
- Регуляция каталитической активности ферментов путём фосфорилирования/дефосфорилирования
- Регуляция каталитической активности ферментов частичным (ограниченным) протеолизом
- Регуляция каталитической активности ферментов путём фосфорилирования/дефосфорилирования
- Регуляция каталитической активности ферментов частичным (ограниченным) протеолизом
Регуляция каталитической активности ферментов путём фосфорилирования/дефосфорилирования
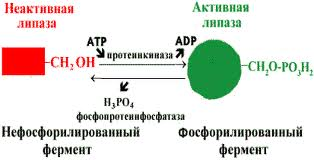
В биологических системах часто встречается механизм регуляции активности ферментов с помощью ковалентной модификации аминокислотных остатков. Быстрый и широко распространённый способ химической модификации ферментов — фосфорилирование/дефосфорилирование (ОН-группы фермента): фосфорилирование осуществляется ферментами протеинкиназами, дефосфорилирование — фосфопротеинфосфатазами. Присоединение остатка фосфорной кислоты приводит к изменению конформации активного центра и его каталитической активности. Результат: одни ферменты при фосфорилировании активируются, другие, напротив, становятся менее активными.
Изменение активности фермента, вызванное фосфорилированием, обратимо. Отщепление остатка фосфорной кислоты осуществляется ферментами фосфопротеинфосфатазами. Активность протеинкиназ и фосфопротеинфосфатаз регулируется гормонами, что позволяет быстро изменять активность ключевых ферментов метаболических путей в зависимости от условий внешней среды. Антагонистичные по функции гормоны противоположным образом влияют на фосфо-рилирование/дефосфорилирование ферментов, вызывая противоположные эффекты изменения метаболизма клетки.
Например, под действием глюкагона (в период между приёмами пищи) в клетках происходит уменьшение синтеза энергетического материала — жира, гликогена и усиление его распада (мобилизация), вызванного фосфорилированием ключевых ферментов этих процессов. А под действием инсулина (во время пищеварения), наоборот, активируется синтез гликогена и ингибируется его распад, так как взаимодействие инсулина с рецептором активирует сигнальный путь, приводящий к дефосфорилированию тех же ключевых ферментов.
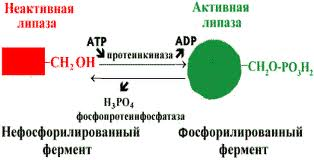
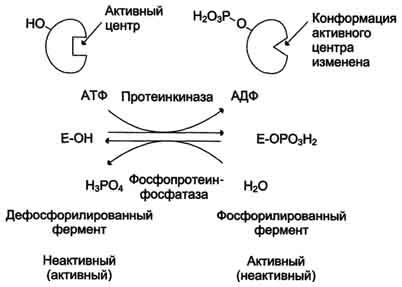
Регуляция активности ферментов фосфорилированием/дефосфорилированием.
Регуляция каталитической активности ферментов частичным (ограниченным) протеолизом
Некоторые ферменты, функционирующие вне клеток (в ЖКТ или в плазме крови), синтезируются в виде неактивных предшественников и активируются только в результате гидролиза одной или нескольких определённых пептидных связей, что приводит к отщеплению части белковой молекулы предшественника. В результате в оставшейся части белковой молекулы происходит конформационная перестройка и формируется активный центр фермента.
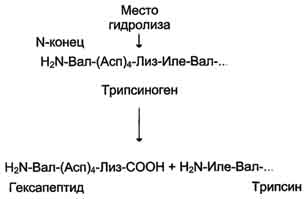
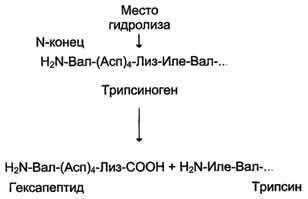
Трипсиноген, синтезируемый в поджелудочной железе, при пищеварении по протокам поджелудочной железы поступает в двенадцатиперстную кишку, где и активируется путём частичного протеолиза под действием фермента кишечника энтеропептидазы. В результате отщепления гексапептида с N-конца формируется активный центр в оставшейся части молекулы. Трипсин относят к семейству «сериновых» протеаз — активный центр фермента содержит функционально важный остаток Сер.
