.)
Кроме регуляторных белков, взаимодействующих с ДНК, у эукариот существуют и другие способы регуляции экспрессии генов.
Конденсация и деконденсация хроматина. Это наиболее универсальный метод регуляции транскрипции. Когда нужно экспрессировать определенные гены, хроматин в этом месте деконденсируется.
Альтернативные промоторы. У гена может быть несколько промоторов, каждый из которых начинает транскрипцию с разных его экзонов в зависимости от типа клетки. В конечном итоге будут синтезированы разные белки.
Метилирование и деметилирование ДНК. Метилирование ДНК происходит в регуляторных областях гена. Метилируется цитозин в последовательности ЦГ, после чего ген инактивируется. При деметилировании активность гена восстанавливается. Процесс регулируется ферментом метилтрансферазой.
Гормональная регуляция. При гормональной регуляции гены активируются в ответ на внешний химический сигнал (поступление в клетку определенного гормона). Этот гормон запускает те гены, которые имеют специфические последовательности нуклеотидов в регуляторных областях.
Геномный импринтинг. Это малоизученный способ регуляции экспрессии генов у эукариот. Он возможен только у диплоидных организмов и выражается в том, что активность генов зависит, от какого из родителей они были получены. Выключение генов осуществляется путем метилирования ДНК.
Альтернативный сплайсинг. Это регуляция на уровне процессинга. При альтернативном сплайсинге порядок сшивки экзонов может быть различным. Отсюда следует, что на основе одной и той же нуклеотидной последовательности ДНК могут быть синтезированы разные белки. Хотя их отличие друг от друга будет в основном заключаться лишь в разных сочетаниях одних и тех же аминокислот.
Тканеспецифическое редактирование РНК также протекает на уровне процессинга. Выражается в замене отдельных нуклеотидов в РНК в определенных тканях организма.
Кроме того, у эукариот иРНК часто не подвергается процессингу вообще (а распадается) или подвергается с задержкой. Это токже можно рассматривать как способ регуляции экспрессии генов.
Регуляция стабильности иРНК. У эукариот существует регуляция и на уровне трансляции, когда готовые иРНК не «допускаются» к рибосомам или разрушаются. Другие же иРНК могут дополнительно стабилизироваться для многократного использования.
Посттрансляционная модификация белка. Чтобы молекула полипептида превратилась в активную молекулу белка, в ней должны произойти различные модификации определенных аминокислот, должны быть сформированы вторичная, третичная и возможно четверичная структуры. На этом этапе также можно повлиять на реализацию генетической информации, например, не дав молекуле сформироваться.
Риборегуляторы. Были обнаружены РНК, выполняющие регуляторные функции путем ослабления работы отдельных генов.
Для высокоорганизованных животных отмечается существование надклеточного уровня регуляции экспрессии генов.
Источник
11.1. Регуляция экспрессии генов у прокариот, на уровне инициации транскрипции.
Этот тип регуляции осуществляется несколькими способами. Из них наиболее значимы два:
1. Регуляция транскрипции путём изменения вторичной структуры участка ДНК, на котором фермент РНК-полимераза осуществляет синтез РНК. Пространственную структуру ДНК изменяют специальные ферменты. Эти изменения могут быть самыми разнообразными, например возможен такой вариант, когда на кодирующей части ДНК формируются изгибы или шпильки. Эти структуры блокируют движение по ДНК ферментов, обеспечивающих самый первый процесс синтеза РНК – инициацию. В результате не происходит деспирализации ДНК, не разрываются водородные связи между нитями ДНК, не отходят друг от друга нити ДНК, т.е. не формируется вилка транскрипции. Процесс транскрипции не начинается.
2. Регуляция путём взаимодействия специальных белков-регуляторов с оператором.
Этот механизм был предложен Ф. Жакобом и Ж. Моно в 1961 г. Предложенная ими гипотеза регуляции в настоящее время полностью подтверждена и хорошо исследована. Этот тип регуляции называется «Контроль синтеза белка на уровне транскрипции» или просто «Теория оперона».
Как мы уже писали ранее, оперон прокариот состоит из промотора, оператора и кодирующей части. К промотору присоединяется РНК-полимераза, которая «проходит» через оператор и транскрибирует РНК с матричной цепи ДНК. Промотор и оператор взаимно перекрываются. К оператору присоединяются два типа белков-регуляторов – активатор и репрессор. Это генетические факторы регуляции.
Репрессор, взаимодействуя с оператором, перекрывает его и частично промотор, создавая стерическое препятствие для присоединения РНК-полимераза к промотору (рис. 70, Б). Такая регуляция носит название негативный контроль. Активатор взаимодействует с оператором таким образом, что эта связь, не только не нарушает движение РНК-полимеразы через оператор (рис. 70, А), но и ускоряет осаждение на промотор всё новых и новых молекул РНК-полимераз. Это повышает интенсивность транскрипции гена, что приводит к существенному увеличению синтеза РНК и синтеза белка. Этот тип регуляции называется позитивным контролем.
Важно отметить, что без активатора связывание РНК-полимеразы с промотором так же происходит, но очень медленно.
Вышеописанный механизм интегрирован и в более высокие уровни регуляции экспрессии генов. Его легко понять, если помнить, что на более высоком уровне регуляции происходят те же процессы, что и изложенные выше, но к ним добавляется звено, которое регулирует действие активаторов и репрессоров. Эндогенные или экзогенные факторы (ими могут быть физические факторы или химические соединения) могут активировать или инактивировать активатор или репрессор. Если происходит активация активатора и инактивация репрессора, то интенсивность транскрипции повысится, И наоборот, при активации репрессора и инактивации активатора интенсивность транскрипции снизится.
Разберём эти процессы более подробно. Рассмотрим три момента, которые определяют принцип регуляции на высоком уровне.
1. На этом уровне в процессе регуляции вовлекаются два новых участника – субстрат и продукт. Субстрат может расщепляться (лактоза), под действием какого либо фермента, или наоборот, синтезироваться из каких либо метаболитов(например, аминокислота). Тогда это будет уже не субстрат, а продукт. Субстрат и продукт относятся к не генетические факторы регуляции. Их часто называют эффекторы.
2. Субстрат и продукт осуществляют регуляцию экспрессии генов путём взаимодействия с генетическими факторами регуляции – активатором и репрессором.
3. В результате взаимодействия с субстратом или продуктом активатор и репрессор могут инактивироваться или наоборот активироваться.







