
Научная электронная библиотека

§ 3.1.4. Строение клетки
Размеры клетки широко варьируют от 0,1 мкм (некоторые бактерии) до 155 мм (яйцо страуса). У всех клеток, независимо от их формы, размеров, функциональной нагрузки обнаруживается сходное строение (рис. 3.13).
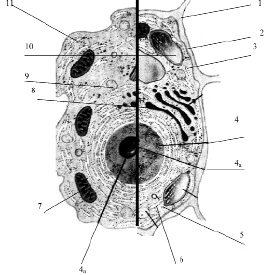
Рис. 3.13. Схема строения живой клетки: 1 – оболочка; 2 – мембрана; 3 – цитоплазма; 4 – ядро; 4а – ядрышко; 5 – рибосомы; 6 – эндоплазматическая сеть (ЭПС); 7 – митохондрии; 8 – комплекс гольджи; 9 – лизосомы; 10 – пластиды; 11 – клеточные включения
Снаружи клетка одета мембраной. Внутренняя часть клетки содержит многочисленные органоиды – структурные образования клетки, выполняющие определенные функции жизнедеятельности клетки.
1. Оболочка. Присутствует только у растительных клеток. Состоит из волокон целлюлозы. Функции оболочки: защита клетки от внешних повреждений, придает стабильную форму клетки, эластичность растительным тканям.
Повреждение наружной оболочки приводит к гибели клетки (цитолиз).
2. Мембрана. Тончайшая структура (75 Ǻ), состоит из двойного слоя молекул липидов и одного слоя белков. Такая структура обеспечивает уникальную эластичность и прочность мембране
 участие в обмене веществ. Эта функция связана с избирательной проницаемостью в клетку определенных веществ и выведение из нее продуктов обмена. В процессе питания в клетку могут проникать определенные растворы веществ (пиноцитоз) и твердые частицы (фагоцитоз).
участие в обмене веществ. Эта функция связана с избирательной проницаемостью в клетку определенных веществ и выведение из нее продуктов обмена. В процессе питания в клетку могут проникать определенные растворы веществ (пиноцитоз) и твердые частицы (фагоцитоз).
Явление фагоцитоза – поглощение клеткой твердых частиц – впервые было описано русским врачом Мечниковым. Фагоцитарная особенность лежит в основе процесса иммунитета. Особенно развита у лейкоцитов, клеток костного мозга, лимфатических узлов, селезенки, надпочечников и гипофиза.
Пиноцитоз – поглощение клеткой растворов – состоит в том, что мельчайшие пузырьки жидкости втягиваются через образующуюся воронку, проникают через мембрану и усваиваются клеткой.
3. Цитоплазма – внутренняя среда клетки. Представляет собой гелеобразную жидкость (коллоидная система), состоит на 80 % из воды, в которой растворены белки, липиды, углеводы, неорганические вещества. Цитоплазма живой клетки находится в постоянном движении (циклоз).
 транспортировка питательных веществ и утилизация продуктов обмена клетки;
транспортировка питательных веществ и утилизация продуктов обмена клетки;
 буферность цитоплазмы (постоянство физико-химических свойств) обеспечивает гомеостаз клетки, поддерживает постоянные нужные параметры жизнедеятельности;
буферность цитоплазмы (постоянство физико-химических свойств) обеспечивает гомеостаз клетки, поддерживает постоянные нужные параметры жизнедеятельности;
 поддержание тургора (упругость) клетки;
поддержание тургора (упругость) клетки;
 все биохимические реакции происходят только в водных растворах, что обеспечивается в среде цитоплазмы.
все биохимические реакции происходят только в водных растворах, что обеспечивается в среде цитоплазмы.
