
- Общая характеристика способов обмена генетической информацией у бактерий
- Механизмы генетической рекомбинации бактериальной ДНК: трансформация, трансдукция, конъюгация
- Рекомбинация у бактерий: трансформация, трансдукция, конъюгация
- Механизмы передачи генетического материала у бактерий: конъюгация, трансдукция, трансформация.
Общая характеристика способов обмена генетической информацией у бактерий
У бактерий, как и у высших организмов, существуют механизмы, которые обеспечивают перекомбинацию генетического материала между родственными, но генетически неидентичными клетками. В результате такого обмена возникает потомство с новой комбинацией наследственных признаков – рекомбинанты. Однако способы обмена генетической информацией и процессы формирования рекомбинантов у бактерий отличаются от существующих у эукариот, что вносит своеобразие в рекомбинационный анализ генетической организации бактериальной хромосомы.
Известны три механизма передачи генетического материала у бактерий: конъюгация, трансформация и трансдукция.
Конъюгация – это направленный перенос генетического материала из клетки-донора в клетку-реципиент при непосредственном физическом контакте между ними.
Трансформация – способ передачи генетической информации от клеток-доноров к клеткам-реципиентам с помощью химически чистой ДНК.
Трансдукция – это перенос генетического материала из клеток-доноров в клетки-реципиенты с помощью бактериофагов.
Основная отличительная черта всех трех механизмов – это однонаправленный перенос от одной клетки (донора) к другой (реципиенту) части генетического материала (рис.5). Вследствие такого обмена возникает не полноценная диплоидная зигота, а частично диплоидная (меродиплоидная) зигота, или, так называемая, мерозигота. В мерозиготе генетический материал реципиентной клетки представлен целой хромосомой, а материал донорной клетки только фрагментом хромосомы или частью генома.
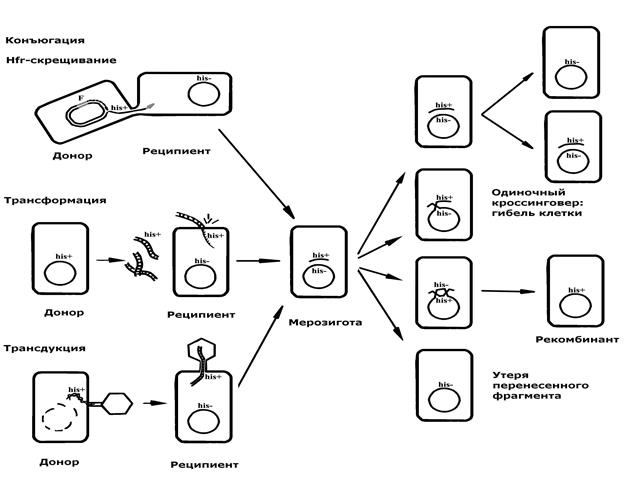 |
Рис. 5. Механизмы передачи генетического материала у бактерий и судьба перенесенного фрагмента
Судьба перенесенного фрагмента донорной ДНК в мерозиготе неоднозначна и зависит от многих факторов. Обычно он не способен к самостоятельной редупликации в клетке реципиента, так как не является репликоном, но некоторое время может в ней сохраняться и даже функционировать (транскрибироваться). В этом случае фрагмент ДНК донора при каждом делении мерозиготы будет передаваться лишь одной из дочерних клеток (однолинейная передача). Однако такое состояние не стабильно и может привести либо к утере фрагмента, либо к включению его в хромосому реципиентной клетки за счет рекомбинации (кроссинговера или конверсии) с ее гомологичной последовательностью.
