
Механизмы передачи гормональных сигналов в клетки
По механизму действия гормоны можно разделить на 2 группы. К первой группе относят гормоны, взаимодействующие с мембранными рецепторами (пептидные гормоны, адреналин, а также гормоны местного действия — цитокины, эйкозаноиды). Вторая группа включает гормоны, взаимодействующие с внутриклеточными рецепторами.
Связывание гормона (первичного посредника) с рецептором приводит к изменению конформации рецептора. Это изменение улавливается другими макромолекулами, т.е. связывание гормона с рецептором приводит к сопряжению одних молекул с другими (трансдукция сигнала). Таким образом, генерируется сигнал, который регулирует клеточный ответ путём изменения активности или количества ферментов и других белков. В зависимости от способа передачи гормонального сигнала в клетках меняется скорость реакций метаболизма путем воздействия на активность ферментов либо на количество ф 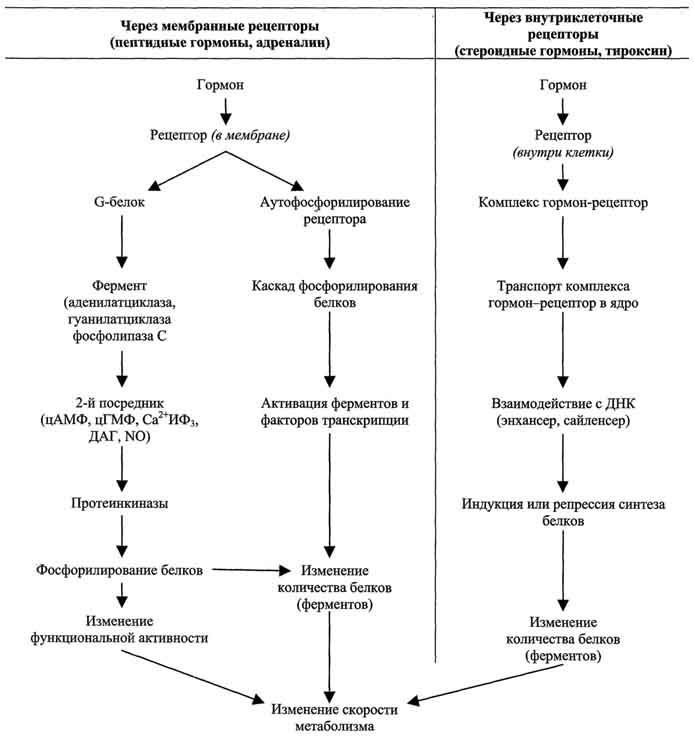 ерментов.
ерментов.
1. Передача гормональных сигналов через мембранные рецепторы
Гормоны (первичные посредники), связываясь с рецепторами на поверхности клеточной мембраны, образуют комплекс гормон-рецептор, который трансформирует сигнал первичного посредника в изменение концентрации особых молекул внутри клетки — вторичных посредников. Вторичными посредниками могут быть следующие молекулы: цАМФ, цГМФ, ИФ3, ДАТ, Са 2+ , NO.
Гормоны, взаимодействие которых с рецептором клетки-мишени приводит к образованию цАМФ, действуют через трёхкомпонентную систему, которая включает белок-рецептор, G-белок и фермент аденилатциклазу. Образующийся под действием аденилатциклазы цАМФ активирует протеинкиназу А, фосфорилируюшую ферменты и другие белки. Известно более 200 различных G-белков, в структуре которых обнаружены 3 субъединицы α, β и γ. В отсутствие гормона α-субъединица G-белка связана с ГДФ. Образование комплекса гормонрецептора приводит к конформационным изменениям α-субъединицы, замене ГДФ на ГТФ и отщеплению димера βγ от α-ГТФ. В случае рецепторов, сопряжённых с Gs-белком, субъединица αs-ГТФ активирует аденилатциклазу.
( 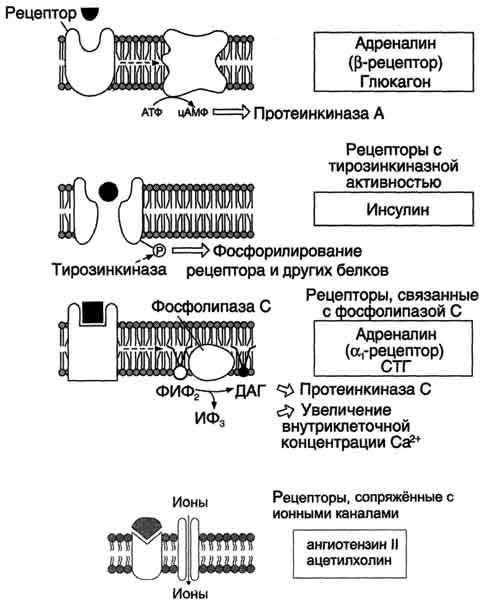 ИФ3 — инозитол-3-фосфат; ДАГ — диацилглицерол; ФИФ2 — фосфоинозитолбисфосфат; СТГ — соматотропный гормон. инозитоп-3-фосфат; ДАГ — диаципглицерол; ФИФ2 — фосфоинозитолбифосфат; СТГ — соматотропный гормон.)