4. Ядро – обязательный органоид эукариотических клеток. Впервые было исследовано и описано Р. Броуном в 1831 г. В молодых клетках расположено в центре клетки, в старых – смещается в сторону. Снаружи ядро окружено мембраной с крупными порами, способными пропускать крупные макромолекулы. Внутри ядро заполнено клеточным соком – кариоплазмой, основная часть ядра заполнена хроматином – ядерным веществом, содержащим ДНК и белок. Перед делением хроматин образует палочковидные хромосомы. Причём, хромосомы одинакового строения (но содержащие разные ДНК!) образуют пары, зрительно воспринимаемые как одно целое (рис. 3.14).

Рис. 3.14. Хромосомный набор человеческой клетки перед началом деления
Структурирование всех хромосом в пары свидетельствует о том, что число хромосом – чётное. Поэтому, его часто обозначают 2n, где n – количество хромосомных пар, а соответствующий набор хромосом называют диплоидным. Например, у голубей n = 40 (80 хромосом), у мухи n = 6 (12 хромосом), у собаки n = 39 (78 хромосом), у аскариды n = 1 (2 хромосомы). У человека n = 23 (46 хромосом). Однако, в половых клетках число хромосом в два раза меньше. Поэтому набор хромосом в половых клетках называется гаплоидным. Клетки, не являющиеся половыми называются соматическими. Иногда клетки с гаплоидным набором хромосом называют гаплоидными клетками, а с диплоидным набором хромосом – диплоидными клетками.
При слиянии двух родительских гаплоидных половых клеток образуется диплоидная клетка, дающая начало новому организму с набором генов отца и матери
Совокупность всех хромосом ядра (а значит и генов) клетки называется генотип. Именно генотип определяет все внешние и внутренние признаки конкретного организма.
В соматических клетках 44 Х-образные хромосомы (22 пары) у женщин и мужчин идентичны (сходны по строению), их называют аутосомами. А 23-я пара имеет конфигурацию ХХ – у женщин и ХY – у мужчин. Эти пары хромосом именуются половыми хромосомами.
В половых клетках 22 хромосомы также одинаковые у яйцеклеток и у сперматозоидов, а 23-я хромосома конфигурации Х – у яйцеклетки и Х или Y – у сперматозоидов. Поэтому при слиянии половых клеток и образовании пар хромосом, 23-я пара будет ( <ХY>или <ХХ>) определять пол будущего ребенка.
Необходимо помнить, что хотя в соматических клетках набор хромосом диплоидный (2n), однако, перед началом деления клеток происходит репликация ДНК, то есть, удвоение их количества, а, значит, и удвоение
количества хромосом. Поэтому перед началом деления соматической клетки в ней насчитывается 4n хромосом (рис. 16). Она становится тетраплоидной.
– хранение генетической информации;
– контроль за всеми процессами, происходящими в клетке: делением, дыханием, питанием и др.
4а. Ядрышко – структура, содержащаяся в ядре. Ядро может содержат 1, 2 или более ядрышек. Функция ядрышка – формирование рибосом.
Следует отметить, что не все клетки имеют оформленное ядро. Клетки, имеющие ядро называются эукариотическими или эукариотами. Клетки, не имеющие ядра, называются прокариотическими или прокариотами. Функции ядра у прокариот несёт одна нить ДНК (именуется хромосома), в которой хранится вся генетическая информация. К прокариотам относятся бактерии и сине-зеленые водоросли. Как правило, у прокариотов отсутствуют и некоторые другие органоиды. Размеры прокариотических клеток меньше, чем размеры эукариот.
5. Рибосомы – самые мелкие органоиды клетки. Были обнаружены в 1954 г. Французским ученым Паладом. Рибосомы были обнаружены в цитоплазме, а также на гранулярной ЭПС и в ядре.
Функция рибосом: обеспечение биосинтеза белка.
6. Эндоплазматическая сеть. Представляет собой каналы и полости, ограниченные мембраной. Различают две разновидности ЭПС: гранулярная ЭПС и агранулярная ЭПС. Гранулярная ЭПС морфологически отличается от агранулярной наличием на ее поверхности многочисленных рибосом (на агранулярной ЭПС рибосомы отсутствуют).
