
Транспирация и ее регулирование растением
Биологическое значение и размеры транспирации
В основе расходования воды растительным организмом лежит физический процесс испарения — переход воды из жидкого состояния в парообразное, происходящий в результате соприкосновения органов растения с ненасыщенной водой атмосферой. Расходование воды растением регулируется целым рядом анатомических и физиологических механизмов. Термин «транспирация» введен для того, чтобы подчеркнуть отличие этого физиологического процесса от чисто физического испарения.
Биологическое значение транспирации состоит в терморегуляции растения, в обеспечении деятельности верхнего концевого двигателя водного тока, при помощи которого осущесталяется и поступление различных веществ, а также в регулировании насыщенности клеток водой, благодаря чему создаются оптимальные условия для процессов жизнедеятельности.
Одной из важных характеристик процесса является интенсивность транспирации — количество воды, испаряемое растением с единицы листовой поверхности в единицу времени. В некоторых случаях удобнее проводить расчет не на единицу площади, а на единицу массы листьев. Для большинства сельскохозяйственных растений интенсивность транспирации составляет днем 15-250, а ночью 1-20 г/м 2 ·ч.
Поэтому небольшой перепад относительном влажности приводит к значительной депрессии его водного потенциала. Так, при относительной влажности воздуха 100 % водный потенциал равен нулю, при влажности 99,6 — 0,5 МПа, при 99 и 97 % он составляет соответственно -1,36 и -4,0 МПа. Относительная влажность воздуха летом наиболее часто не превышает 60 %, водный потенциал при этом снижается до -68 МПа, а во время суховея (влажность 30 %) депрессия водного потенциала достигает 200 МПа.
Кутикулярная транспирация. С самого начала существования наземных растений возникла дилемма: ассимиляция СО2 из атмосферы требует интенсивного газообмена; предотвращение значительной потери воды возможно только при наличии хорошей изоляции от окружающего воздуха, имеющего крайне низкие значения водного потенциала. Главная проблема газообмена, как ее сформулировал О. Штоккер (1923), состоит «в лавировании между жаждой и голодом».
Поддержание водного гомеостаза листа достигается наличием покровной ткани — эпидермиса, представляющей слой довольно сильно примыкающих друг к другу клеток, наружные стенки которых утолщены. Снаружи эпидермис покрыт кутикулой, в состав которой входит кутин — полимерные эфиры оксимонокарбононовых кислот, содержащих по 16-28 атомов углерода и по 2-3 гидроксильные группы. В состав кутикулы входят также пластинки воска. От плазмалеммы эпидермальных клеток к кутикуле отходят канальцы, заполненные рыхлой сетью из целлюлозных фибрилл. Они служат полярными путями поглощения и экскреции веществ листом.
Кутикулярное диффузионное сопротивление в большинстве случаев очень велико. Оно зависит от толщины кутикулы, расположения, плотности и числа прослоек кутана и воска. На формировании изолирующих структур на поверхности листьев в сильной степени сказываются видовые особенности и возраст растений, а также условия их произрастания. При атмосферной и почвенной засухе взрослые листья обладают обычно более толстой кутикулой и более плотным восковым налетом. Кутикулярная защита от транспирации весьма эффективна. У взрослых листьев кутикулярная транспирация составляет 10-20 % общего испарения воды. Например, у семечковых плодовых деревьев она измеряется 120-160 мг/дм 2 ч.
У кутикулы есть уникальное свойство, обусловленное особенностями ее состава — изменять гидравлическую проводимость в зависимости от оводненности. При подсыхании наружных слоев стенок эпидермиса гидрофобные слои кутикулы плотнее придвигаются друг к другу, поэтому кутикулярное сопротивление межет удвоиться. При низких температурах оно также возрастает. И, наоборот, при увеличении оводненности эпидермиса кутикула набухает за счет гидратации карбоксильных и оксигрупп, разрыхляется, кутикулярное сопротивление диффузии значительно снижается и транспирация возрастает. Таким образом, потеря воды через кутикулу регулируется оводненностью листа. По ночам, например, при более сильном набухании кутикулы кутикулярная транспирация идет интенсивнее, чем днем. Смоченные листья могут поглощать воду через кутикулу.