ИФ3 — инозитол-3-фосфат; ДАГ — диацилглицерол; ФИФ2 — фосфоинозитолбисфосфат; СТГ — соматотропный гормон. инозитоп-3-фосфат; ДАГ — диаципглицерол; ФИФ2 — фосфоинозитолбифосфат; СТГ — соматотропный гормон.)
В случае рецепторов, сопряжённых с Giбелком, субъединица αi-ГТФ ингибирует аденилатциклазу. В таблице 11-4 приведены примеры гормонов, взаимодействие которых с соответствующим рецептором активирует или ингибирует аденилатциклазу.
Другая система, генерирующая цГМФ как вторичный посредник, сопряжена с гуанилатциклазой. Цитоплазматический домен такого типа рецепторов обладает активностью гуанилатциклазы, которая катализирует реакцию образования цГМФ из ГТФ> (подобно аденилатциклазе). Молекулы цГМФ могут активировать ионные каналы либо активировать цГМФ-за-висимую протеинкиназу G, участвующую в фосфорилировании других белков в клетке. Например, фосфодиэстерааа, которая гидролизует цАМФ до АМФ, активируется в результате фосфорилирования цГМФ-зависимой протеинкиназой.
Некоторые гормоны (например, вазопрессин или адреналин), образуя комплекс с соответствующими рецепторами (рецептор V1 для вазопрессина и αi-рецептор для адреналина), через активацию соответствующих G-белков активируют фосфолипазу С, в результате чего в клетке появляются вторичные посредники ИФ3, ДАГ. Молекула ИФ3 стимулирует высвобождение Са 2+ из ЭР. Кальций связывается с белком кальмодулином. Этот комплекс активирует Са 2+ -кальмодулинзависимую протеинкиназу. Ионы кальция и ДАГ участвуют в активации протеинкиназы С.
Источник
43. Механизмы передачи сигнала в клетку.
Передача сигнала в клетке (клеточная сигнализация, англ. cell signaling) — это часть сложной системы коммуникации, которая управляет основными клеточными процессами и координирует действия клетки. Возможность клеток корректно отвечать на изменения окружающей их среды (microenvironment) является основой развития, репарации тканей, иммунитета и системы поддержания гомеостаза в целом. Ошибки в системах обработки клеточной информации могут привести к раку, аутоиммунным заболеваниям и диабету. Понимание механизмов передачи сигнала внутри клетки может привести к разработке методов лечения заболеваний и даже созданию искусственных тканей.
Сигналы, передающиеся через сигнальные молекулы, являются первичными по отношению к каскадам биохимических реакций, запускающимся в клетках в ответ на их воздействие. Передача сигнала это последовательность реакций, включающих взаимодействие внеклеточных лигандов (сигналы клеточные) с рецепторами на поверхности клетки с последующей активацией рецептора, заключающейся в изменении состояния его внутриклеточного домена. Активация рецептора вызывает каскад событий в клетке, в результате которых клетка адекватно реагирует на внешний сигнал.
Первичные сигналы распознаются клетками благодаря наличию у них специальных молекул-рецепторов белковой природы, взаимодействующих с первичными сигнальными молекулами или с физическими факторами. Первичный сигнал, как правило, не действует прямо на те метаболические процессы в клетке, для регуляции которых он предназначен. Воспринимающий его рецептор инициирует образование в клетке промежуточных химических соединений, запускающих внутриклеточные процессы, воздействие на которые было целью первичного внеклеточного сигнала. Такие промежуточные соединения несут в себе информацию о первичном регуляторном сигнале и являются вторичными его переносчиками, поэтому они получили название вторичных мессенджеров . Ими могут быть различные ионы, циклические нуклеотиды , продукты деградации липидов и целый ряд других химических соединений биогенного происхождения.
Вторичные мессенджеры позволяют усиливать первичный регуляторный сигнал от внеклеточных регуляторных молекул. Группы клеток и тканей приобретают способность к однотипной и одновременной реакции на первичный регуляторный сигнал, например, на действие гормона эндокринной системы. Это обеспечивает возможность быстрой адаптации многоклеточного организма к изменяющимся условиям внутренней и окружающей среды.