Важно отметить ту особенность рекомбинации у бактерий, что только четное число обменов между хромосомой реципиента и переданным фрагментом донора дает полноценную хромосому. Единичный или нечетный кроссинговер приводит к размыканию хромосомы и дупликации концевых фрагментов (рис.5). Следовательно, элементарным регистрируемым актом рекомбинации в мерозиготе является двойной кроссинговер. Результатом двойного (или четного) обмена является реципрокная интеграция донорного фрагмента в реципиентную хромосому и участка реципиентной хромосомы в донорный фрагмент. Так как донорный фрагмент обычно не реплицируется и в дальнейшем теряется, регистрируется только интеграция в хромосому. Включенный фрагмент (или его участок) будет реплицироваться совместно с хромосомой реципиента и передаваться потомству. В результате в популяции реципиентных клеток появятся стабильные рекомбинанты, получившие некоторые признаки донора. Отмеченная особенность рекомбинации приводит к нереципрокности и уменьшенному вдвое выходу рекомбинантов.
Следует также напомнить, что бактериальные клетки в различных условиях могут содержать несколько копий геномных молекул ДНК. Поэтому рекомбинанты появляются не сразу, а после 3-4 последовательных делений, когда клетки становятся гаплоидными (несут одинаковые копии).
Размер перенесенного фрагмента хромосомы донора зависит от способа передачи генетического материала. Например, при трансдукции он ограничен объемом головки бактериофага, в которую может быть ошибочно упакована ДНК клетки-хозяина. При трансформации длина переносимой молекулы очень мала и зависит от способа приготовления препарата ДНК и специфики конкретной системы. Только при конъюгации размер переносимого генетического материала может значительно колебаться и в редких случаях достигать размера полного генома.
Дата добавления: 2015-02-07 ; просмотров: 2398 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник
Механизмы генетической рекомбинации бактериальной ДНК: трансформация, трансдукция, конъюгация




![]()
![]()
Длительное время считалось, что бактерии представляют собой изолированные генетические системы и каждая особь имеет одного (и только одного) родителя, т. е. их изменчивость обусловленалишь мутациями; никто не мог себе представить, что подобно гаметам высших организмов бактерии способны обмениваться генетическим материалом и, по аналогии с половым размножением, давать начало потомству с новыми свойствами.
Прокариотам не свойственно половое размножение. Рекомбинация у них происходит в результате внутригеномных перестроек, заключающихся в изменении локализации генов в пределах хромосомы, или при проникновении в клетку реципиента части ДНК донора. Последнее приводит к формированию неполной зиготы — мерозиготы. В результате рекомбинаций в мерозиготе образуется только один рекомбинант, генотип которого представлен в основном генотипом реципиента с включенным в него фрагментом ДНК донора.
Генетические рекомбинации происходят при участии ряда ферментов в пределах отдельных генов или групп сцепленных генов. Существуют специальные rec-гены, детерминирующие рекомбинационную способность бактерий. Передача генетического материала (хромосомных генов) от одних бактерий к другим происходит путем трансформации, трансдукции и конъюгации.
Передача плазмидных генов — путем трансдукции и конъюгации.
Трансформация (изменение одного типа клеток при действии активного начала из другого типа клеток). Феномен открыл Гриффит у Streptococcus pneumoniae (1928); позднее Эвери, Маклеод и Мак Карти (1944) выделили трансформирующее начало пневмококков в форме молекулы ДНК. Это и явилось первым прямым доказательством того, что носителем генетической информации является ДНК. Погибшие бактерии постоянно высвобождают ДНК, которая может быть воспринята другими бактериями. Как правило, любая чужеродная ДНК, попадающая в бактериальную клетку, расщепляется рестрикционными эндонуклеазами; но при некоторых условиях такая ДНК может быть интегрирована в геном бактерий. По происхождению ДНК может быть плазмидной либо хромосомной и нести гены «трансформирующие» реципиента. Подобным путем процессы трансформации могут распространять гены, кодирующие факторы вирулентности, среди бактериальных популяций; однако в обмене генетической информацией трансформация играет незначительную роль.