Функции эндоплазматической сети:
– участие в синтезе органических веществ: на гранулярной ЭПС синтезируются белки, на агранулярной – липиды и углеводы;
– транспортировка продуктов синтеза ко всем частям клетки.
Несложно уяснить, что гранулярная ЭПС характерна для клеток, синтезирующих белки (например клетки желез внутренней секреции), агранулярная ЭПС характерна для клеток-производителей углеводов и липидов (например клетки жировой ткани).
7. Митохондрии – крупные органоиды, состоящие из двойного слоя мембран: наружная – гладкая, внутренняя образует многочисленные гребнеобразные складки – кристы. Внутри митохондрии заполнены жидкостью (матрикс).
Функции митохондрий: основная функция митохондрий – обеспечение клетки энергией. Этот процесс происходит за счет синтеза аденозинтрифосфорной кислоты (АТФ) (рис. 3.15), в которой фрагмент

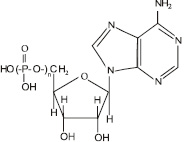
Рис. 3.15. Структурная формула аденозинфосфорных кислот. Для аденозинтрифосфорной кислоты n = 3, для аденозиндифосфорной кислоты n = 2, для аденозинмонофосфорной кислоты n = 1
При взаимодействии молекулы аденозинтрифосфорной кислоты с водой отщепляется один остаток фосфорной кислоты, в результате чего образуется аденозиндифосфорная кислота – АДФ и выделяется огромное количество энергии:
АТФ + Н2О = АДФ + Н3РО4 + 10 000 калорий.
Впоследствии от АДФ может отщепляться еще один остаток фосфорной кислоты, образуя АМФ – аденозинмонофосфорную кислоту.
АДФ + Н2О = АМФ + Н3РО4 + 10 000 калорий[37].
Освободившаяся энергия используется для жизнедеятельности клетки (КПД процесса превышает 80 %!).
Наряду с распадом АТФ и выделением энергии в клетке постоянно происходит синтез АТФ и накопление энергии (обратные реакции).
Количество митохондрий в клетке зависит от потребности последней в энергии. Так, в клетках кожи человека находится в среднем 5–6 митохондрий, в клетках мышц – до 1000, в клетках печени – до 2500!
8. Комплекс Гольджи. Итальянский ученый Гольджи обнаружил и описал структуру клетки, напоминающую стопки мембран, цистерны, пузырьки и трубочки. Расположена эта система чаще всего возле ядра.
Функции комплекса Гольджи: в полостях комплекса накапливаются всевозможные продукты обмена клетки, которые по каким-либо причинам не вывелись наружу. В последствии эти продукты могут быть использованы клеткой для процессов жизнедеятельности. Из пузырьков и цистерночек комплекса Гольджи в растительных клетках образуются вакуоли, заполненные клеточным соком.
9. Лизосомы – мелкие органоиды. Представляют собой пузырьки, окруженные мембраной. Внутри лизосомы заполнены пищеварительными ферментами (обнаружено 12 ферментов), которые расщепляют и переваривают крупные макромолекулы (белки, полисахариды, нуклеиновые кислоты).
Функции лизосом: растворение и переваривание макромолекул. Лизосомы участвуют в фагоцитозе. Понятно, что основная функция по перевариванию поступающих в клетку частиц принадлежит лизосомам.
10. Пластиды. Эти органоиды характерны только для растительных клеток. Форма напоминает двояковыпуклую линзу. Структура пластид напоминает таковую у митохондрий: двойной слой мембраны. Наружная – гладкая, внутренняя образует складки, называемые тилакоидами. На тилакоидах происходит основной жизненно важный для всех зеленых растений процесс – фотосинтез:

Пластиды бывают трех типов:
1) Хлоропласты – зеленые пластиды. Их цвет обусловлен наличием хлорофилла. Хлорофилл – основное вещество хлоропластов (имеет зеленый цвет). Только благодаря хлорофиллу возможен процесс фотосинтеза (см. раздел 4.2). Хлоропласты придают зеленый цвет растительным организмам.