Изучение механизмов передачи и усиления сигналов является одной из основных задач биологии клетки. Их знание необходимо для понимания механизмов формирования функционального ответа клеток в норме, его регуляции и коррекции при патологических состояниях. В настоящее время известно около 50 белков-лигандов и 14 семейств рецепторов.
Существует несколько более или менее стандартных способов передачи сигнала от клеточной поверхности внутрь клетки, хотя эта проблема еще далека от окнчательного понимания и постоянно появляются новые варианты сигнализации. Например классический обобщенный путь передачи сигнала заключается в цепоче взаимодействий -сигнальная молекула — рецептор на поверхности клетки-внутриклеточный усилительный механизм -включение определенных специфичных для данного сигнала генов. дает упрощенную схему двух возможных путей многостадийного процесса предачи сигнала, которая начинается со взаимодействия некоторого внешнего фактора с рецептором на поверхности клетки. Таким внешним фактором может быть какой либо гормон или какой-нибудь фактор роста, в частности, цитокин.
Источник
Тема 7. Молекулярные механизмы передачи сигнала в клетку. Основные механизмы внутриклеточной сигнализации.
Цель: Изучение молекулярных механизмов межклеточной сигнализации, основных этапов передачи сигналов в клетку, способов доставки сигнальных молекул к клеткам.
Изучить способы передачи в клетку внешних сигналов и механизмов клеточного ответа.
Изучить роль вторичных мессенджеров в системе жизнеобеспечения клеток.
Основные вопросы темы:
1. Роль сигнализации для жизнедеятельности клеток
2. Общая характеристика сигнальных молекул.
3.Общая характеристика семейства мембраносвязанных и внутриклеточных рецепторов.
4. Этапы передачи сигналов.
5. Характеристика вторичных мессенджеров.
Для нормальной жизнедеятельности любого организма составляющие его клетки, подобно людям в социуме, должны чутко реагировать на меняющуюся ситуацию, регулируя свою работу в зависимости от окружающих условий.
Клетка изменяет активности своих ферментов в ответ на получаемые ею из внешней среды специфические сигналы. Эти сигналы представляют собой определенные низкомолекулярные вещества (лиганды), связывающиеся со специальными участками клеточной поверхности – рецепторами. В организме человека лигандами являются, например, нейротрансмиттеры, которые выделяются в синаптических щелях нервными клетками в ответ на нервный импульс, а также вещества, секретируемые другими клетками в окружающую их среду.
Рисунок 1 Основные механизмы трансмембранной передачи сигнальной информации:

I — прохождение растворимой в жирах сигнальной молекулы через клеточную мембрану;
II — связывание сигнальной молекулы с рецептором и активация его внутриклеточного фрагмента; III — регулирование активности ионного канала; IV — передача сигнальной информации с помощью вторичных передатчиков.
1 — лекарство; 2 — внутриклеточный рецептор; 3 — клеточный (трансмембранный) рецептор; 4 — внутриклеточное превращение (биохимическая реакция); 5 — ионный канал; 6 — поток ионов; 7 — вторичный посредник; 8 — фермент или ионный канал; 9 — вторичный посредник.
Первый механизм (участок I на рисунке 1) – растворимая в липидах сигнальная молекула проходит через клеточную мембрану и активирует внутриклеточный рецептор (например, фермент). Так действует оксид азота, ряд жирорастворимых гормонов (глюкокортикоиды, минералокортикоиды, половые гормоны и тиреоидные гормоны) и витамин D. Они стимулируют транскрипцию генов в ядре клетки и, таким образом, синтез новых белков. Механизм действия гормонов заключается в стимуляции синтеза новых белков в ядре клетки.
Второй механизм передачи сигнала через клеточную мембрану (участок II на рисунке 1) – это связывание с клеточными рецепторами, имеющими внеклеточный и внутриклеточный фрагменты (то есть трансмембранными рецепторами). Внеклеточная и внутриклеточная части подобных рецепторов связаны полипептидным мостиком, проходящим через клеточную мембрану. Внутриклеточный фрагмент обладает ферментативной активностью, которая повышается при связывании сигнальной молекулы с рецептором. Соответственно возрастает скорость внутриклеточных реакций, в которых участвует этот фрагмент.