Частичный протеолиз — пример регуляции, когда активность фермента изменяется необратимо. Такие ферменты функционируют, как правило, в течение короткого времени, определяемого временем жизни белковой молекулы. Частичный протеолиз лежит в основе активации протеолитических ферментов, белков свёртывающей системы крови и фибринолиза, белков системы комплемента, а также пептидных гормонов.
Интенсивность протекания биохимических процессов определяется функционированием ферментных систем клетки.
Регуляция деятельности ферментного аппарата клетки осуществляется изменением каталитической активности ферментов — быстрый механизм регуляции, или
изменением концентрации ферментов — медленный механизм регуляции.
Различают следующие уровни регуляции биохимических процессов через ферментные системы клетки:
1.Низший — изменение каталитической активности ферментов вследствие изменения факторов среды: концентрация субстрата, концентрация кофакторов, концентрация продуктов реакции, рН среды, температура, присутствие ингибиторов или активаторов.
2.Изменение каталитической активности регуляторных ферментов.
3.Изменение интенсивности синтеза ферментов под влиянием метаболитов (в основном у прокариот).
4.Высший — изменение функционирования ферментного аппарата клетки под влиянием гормонов, которые или сами проникают в клетку, в ядро (стероидные и тиреоидные гормоны), или действуют на наружные рецепторы клетки (белково-пептидные гормоны и катехоламины).
МЕДИЦИНСКИЕ АСПЕКТЫ ЭНЗИМОЛОГИИ
Медицинской биохимии принадлежит существенная роль в процессе познания патогенетических механизмов. Еще два века назад М.В. Ломоносов писал, что «медик без довольного знания химии совершенно быть не может».
Основная задача медицинской биохимии заключается в выяснении компенсаторных механизмов нарушенных этапов метаболических процессов в клетке и способов управления этой компенсацией. Успешно развивается новое направление энзимологии — медицинская энзимология по трем главным направлениям: энзимопатологии, энзимодиагностики и энзимо-терапии.
Нарушения функционирования ферментов в клетке – энзимопатии:
Первичные энзимопатии: дефектные ферменты наследуются, в основном, по аутосомнорецессивному типу. Гетерозиготы, чаще всего, не имеют фенотипических отклонений. Первичные энзимопатии обычно относят к метаболическим болезням, так как происходит нарушение определённых метаболических путей. При этом развитие заболевания может протекать по одному из ниже перечисленных «сценариев». Условную схему метаболического пути:
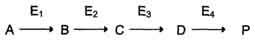
Нарушение образования конечных продуктов
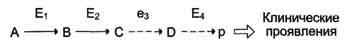
Клинические проявления. Альбинизм: нарушен синтез в меланоцитах пигментов — меланинов. Меланин находится в коже, волосах, радужке, пигментном эпителии сетчатки глаза и влияет на их окраску. При альбинизме наблюдают слабую пигментацию кожи, светлые волосы, красноватый цвет радужки глаза из-за просвечивающих капилляров. Проявление альбинизма связано с недостаточностью фермента тирозингидроксилазы (тирозиназы) — одного из ферментов, катализирующего метаболический путь образования меланинов.