2) Хромопласты – пластиды, имеющие различные окраски: от ярко-желтого до пурпурно-багряного. Наличие различных пигментов окрашивают плоды, цветки и осенние листья растений в соответствующие цвета. Этот факт особенно важен для привлечения насекомых к цветкам, как природный индикатор созревания плодов и др.
3) Лейкопласты – бесцветные пластиды, в которых происходит накопление запасных питательных веществ (например, крахмала).
Некоторые виды пластид могут переходить друг в друга: например, переход хлоропластов в хромопласты: созревание томатов, яблок, вишни, и т. д.; изменение окраски листьев в осенний период времени. Лейкопласты могут переходить в хлоропласты: позеленение картофеля на свету. Это доказывает общность происхождения пластид.
11. Клеточные включения. Вакуоли. Это непостоянные и необязательные составляющие клетки. Они могут появляться и исчезать в течение всей жизни клетки. К ним относятся капли жира, зерна крахмала и гликогена, кристаллы щавелево-кислого кальция и др. Жидкие продукты обмена называются клеточным соком и накапливаются они в вакуолях. В клеточном соке растворены сахара, минеральные соли, пигменты и т. д. Чем старше клетка, тем больше клеточного сока накапливает клетка. Молодые клетки практически не содержат вакуолей.
Помимо перечисленного некоторые специализированные клетки обладают специальными органоидами. К ним относятся:
– реснички и жгутики, представляющие собой выросты мембраны клетки, осуществляющие движения клетки. Они имеются у одноклеточных организмов и многоклеточных (кишечный эпителий, сперматозоиды, эпителий дыхательных путей);
– миофибриллы – тонкие нити мышечных клеток, участвующие в сокращении мышц;
– нейрофибриллы – органоиды, характерные для нервных клеток и участвующие в проведении нервных импульсов. Кроме того, в состав клеток входят центриоли – две (иногда более) цилиндрические структуры диаметром около 0,1 мкм и длиной 0,3 мкм. Место расположения центриолей в период между делениями клетки считается серединой клеточного центра. При делении клетки центриоли расходятся в противоположные стороны – к полюсам, определяя ориентацию веретена деления (рис. 16).
Следует иметь в виду, что, хотя животные и растительные клетки имеют много общего, но между ними существуют и серьёзные различия (табл. 3.1).
Более общая классификация клеток представлена на рис. 3.16.
Одно из основных отличий бактерий от архей, состоит в химическом составе мембраны. Бактерии отделены от внешней среды двойным слоем липидов (жиров и жироподобных веществ). Мембраны архей состоят из терпеновых спиртов.
Источник
Типы питания прокариот
Прокариоты характеризуются многообразием типов питания в отличие от растений (автотрофы) и животных (ге- теротрофы). Для характеристики типов питания используют три критерия: источник углерода, источник энергии, донор электронов (водорода). По источнику углерода делят на ав- тотрофы и гетеротрофы, по источнику энергии – на фото- трофы (используют солнечный свет) и хемотрофы (получа- ют энергию за счет окислительно-восстановительных реак- ций), по донору – электроны (водорода) на литотрофы (неорганические соединения Н2, NH3, H2S, Fe2 + , CO и др.) и органотрофы (используют в качестве донора электрона ор- ганические соединения). Таким образом, выделяют 4 основных типа питания прокариот: фотолитоавтотрофы, фотоорганоавтотрофы, хемолитоавтотрофы, хемоорганоге- теротрофы.