Устьютная транспирация. Основной путь сообщения мезофилла листа с атмосферой — устьица. Процесс устьичной транспирации можно разделить на ряд этапов (См. рисунок! ).
П е р в ы и э т а п — испарение воды с поверхности клеток в межклетники. Каждая клетка мезофилла хотя бы одной своей стороной граничите межклеточным пространством. Поверхность всех клеточных стенок, соприкасающихся с межклетниками превышает поверхность листа примерно в 10-30 раз. Необходимо отметить, что уже на этом этапе растение способно регулировать транспирацию. Уменьшение испарения воды в межклетники достигается по крайней мере двумя механизмами. Первый обусловлен изменениями водоудерживающей способности цитоплазмы путем увеличения осмотического и коллоидного связывания воды, ее компартментации в отдельных органеллах клетки и снижения проницаемости мембран. Второй механизм связан с уменьшением оводненности клеточных стенок. Он получил название механизма начинающегося подсыхания, или корневого регулирования транспирации.
При снижении подачи воды корнем и увеличении водоудерживающей способНасти цитоплазмы клеток мезофилла оболочки клеток оказываются менее насыщенными водой, водные мениски в капиллярах между фибриллами становятся вогнутыми, что увеличивает силы поверхностного натяжения и затрудняет переход воды в парообразное состояние. Поэтому при открытых устьицах происходит снижение транспирации за счет уменьшения количества водяного пара в межклетниках. Это внеустьичный способ регулирования транспирации, который представляет несомненную выгоду для в листе растениям так как позволяет снижать расхад воды без ущерба для ассимиляции углекислого газа. У хлопчатника преобладает именно этот способ регулирования транспирации. У растений умеренной зоны внеустьичное регулирование выражено в меньшей степени: транспирация может быть снижена на 30 % без закрывания устьиц. Возможности внеустьичного регулирования в значительной степени зависят от условий выращивания и возраста растений. Например, оптимизация минерального питания увеличивает водоудерживающую способность тканей, при старении листьев она снижается.
В т о р о й э т а п — выход паров воды из межклетников через устьичные щели. Число устьиц и их размещение сильно варьируют у разных видов растений. У ксерофитов — растений засушливых мест обитания — на единицу площади обычно приходится меньшее количество устьиц, чем у мезофитов. Кроме того, устьица у них часто углублены в сильно кутинизированную поверхность листьев, что уменьшает потерю воды. У большинства сельскохозяйственных культур устьица в основном расположены с нижней стороны листа (табл. ). Это одно из приспособлений для меньшей затраты воды.
Количество устьиц у сельскохозяйственных культур, шт/см 2
Источник
Транспирация и ее регулирование растением
Биологическое значение и размеры транспирации
В основе расходования воды растительным организмом лежит физический процесс испарения — переход воды из жидкого состояния в парообразное, происходящий в результате соприкосновения органов растения с ненасыщенной водой атмосферой. Расходование воды растением регулируется целым рядом анатомических и физиологических механизмов. Термин «транспирация» введен для того, чтобы подчеркнуть отличие этого физиологического процесса от чисто физического испарения.
Биологическое значение транспирации состоит в терморегуляции растения, в обеспечении деятельности верхнего концевого двигателя водного тока, при помощи которого осущесталяется и поступление различных веществ, а также в регулировании насыщенности клеток водой, благодаря чему создаются оптимальные условия для процессов жизнедеятельности.
Одной из важных характеристик процесса является интенсивность транспирации — количество воды, испаряемое растением с единицы листовой поверхности в единицу времени. В некоторых случаях удобнее проводить расчет не на единицу площади, а на единицу массы листьев. Для большинства сельскохозяйственных растений интенсивность транспирации составляет днем 15-250, а ночью 1-20 г/м 2 ·ч.