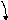

 Активация транскрипции
Активация транскрипции



 А ДНК
А ДНК

 Репрессор
Репрессор




 ДНК
ДНК



 — РНК-полимераза, — активатор, — репрессор
— РНК-полимераза, — активатор, — репрессор

 Промотор Оператор Ген
Промотор Оператор Ген




Рис.70. Влияние белков регуляторов (активатора и репрессора) на связывание РНК-полимераза с промотором и модификация транскрипции. А – активатор связался с оператором и активировал связывание РНК-полимеразы с промотором. Синтез РНК увеличился. Б – репрессор связался с оператором и частично с промотором препятствуя соединению РНК-полимеразы с промотором. Транскрипция не происходит.
а. В случае активации активатора транскрипция усилится, в случае активации репрессора транскрипция прекратиться .
б. В случае инактивации активатора транскрипция снизится, в случае инактивации репрессора транскрипция усилится.
Таким образом, субстрат и продукт могут выступать как активаторы транскрипции – тогда их называют индукторами (см. рис. 71 ) или как ингибиторы – тогда их называют корепрессоры (см. рис. 72).
Приведём пример регуляции экспрессии гена на примере кишечной палочки, утилизирующей из среды лактозу. Клетки E.coli в качестве источника углерода,








 Усиление транскрипции
Усиление транскрипции

 А
А





 Активатор
Активатор




 Индуктор
Индуктор





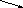
 Репрессор
Репрессор




 Инактивация
Инактивация
 Усиление транскрипции
Усиление транскрипции

Рис. 71. Влияние индуктора на активатор (активация) и репрессор (инактивация) и модификация транскрипции. Обозначения те же что и на рис .
обычно используют глюкозу, расщепляя её специальными ферментами. При её отсутствии и наличии в среде лактозы клетки через несколько минут начинают утилизировать и этот дисахарид.
Теория оперона объясняет это явление следующим образом. В кишечной палочке имеются гены способные утилизировать лактозу. В
её отсутствии нет необходимости для их синтеза, поэтому репрессор, связанный с оператором, блокирует движение РНК-полимеразы и транскрипция гена, контро-
лирующего синтез фермента расщепляющего лактозу, не осуществляется. При наличии в среде лактозы последняя взаимодействует с репрессором инактивируя его, в результате он теряет способность связываться с оператором. РНК-полимераза свободно проходит через оператор, транскрибируя иРНК, которая реализует информацию (в рибосомах) в фермент, расщепляющий лактозу.
РНК-полимераза с трудом
 осаждается на промоторе
осаждается на промоторе
 А Торможение транскрипции
А Торможение транскрипции


 ДНК
ДНК
И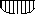



 нактивация
нактивация
а
 ктиватора
ктиватора
И

 нактивированный ре-
нактивированный ре-
п
 рессор активируется
рессор активируется







 Б ДНК
Б ДНК
Рис. 72. Влияние корепрессора на репрессор (активация) и индуктор (инактивация) и модификация транскрипции. Обозначения те же что и на рис. .
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Источник