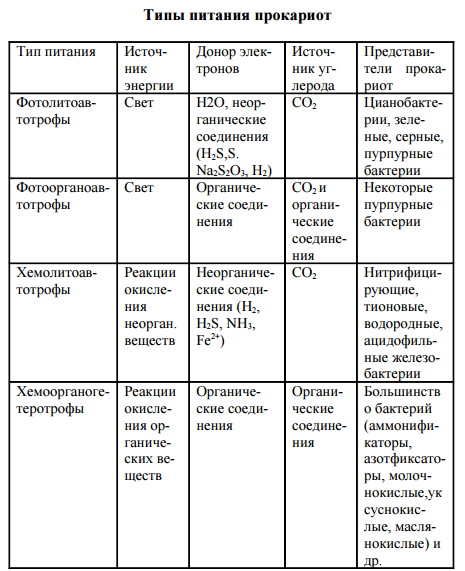
Фотолитоавтотрофы. Бактериальный фотосинтез (фоторедукция). В 1931 г. К. ван Ниль впервые доказал способность бактерий к фотосинтезу. В основе бактериального фотосинтеза лежит превращение световой энергии, поглощаемой фотосинтетическим пигментом, в биохимическую энергию макроэргических связей (АТФ) и далее использо- вание этой энергии для восстановления углекислого газа в процессе биосинтеза. У этих организмов есть пигмент бак- териохлорофилл. В клетках всех фотосинтезирующих бактерий содержатся фотосинтетические пигменты. К ним относятся особые хлорофиллы, получившие название бактериохлорофиллов а, в, с, d, и каротиноиды. По строению бактерио- хлорофиллы близки к хлорофиллу а растений. Так, бакте- риохлорофилл а отличается от хлорофилла а растений тем, что в первом его пирольном кольце в положении 2 стоит ацетильная группа СН3-СО- вместо обычной винильной группы СН2=СН-, а второе пирольное кольцо восстановле- но и содержит на два атома водорода больше. Содержание в клетке фотосинтезирующих прокариот четырех видоизменений бактериохлорофилла обусловли- вает более широкую полосу спектра поглощения световой энергии по сравнению с хлорофиллом растений. Предел спектра поглощения хлорофилла растений лежит в области 700-780 нм, в то время как фотосинтезирующие бактерии используют световую энергию длинноволновой части спектра до 1100 нм. Разница в спектрах поглощения хлоро- филлов растений и бактериохлорофиллов фотосинтезиру- ющих бактерий расширяет экологические ниши последних и позволяет фотосинтезирующим бактериям развиваться в водоемах под слоем водорослей. Помимо бактериохлорофиллов, в клетках фотосинтезирующих бактерий открыты более 20 дополнительных каро- тиноидных пигментов. Особенно разнообразны каротинои- ды в клетках пурпурных бактерий. Каротиноиды фотосин- тезирующих бактерий поглощают световую энергию ко- ротковолновой видимой области спектра с длиной волны 400-550 нм и передают эту энергию на бактериохлоро- филл. В клетке прокариот фотосинтетические пигменты нахо- дятся на инвагинациях ЦПМ – хроматофорах, визикулах (пузырьках), трубочках, тиллакоидах, ламеллах. Ван Ниль назвал этот процесс бактериальным фотосин- тезом, или фоторедукцией.
Для восстановления одной молекулы углекислого газа требуется 1 молекула АТФ (у растений 4 АТФ). В том и другом случае идет восстановление углекислого газа до углеводов. В Определителе Берджи (1974) фотосинтезирующие бактерии представлены тремя семействами (см. выше).