Поэтому небольшой перепад относительном влажности приводит к значительной депрессии его водного потенциала. Так, при относительной влажности воздуха 100 % водный потенциал равен нулю, при влажности 99,6 — 0,5 МПа, при 99 и 97 % он составляет соответственно -1,36 и -4,0 МПа. Относительная влажность воздуха летом наиболее часто не превышает 60 %, водный потенциал при этом снижается до -68 МПа, а во время суховея (влажность 30 %) депрессия водного потенциала достигает 200 МПа.
Кутикулярная транспирация. С самого начала существования наземных растений возникла дилемма: ассимиляция СО2 из атмосферы требует интенсивного газообмена; предотвращение значительной потери воды возможно только при наличии хорошей изоляции от окружающего воздуха, имеющего крайне низкие значения водного потенциала. Главная проблема газообмена, как ее сформулировал О. Штоккер (1923), состоит «в лавировании между жаждой и голодом».
Поддержание водного гомеостаза листа достигается наличием покровной ткани — эпидермиса, представляющей слой довольно сильно примыкающих друг к другу клеток, наружные стенки которых утолщены. Снаружи эпидермис покрыт кутикулой, в состав которой входит кутин — полимерные эфиры оксимонокарбононовых кислот, содержащих по 16-28 атомов углерода и по 2-3 гидроксильные группы. В состав кутикулы входят также пластинки воска. От плазмалеммы эпидермальных клеток к кутикуле отходят канальцы, заполненные рыхлой сетью из целлюлозных фибрилл. Они служат полярными путями поглощения и экскреции веществ листом.
Кутикулярное диффузионное сопротивление в большинстве случаев очень велико. Оно зависит от толщины кутикулы, расположения, плотности и числа прослоек кутана и воска. На формировании изолирующих структур на поверхности листьев в сильной степени сказываются видовые особенности и возраст растений, а также условия их произрастания. При атмосферной и почвенной засухе взрослые листья обладают обычно более толстой кутикулой и более плотным восковым налетом. Кутикулярная защита от транспирации весьма эффективна. У взрослых листьев кутикулярная транспирация составляет 10-20 % общего испарения воды. Например, у семечковых плодовых деревьев она измеряется 120-160 мг/дм 2 ч.
У кутикулы есть уникальное свойство, обусловленное особенностями ее состава — изменять гидравлическую проводимость в зависимости от оводненности. При подсыхании наружных слоев стенок эпидермиса гидрофобные слои кутикулы плотнее придвигаются друг к другу, поэтому кутикулярное сопротивление межет удвоиться. При низких температурах оно также возрастает. И, наоборот, при увеличении оводненности эпидермиса кутикула набухает за счет гидратации карбоксильных и оксигрупп, разрыхляется, кутикулярное сопротивление диффузии значительно снижается и транспирация возрастает. Таким образом, потеря воды через кутикулу регулируется оводненностью листа. По ночам, например, при более сильном набухании кутикулы кутикулярная транспирация идет интенсивнее, чем днем. Смоченные листья могут поглощать воду через кутикулу.
Устьютная транспирация. Основной путь сообщения мезофилла листа с атмосферой — устьица. Процесс устьичной транспирации можно разделить на ряд этапов (См. рисунок! ).
П е р в ы и э т а п — испарение воды с поверхности клеток в межклетники. Каждая клетка мезофилла хотя бы одной своей стороной граничите межклеточным пространством. Поверхность всех клеточных стенок, соприкасающихся с межклетниками превышает поверхность листа примерно в 10-30 раз. Необходимо отметить, что уже на этом этапе растение способно регулировать транспирацию. Уменьшение испарения воды в межклетники достигается по крайней мере двумя механизмами. Первый обусловлен изменениями водоудерживающей способности цитоплазмы путем увеличения осмотического и коллоидного связывания воды, ее компартментации в отдельных органеллах клетки и снижения проницаемости мембран. Второй механизм связан с уменьшением оводненности клеточных стенок. Он получил название механизма начинающегося подсыхания, или корневого регулирования транспирации.