Накопление субстратов-предшественников. При недостаточности фермента Е3 будут накапливаться вещество С, а также во многих случаях и предшествующие соединения. Увеличение субстратов-предшественников дефектного фермента — ведущее звено развития многих заболеваний:
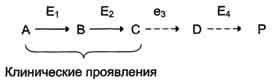
Клинические проявления. Алкаптонурия: нарушено окисление гомогентизиновой кислоты в тканях (гомогентизиновая кислота — промежуточный метаболит катаболизма тирозина). У таких больных наблюдают недостаточность фермента окисления гомогентизиновой кислоты — диоксигеназы гомогентизиновой кислоты, приводящей к развитию заболевания. В результате увеличиваются концентрация гомогентизиновой кислоты и выведение её с мочой. В присутствии кислорода гомогентизиновая кислота превращается в соединение чёрного цвета – алкаптон, поэтому моча на воздухе окрашивается в чёрный цвет. Алкаптон также образуется и в биологических жидкостях, оседая в тканях, коже, сухожилиях, суставах. При значительных отложениях алкаптона в суставах нарушается их подвижность.
Нарушение образования конечных продуктов и накопление субстратов предшественников. Отмечают заболевания, когда одновременно недостаток продукта и накопление исходного субстрата вызывают клинические проявления.
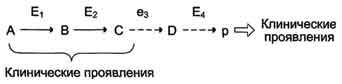
Клинические проявления. Болезнью Гирке (гликогеноз I типа): снижение концентрации глюкозы в крови (гипогликемия) в перерывах между приёмами пищи. Это связано с нарушением распада гликогена в печени и выходом из неё глюкозы вследствие дефекта фермента глюкозо-6-фосфатфосфатазы. Одновременно у таких людей увеличиваются размеры печени (гепатомегалия) вследствие накопления в ней не используемого гликогена.
Также, фенилкетонурия (фенилпировиноградная олигофрения) связана с дефектом фенилаланин-4-монооксигеназы (фенилаланингидролазе), которая гидроксилирует фенилаланин в тирозин (первый этап метаболического пути). В результате накапливаются аномальные метаболиты фенилаланина, оказывающие сильный токсический эффект на развитие нервной ткани, что приводит к умственной отсталости. Нормальное умственное развитие такого ребенка зависит от ранней диагностики и своевременного назначения специальной диеты.
Заболевание подагра связано с дефектом ферментов метаболизма пуриновых оснований и накоплением мочевой кислоты; галактоземия, недостаточность лактазы и сахаразы, различные липидозы.
По классификации А.А. Покровского к ферментопатиям относятся:
1) наследственные энзимопатии;
2) токсические энзимопатии;
3) алиментарные энзимопатии, вызванные нарушениями нейрогуморальных регуляций; и
4) группа различных патологических процессов, сопровождающихся нарушением и/или изменением ферментативных реакций.
Вторичные (приобретенные) энзимопатии возникают как следствие заболеваний органов, вирусных инфекций и т.п., что приводит к нарушению синтеза фермента или условий его работы: гипераммониемия при заболеваниях печени, при которых ухудшается синтез мочевины и в крови накапливается аммиак; недостаточность ферментов желудочно-кишечного тракта при заболеваниях желудка, поджелудочной железы или желчного пузыря; недостаток витаминов и их коферментных форм также является причиной приобретенных ферментопатий.
Источник
Регуляция каталитической активности ферментов путём фосфорилирования/дефосфорилирования
В биологических системах часто встречается механизм регуляции активности ферментов с помощью ковалентной модификации аминокислотных остатков. Быстрый и широко распространённый способ химической модификации ферментов — фосфорилирование/дефосфорилирование (ОН-группы фермента): фосфорилирование осуществляется ферментами протеинкиназами, дефосфорилирование — фосфопротеинфосфатазами. Присоединение остатка фосфорной кислоты приводит к изменению конформации активного центра и его каталитической активности. Результат: одни ферменты при фосфорилировании активируются, другие, напротив, становятся менее активными.
Изменение активности фермента, вызванное фосфорилированием, обратимо. Отщепление остатка фосфорной кислоты осуществляется ферментами фосфопротеинфосфатазами. Активность протеинкиназ и фосфопротеинфосфатаз регулируется гормонами, что позволяет быстро изменять активность ключевых ферментов метаболических путей в зависимости от условий внешней среды. Антагонистичные по функции гормоны противоположным образом влияют на фосфо-рилирование/дефосфорилирование ферментов, вызывая противоположные эффекты изменения метаболизма клетки.