Фотоорганоавтотрофы Фотоорганоавтотрофы представлены немногочисленным семейством Rhodospirillaceae, включающим три рода: Rhodospirillum, Rhodopseudomonas и Rhodomicrobium. Бактерии-фотоорганоавтотрофы способны перестраивать свой обмен и одинаково успешно развиваться как на свету, так и в темноте, переходя соответственно от ана- эробного образа жизни к аэробному. На свету они ведут себя как фотоорганоавтотрофы: усваивая углекислый газ, они восстанавливают его в процессе фотосинтеза до углевода. В качестве доноров электронов несерные пурпурные бактерии используют различные органические вещества — сахара, спирты, органические кислоты, аминокислоты. Характер использования органических веществ различными фотосинтезирующими бактериями существенно раз- личается. Чаще всего органическое вещество выполняет единственную функцию донора электронов при фотоасси- миляции углекислого газа. Однако в некоторых случаях органическое вещество используется фотосинтезирующими бактериями не только в качестве донора электронов, но одновременно и как источник углерода. Попадая в темноту, пурпурные бактерии переходят к хемоорганогетеротрофному типу питания. При этом энергию для процессов жизнедеятельности они получают за счет реакций окисления органического субстрата по циклу Кребса. Непосредственным источником углерода и донором электронов для них являются органические соедине- ния субстрата. В клетках бактерий этой группы имеется универсальный набор дыхательных ферментов (НАД- и ФАД-дегидрогеназы и цитохромы), обеспечивающих им возможность перехода от анаэробного образа жизни на свету к аэробному в темноте и, соответственно, от авто трофного типа питания к гетеротрофному. Таким образом, среди разных групп прокариот нет, да и не может быть, резкой границы между различными типами питания.
ХемолитоавтотрофыХемолитоавтотрофы представлены микроорганизмами, способными в качестве основного источника углерода усваивать углекислый газ и синтезировать в клетке органические соединения, используя энергию реакций окисления неорганического субстрата. Для хемолитоавтотрофов неорганические вещества субстрата выступают донорами электронов в реакциях энергетического метаболизма и в процессе хемоассимиляции углекислого газа. Заслуга открытия процесса хемосинтеза принадлежит С. Н. Виноградскому. Он определил химизм как жизнь без органики. Им впервые была показана возможность образования органических веществ из неорганических в клетках бактерий, помимо процесса фотосинтеза. К хемолитоавтотрофам относится большинство видов нитрифицирующих, тионовых бактерий, некоторые виды из группы одноклеточных железобактерий и водородные бактерии. Хемолитоавтотрофные бактерии характеризуются специфичностью в отношении использования окисляемого субстрата. Нитрифицирующие бактерии для процесса хемоассимиляции углекислого газа получают энергию от окисления аммиака и нитритов. Процесс нитрификации проходит в две фазы. Первая фаза заключается в аэробном окислении аммиака до нитритов нитрозными бактериями родов Nitrosomonas, Nitrosococcus, Nitrosolobus, Nitrosospira: NН3 + 11 /202 = HN02 + Н2О + 274,7 кДж. Вторая фаза нитрификации предусматривает окисление нитритов в нитраты нитратными бактериями родов Nitrobacter, Nitrospina, Nitrococcus: HN02 + 11 /202 =НNО3 + 87,5 кДж. При процессах окисления неорганического субстрата выделяется сравнительно небольшое количество энергии, и усваивается она клеткой с низким КПД, всего 5-10%. Поэтому для получения энергии на процессы жизнедеятельности нитрифицирующим бактериям приходится перерабатывать огромное количество субстрата. К хемолитоав- тотрофам относится большинство видов тионовых бактерий: Thiobacillus denitrificans, Т. thiooxidans, Т. acidophilus, Thiomicrospira pelophila и др. Они ведут процесс хемоасси- миляции углекислого газа, получая энергию за счет окисления восстановленных или частично восстановленных соединений серы, сероводорода, элементарной серы, тиосульфата и сульфита. Конечным продуктом окисления обычно является сульфат.