При снижении подачи воды корнем и увеличении водоудерживающей способНасти цитоплазмы клеток мезофилла оболочки клеток оказываются менее насыщенными водой, водные мениски в капиллярах между фибриллами становятся вогнутыми, что увеличивает силы поверхностного натяжения и затрудняет переход воды в парообразное состояние. Поэтому при открытых устьицах происходит снижение транспирации за счет уменьшения количества водяного пара в межклетниках. Это внеустьичный способ регулирования транспирации, который представляет несомненную выгоду для в листе растениям так как позволяет снижать расхад воды без ущерба для ассимиляции углекислого газа. У хлопчатника преобладает именно этот способ регулирования транспирации. У растений умеренной зоны внеустьичное регулирование выражено в меньшей степени: транспирация может быть снижена на 30 % без закрывания устьиц. Возможности внеустьичного регулирования в значительной степени зависят от условий выращивания и возраста растений. Например, оптимизация минерального питания увеличивает водоудерживающую способность тканей, при старении листьев она снижается.
В т о р о й э т а п — выход паров воды из межклетников через устьичные щели. Число устьиц и их размещение сильно варьируют у разных видов растений. У ксерофитов — растений засушливых мест обитания — на единицу площади обычно приходится меньшее количество устьиц, чем у мезофитов. Кроме того, устьица у них часто углублены в сильно кутинизированную поверхность листьев, что уменьшает потерю воды. У большинства сельскохозяйственных культур устьица в основном расположены с нижней стороны листа (табл. ). Это одно из приспособлений для меньшей затраты воды.
Количество устьиц у сельскохозяйственных культур, шт/см 2
Источник
Транспирация и ее регулирование растением
ЛЕКЦИЯ 17
Основные закономерности водопотребления растений.
(Транспирация. Дыхание растений)
В основе расходования воды растительным организмом лежит физический процесс испарения – переход воды из жидкого состояния в парообразное, происходящий в результате соприкосновения органов растения с ненасыщенной водой атмосферой. Расходование воды растением регулируется целым рядом анатомических и физиологических механизмов. Термин «транспирация» введен для того, чтобы подчеркнуть отличие этого физиологического процесса от чисто физического испарения.
Одной из важных характеристик процесса является интенсивность транспирации – количество воды, испаряемое растением с единицы листовой поверхности в единицу времени. В некоторых случаях удобнее проводить расчет на единицу массы листьев. Для большинства сельскохозяйственных растений интенсивность транспирации составляет днем 15-250, а ночью 1-20 г/(м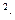 ч).
ч).
Высокая интенсивность транспирации, которую К.А.Тимирязев называл «необходимым злом», обусловлена тем, что атмосфера характеризуется очень низкими значениями водного потенциала. Водный потенциал связан логарифмической зависимостью с относительной влажностью воздуха:
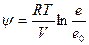 ,
,
Где R- газовая постоянная; T- абсолютная температура; V- парциальный мольный объем; e – давление водяного пара в воздухе; e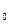 — давление водяного пара, насыщающего воздух при данной температуре.
— давление водяного пара, насыщающего воздух при данной температуре.
Поэтому небольшой перепад относительной влажности приводит к значительной депрессии его водного потенциала. Так, при относительной влажности воздуха 100% водный потенциал равен нулю, при влажности 99,6% — 0,5 МПа, при 99 и 97% он составляет соответственно – 1,36 и — 4,0 МПа. Относительная влажность воздуха летом наиболее часто не превышает 60%, водный потенциал при этом снижается до – 68 МПа, а во время суховея (влажность 30%) депрессия водного потенциала достигает 200 МПа.