Например, под действием глюкагона (в период между приёмами пищи) в клетках происходит уменьшение синтеза энергетического материала — жира, гликогена и усиление его распада (мобилизация), вызванного фосфорилированием ключевых ферментов этих процессов. А под действием инсулина (во время пищеварения), наоборот, активируется синтез гликогена и ингибируется его распад, так как взаимодействие инсулина с рецептором активирует сигнальный путь, приводящий к дефосфорилированию тех же ключевых ферментов.

Регуляция активности ферментов фосфорилированием/дефосфорилированием.
Регуляция каталитической активности ферментов частичным (ограниченным) протеолизом
Некоторые ферменты, функционирующие вне клеток (в ЖКТ или в плазме крови), синтезируются в виде неактивных предшественников и активируются только в результате гидролиза одной или нескольких определённых пептидных связей, что приводит к отщеплению части белковой молекулы предшественника. В результате в оставшейся части белковой молекулы происходит конформационная перестройка и формируется активный центр фермента.
Трипсиноген, синтезируемый в поджелудочной железе, при пищеварении по протокам поджелудочной железы поступает в двенадцатиперстную кишку, где и активируется путём частичного протеолиза под действием фермента кишечника энтеропептидазы. В результате отщепления гексапептида с N-конца формируется активный центр в оставшейся части молекулы. Трипсин относят к семейству «сериновых» протеаз — активный центр фермента содержит функционально важный остаток Сер.
Частичный протеолиз — пример регуляции, когда активность фермента изменяется необратимо. Такие ферменты функционируют, как правило, в течение короткого времени, определяемого временем жизни белковой молекулы. Частичный протеолиз лежит в основе активации протеолитических ферментов, белков свёртывающей системы крови и фибринолиза, белков системы комплемента, а также пептидных гормонов.
Интенсивность протекания биохимических процессов определяется функционированием ферментных систем клетки.
Регуляция деятельности ферментного аппарата клетки осуществляется изменением каталитической активности ферментов — быстрый механизм регуляции, или
изменением концентрации ферментов — медленный механизм регуляции.
Различают следующие уровни регуляции биохимических процессов через ферментные системы клетки:
1.Низший — изменение каталитической активности ферментов вследствие изменения факторов среды: концентрация субстрата, концентрация кофакторов, концентрация продуктов реакции, рН среды, температура, присутствие ингибиторов или активаторов.
2.Изменение каталитической активности регуляторных ферментов.
3.Изменение интенсивности синтеза ферментов под влиянием метаболитов (в основном у прокариот).
4.Высший — изменение функционирования ферментного аппарата клетки под влиянием гормонов, которые или сами проникают в клетку, в ядро (стероидные и тиреоидные гормоны), или действуют на наружные рецепторы клетки (белково-пептидные гормоны и катехоламины).
МЕДИЦИНСКИЕ АСПЕКТЫ ЭНЗИМОЛОГИИ
Медицинской биохимии принадлежит существенная роль в процессе познания патогенетических механизмов. Еще два века назад М.В. Ломоносов писал, что «медик без довольного знания химии совершенно быть не может».
Основная задача медицинской биохимии заключается в выяснении компенсаторных механизмов нарушенных этапов метаболических процессов в клетке и способов управления этой компенсацией. Успешно развивается новое направление энзимологии — медицинская энзимология по трем главным направлениям: энзимопатологии, энзимодиагностики и энзимотерапии.