Третий механизм передачи информации – действие на рецепторы, регулирующие открытие или закрытие ионных каналов (участок III на рисунке 1). К естественным сигнальным молекулам, взаимодействующим с такими рецепторами, относятся, в частности, ацетилхолин, гамма-аминомасляная кислота (ГАМК), глицин, аспартат, глутамат и другие. При их взаимодействии с рецептором происходит увеличение трансмембранной проводимости для отдельных ионов, что вызывает изменение электрического потенциала клеточной мембраны. Например, ацетилхолин, взаимодействуя с н-холинорецепторами, увеличивает вход в клетку ионов натрия и вызывает деполяризацию и мышечное сокращение. Взаимодействие гамма-аминомасляной кислоты со своим рецептором приводит к повышению поступления ионов хлора в клетки, усилению поляризации и развитию торможения (угнетения) центральной нервной системы. Этот механизм передачи сигналов отличает быстрота развития эффекта (миллисекунды).
Четвертый механизм передачи химического сигнала реализуется через рецепторы, активизирующие внутриклеточный вторичный передатчик (участок IV на рисунке 1). При взаимодействии с такими рецепторами процесс протекает в четыре этапа. Сигнальная молекула распознается рецептором на поверхности клеточной мембраны, в результате их взаимодействия рецептор активизирует G-белок на внутренней поверхности мембраны. Активизированный G-белок изменяет активность либо фермента, либо ионного канала. Это приводит к изменению внутриклеточной концентрации вторичного посредника, через который уже непосредственно реализуются эффекты (изменяются процессы обмена веществ и энергии). Такой механизм передачи сигнальной информации позволяет усилить передаваемый сигнал. Так если взаимодействие сигнальной молекулы (например, норадреналина) с рецептором длится несколько миллисекунд, то активность вторичного передатчика, которому рецептор передает по эстафете сигнал, сохраняется в течение десятков секунд.
Вторичные посредники – это вещества, которые образуются внутри клетки и являются важными компонентами многочисленных внутриклеточных биохимических реакций. От их концентрации во многом зависит интенсивность и результаты жизнедеятельности клетки и функционирование всей ткани. Каким же образом первичные мессенджеры оказывают влияние на активность белков и, тем самым, на активность клетки?
Для этого внешний по отношению к клетке сигнал должен превратиться во внутренний. Ключевую роль в таком процессе играют образующиеся внутри клетки вторичные мессенджеры, которых на удивление мало. Главенствующую роль среди них играет циклический аденозинмонофосфат (цАМФ), открытый в 1958 г. Э.Сазерлендом и Т.Роллом.
Это соединение образуется из знаменитой АТФ – аденозинтрифосфорной кислоты, которую часто называют «энергетической разменной монетой» клетки. Как известно, АТФ состоит из азотистого основания аденина, пятиуглеродного циклического сахара и трех остатков фосфорной кислоты. Химические связи между фосфорными остатками богаты энергией. Практически все внутриклеточные процессы, начиная от синтеза белков и кончая мышечным сокращением, получают энергию за счет отщепления одной или двух фосфатных групп АТФ.
Циклический аденозинмонофосфат образуется с помощью фермента аденилатциклазы, который отщепляет от АТФ два остатка фосфорной кислоты, а последний, третий, остаток замыкает через два атома кислорода на сахар рибозу, входящий в состав аденозина. Это удивительное вещество играет роль универсального вторичного мессенджера (посредника).
Наиболее известными вторичными посредниками кроме цАМФ являются циклический гуанозинмонофосфат (цГМФ), ионы кальция, калия, диацилглицерол и инозитолтрифосфат.
Какие воздействия могут реализовываться с участием вторичных посредников?
цАМФ участвует в мобилизации энергетических запасов (распад углеводов в печени или триглицеридов в жировых клетках), в задержке воды почками, в нормализации кальциевого обмена, в увеличении силы и частоты сердечных сокращений, в образовании стероидных гормонов, в расслаблении гладких мышц и так далее.
Диацилглицерол, инозитолтрифосфат и ионы кальция участвуют в реакциях, которые возникают в клетках при возбуждении некоторых типов адрено- и холинорецепторов.
цГМФ участвует в расслаблении гладких мышц сосудов, стимулируя образование оксида азота в эндотелии сосудов под влиянием ацетилхолина и гистамина.
Таким образом, как транспортные системы, так и механизмы передачи сигнала участвуют в реализации и выполнении двух основных функций (задач) клетки: поддержание стабильности “системы жизнеобеспечения” и выполнение специальных функций.
Рис.2. Главные механизмы образования внутриклеточных мессенджеров
Самостоятельная работа студентов :
Раб.1. В схематической форме составить четыре этапа Четвертого механизма передачи химического сигнала в клетку.
Раб.2. Рассмотреть и подготовить схему «Главные механизмы образования внутриклеточных мессенджеров» к итоговому контролю занятия.
1. Контроль исходного уровня знаний
2. Разбор результатов самостоятельной работы студентов
3. Контроль итогового уровня знаний
Методы обучения и преподавания: проведение практических занятий, работа с компьютерными моделями и микропрепаратами.
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Источник