Некоторые виды тионовых бактерий (Thiobacillus ferrooxidans) способны получать энергию за счет окисления не только соединений серы, но и закисного железа (Fe2+) (разрушение труб). Хемолитоавтотрофный тип питания характерен для некоторых одноклеточных ацидофильных железобактерий – Leptospirillum ferrooxidans, Thiobacillus ferrooxidans и представителей рода Sulfolobus. Оптимальный рост ацидофильных железобактерий наблюдается при рН ниже 4,5 (2-3). Они используют энергию окисления Fe2+ до Fе3+ для ассимиляции С02, который служит основным или единственным источником углерода. Реакции окисления железа сопровождаются незначительным выделением энергии, поэтому железобактерии перерабатывают большие количества субстрата: 2Fe2+ + 1/202 + 2Н+ = 2FеЗ+ + Н2О + 33 кДж. Хемолитоавтотрофные бактерии в природе являются геологическими агентами. Они принимают участие в процессах образования полезных ископаемых и осуществляют важнейшие звенья круговорота азота, серы, железа. К факультативным хемолитоавтотрофам относятся водородные бактерии, являющиеся представителями 20 различных родов – Рsеudоmоnаs, Аlcаligеnеs, Nocardia и др. Они способны осуществлять ферментативное окисление водорода кислородом воздуха с образованием воды. Водородные бактерии ферментом дегидрогеназой активируют молекулярный водород и далее используют его для получения энергии и в качестве донора электронов для восстановления углекислого газа до углевода: 6Н2 + 202 + С02 = (СН2О) + 5Н2О. Являясь факультативными хемолитоавтотрофами, водородные бактерии в качестве источника энергии и углеро- да способны использовать и различные органические со- единения (сахара, органические кислоты, спирты). Таким образом, водородные бактерии могут служить еще одним 87 примером возможного перехода микробной клетки от одного типа питания к другому – от хемолитоавтотрофного к хемоорганогетеротрофному. Бактерии, переходящие от одного типа питания к другому, получили название миксо- трофов.
Хемоорганогетеротрофы. К хемоорганогетеротрофам относится большинство прокариот. Источником углерода для них являются самые разнообразные органические соединения. Энергию для жизнедеятельности они получают за счет окислительно- восстановительных реакций органического субстрата, и донором электронов в реакциях метаболизма также выступают различные органические вещества. Хемоорганогетеротрофы наиболее широко распространены в природе. Им принадлежит роль санитаров нашей планеты, так как они ведут процессы минерализации самых разнообразных, подчас сложных органических веществ. Помимо органических соединений как источника углерода, хемоорганогетеротрофы нуждаются в углекислом газе для реакций карбоксилирования промежуточного обмена. Хемоорганогетеротрофные микроорганизмы подразделяют на сапрофитов и паразитов. Сапрофиты потребляют органические вещества опада. Паразиты живут за счет органических веществ живой клетки. Выделяют факультативных и облигатных паразитов. Факультативные паразиты развиваются на обычных органических средах, но, попадая в клетку-хозяина, переходят к паразитическому образу жизни. К ним относится большинство патогенных бактерий, вызывающих заболевания человека, – возбудители пневмонии, менингита, гонореи, дизентерии, брюшного тифа, сибирской язвы, коклюша, туберкулеза и др. Облигатные (строгие) паразиты развиваются исключительно за 88 счет органических веществ клетки-хозяина. Типичным примером облигатных паразитов являются риккетсии и вирусы. В основе всех вышерассмотренных типов питания прокариот лежат различные сочетания их энергетического и конструктивного метаболизма. Всем типам питания соот- ветствуют определенные группы прокариотных организ- мов. Основной массе бактерий присущ один вполне определенный (облигатный) тип питания. Однако среди многообразного мира прокариот встречается много видов бактерий, способных переходить от одного типа питания к другому. Так, среди цианобактерий, зеленых и пурпурных бактерий имеются виды, переходящие от фотолитоавто- трофного типа питания к фотоорганогетеротрофному. Некоторые виды обширного рода Thiobacillus способны переходить от хемолитоавтотрофного типа питания к хемоорга- ногетеротрофному. Эта особенность питания прокариот обусловливает широкие возможности существования микроорганизмов в различных условиях среды и значительно расширяет их экологические ниши.
Источник