Нарушения функционирования ферментов в клетке – энзимопатии:
Первичные энзимопатии: дефектные ферменты наследуются, в основном, по аутосомнорецессивному типу. Гетерозиготы, чаще всего, не имеют фенотипических отклонений. Первичные энзимопатии обычно относят к метаболическим болезням, так как происходит нарушение определённых метаболических путей. При этом развитие заболевания может протекать по одному из ниже перечисленных «сценариев». Условную схему метаболического пути:
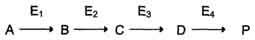
Нарушение образования конечных продуктов
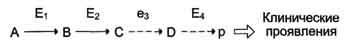
Клинические проявления. Альбинизм: нарушен синтез в меланоцитах пигментов — меланинов. Меланин находится в коже, волосах, радужке, пигментном эпителии сетчатки глаза и влияет на их окраску. При альбинизме наблюдают слабую пигментацию кожи, светлые волосы, красноватый цвет радужки глаза из-за просвечивающих капилляров. Проявление альбинизма связано с недостаточностью фермента тирозингидроксилазы (тирозиназы) — одного из ферментов, катализирующего метаболический путь образования меланинов.
Накопление субстратов-предшественников. При недостаточности фермента Е3 будут накапливаться вещество С, а также во многих случаях и предшествующие соединения. Увеличение субстратов-предшественников дефектного фермента — ведущее звено развития многих заболеваний:
Клинические проявления. Алкаптонурия: нарушено окисление гомогентизиновой кислоты в тканях (гомогентизиновая кислота — промежуточный метаболит катаболизма тирозина). У таких больных наблюдают недостаточность фермента окисления гомогентизиновой кислоты — диоксигеназы гомогентизиновой кислоты, приводящей к развитию заболевания. В результате увеличиваются концентрация гомогентизиновой кислоты и выведение её с мочой. В присутствии кислорода гомогентизиновая кислота превращается в соединение чёрного цвета – алкаптон, поэтому моча на воздухе окрашивается в чёрный цвет. Алкаптон также образуется и в биологических жидкостях, оседая в тканях, коже, сухожилиях, суставах. При значительных отложениях алкаптона в суставах нарушается их подвижность.
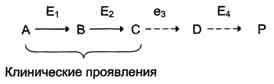
Нарушение образования конечных продуктов и накопление субстратов предшественников. Отмечают заболевания, когда одновременно недостаток продукта и накопление исходного субстрата вызывают клинические проявления.

Клинические проявления. Болезнью Гирке (гликогеноз I типа): снижение концентрации глюкозы в крови (гипогликемия) в перерывах между приёмами пищи. Это связано с нарушением распада гликогена в печени и выходом из неё глюкозы вследствие дефекта фермента глюкозо-6-фосфатфосфатазы. Одновременно у таких людей увеличиваются размеры печени (гепатомегалия) вследствие накопления в ней не используемого гликогена.
Также, фенилкетонурия (фенилпировиноградная олигофрения) связана с дефектом фенилаланин-4-монооксигеназы (фенилаланингидролазе), которая гидроксилирует фенилаланин в тирозин (первый этап метаболического пути). В результате накапливаются аномальные метаболиты фенилаланина, оказывающие сильный токсический эффект на развитие нервной ткани, что приводит к умственной отсталости. Нормальное умственное развитие такого ребенка зависит от ранней диагностики и своевременного назначения специальной диеты.
Заболевание подагра связано с дефектом ферментов метаболизма пуриновых оснований и накоплением мочевой кислоты; галактоземия, недостаточность лактазы и сахаразы, различные липидозы.
По классификации А.А. Покровского к ферментопатиям относятся:
1) наследственные энзимопатии;
2) токсические энзимопатии;
3) алиментарные энзимопатии, вызванные нарушениями нейрогуморальных регуляций; и
4) группа различных патологических процессов, сопровождающихся нарушением и/или изменением ферментативных реакций.
Вторичные (приобретенные) энзимопатии возникают как следствие заболеваний органов, вирусных инфекций и т.п., что приводит к нарушению синтеза фермента или условий его работы: гипераммониемия при заболеваниях печени, при которых ухудшается синтез мочевины и в крови накапливается аммиак; недостаточность ферментов желудочно-кишечного тракта при заболеваниях желудка, поджелудочной железы или желчного пузыря; недостаток витаминов и их коферментных форм также является причиной приобретенных ферментопатий.
Источник