Кутикулярная транспирация. С самого существования наземных растений возникла дилемма: ассимиляция СО 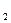 из атмосферы требует интенсивного газообмена; предотвращение значительной потери воды возможно только при наличии хорошей изоляции от окружающего воздуха, имеющего крайне низкие значения водного потенциала. Главная проблема газообмена, как ее сформулировал О.Штоккер (1923), состоит в «лавировании между жаждой и голодом»
из атмосферы требует интенсивного газообмена; предотвращение значительной потери воды возможно только при наличии хорошей изоляции от окружающего воздуха, имеющего крайне низкие значения водного потенциала. Главная проблема газообмена, как ее сформулировал О.Штоккер (1923), состоит в «лавировании между жаждой и голодом»
Поддержание водного гомеостаза листа достигается наличием покровной ткани – эпидермиса.
Снаружи эпидермис покрыт кутикулой, в состав которой входят кутин – полимерные эфиры оксимонокарбоновых кислот и пластинки воска.
Кутикулярное диффузное сопротивление в большинстве случаев очень велико. Оно зависит от толщины кутикулы, расположения, плотности и числа прослоек Кутина и воска. Кутикулярная защита от транспирации весьма эффективна. У взрослых листьев кутикулярная транспирация составляет 10-20% общего испарения воды.
У кутикулы есть уникальное свойство, обусловленное особенностями ее состава, — изменять гидравлическую проводимость в зависимости от оводненности. Таким образом, потеря воды через кутикулу регулируется оводненностью листа. По ночам, например, при более сильном набухании кутикулы кутикулярная транспирация идет интенсивнее, чем днем. Смоченные листья могут поглощать воду через кутикулу.
Устьичная транспирация.Основной путь сообщения мезофилла листа с атмосферой – устьица. Процесс устьичной транспирации можно разделить на несколько этапов.
Первый этап – испарение воды с поверхности клеток в межклетники. Каждая клетка мезофилла хотя бы одной своей стороной граничит с межклеточным пространством. Необходимо отметить, что уже на этом этапе растение способно регулировать транспирацию. Уменьшение испарения достигается двумя механизмами. Первый обусловлен изменениями водоудерживающей способности цитоплазмы путем увеличения осмотического и коллоидного связывания воды, ее компартментации в отдельных органеллах клетки и снижения проницаемости мембран. Второй механизм связан с уменьшением оводненности клеточных стенок.
При снижении подачи воды корнем и увеличении водоудерживающей способности цитоплазмы клеток мезофилла клеточные стенки оказываются менее насыщенными водой, водные мениски в капиллярах между фибриллами становятся вогнутыми, что увеличивает силы поверхностного натяжения и затрудняет переход воды в парообразное состояние. Поэтому при открытых устьицах происходит снижение транспирации за счет уменьшения количества водяного пара в межклетниках. Это внеустьичный способ регулирования транспирации, который представляет несомненную выгоду для растения, так как позволяет снижать расход воды без ущерба для ассимиляции диоксида углерода.
Второй этап – выход паров воды из межклетников через устьичные щели. Число устьиц и их размещение сильно варьируют у разных видов растений. У большинства сельскохозяйственных растений устьица расположены в основном с нижней стороны листа. Это одно из приспособлений для снижения расходования воды.
Обычно устьица занимают 1-3% всей поверхности листа. Однако относительная транспирация, под которой понимают отношение испарения воды листом к испарению с такой же по величине свободной поверхности, составляет 0,5- 0,8 и может приближаться к единице. Высокая скорость диффузии через устьица объясняется тем, что испарение из ряда мелких отверстий происходит быстрее, чем из одного крупного той же площади. Это связано с повышенной краевой диффузией.
При открытых устьицах испарение может быть таким же, как с открытой водной поверхности. Закрывание устьиц наполовину еще мало влияет на интенсивность транспирации. Полное закрывание устьиц сокращает транспирацию примерно на 90% . Таким образом, изменение степени открытости устьиц – устьичная регулировка – является основным механизмом контроля транспирации растением.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Источник