Трансдукция. Явление открыл Ледерберг и Циндер (1952); в ходе трансдукции ДНК (но не нуклеопротеид) переносится от одной клетки (донор) к другой (реципиент) с помощью бактериофагов; обычно при этом фаг переносит лишь небольшой фрагмент ДНК хозяина. Выделяют 3 типа трансдукции: неспецифическую (общую), специфическую и абортивную. Неспецифическая трансдукция — в клетке, инфицированной бактериофагом, в ходе сборки дочерней популяции в головки некоторых фагов вместе с вирусной ДНК может проникнуть фрагмент бактериальной ДНК или плазмиды. При этом фаговая частица может утратить часть своего генома и стать дефектной. При такой форме трансдукции в клетки-реципиенты могут быть внесены практически любые гены. Специфическая трансдукция — характеризуется способностью фага переносить определенные гены от бактерии-донора к бактерии-реципиенту. Это связано с тем, что образование трансдуцирующего бактериофага происходит путем выщепления профага из бактериальной хромосомы вместе с генами, расположенными на хромосоме в клетке-донора рядом с профагом. При взаимодействии трансдуцирующих фагов с клетками реципиентного штамма происходит включение гена бактерии-донора вместе с ДНК дефектного фага в хромосому бактерии-реципиента. Бактерии, лизогенированные дефектным фагом, невосприимчивы, как и все лизогенные клетки, к последующему заражению гомологичным вирулентным фагом. Абортивная трансдукция — принесенный фагом фрагмент ДНК бактерии-донора не включается в хромосому бактерии-реципиента, а располагается в ее цитоплазме и может в таком виде функционировать. Во время деления бактериальной клетки трансдуцированный фрагмент ДНК-донора может передаваться только одной из двух дочерних клеток, т. е. наследоваться однолинейно и постепенно утрачиваться.
Конъюгация — перенос генетического материала их клетки-донора в клетку-реципиента при их скрещивании. Процесс конъюгации у бактерий впервые обнаружен Д. Ледербергом и Э. Тейтумом в 1946 г. Позднее выяснилось, что донорами генетического материала являются клетки, несущие F-плазмиду (половой фактор). При скрещивании F + c F — клеткой половой фактор передается независимо от хромосомы донора, если плазмида находится в автономном состоянии. При этом почти все реципиентные клетки получают F плазмиду и становятся F + клетками. Первым этапом конъюгации является прикрепление клетки-донора к реципиентной клетке с помощью половых ворсинок (sex pili). Затем между обеими клетками образуется конъюгационный мостик, через который из клетки-донора в клетку-реципиент могут передаваться F-фактор и другие плазмиды, находящиеся в цитоплазме бактерии-донора в автономном состоянии.

Интеграция F-плазмиды в состав бактериальной хромосомы приводит к разрыву одной из нитей ДНК, что обеспечивает возможность переноса в реципиентную клетку. Такие штаммы бактерий-доноров получили название Hfr-штаммы (с англ. — высокая частота рекомбинаций). При скрещивании Hfr-штамма с F — бактериями F-фактор, как правило, не передается, поскольку он расположен в дистальной части хромосомы.
Источник
Рекомбинация у бактерий: трансформация, трансдукция, конъюгация
![]()
![]()
Рекомбинации (обмен генетическим материалом) у бактерий отличаются от рекомбинаций у эукариот:
• у бактерий имеется несколько механизмов рекомбинаций;
• при рекомбинациях у бактерий образуется не зигота, как у эукариот, а мерозигота (несет полностью генетическую информацию реципиента и часть генетической информации донора в виде дополнения);
• у бактериальной клетки-рекомбината изменяется не только качество, но и количество генетической информации.
Трансформация — это обмен генетической информацией у бактерий путем введения в бактериальную клетку-реципиент готового препарата ДНК (специально приготовленного или непосредственно выделенного из клетки-до нора). Чаще всего передача генетической информации происходит при культивировании реципиента на питательной среде, содержащей ДНК донора. Для восприятия донорской ДНК при трансформации клетка-реципиент должна находиться в определенном физиологическом состоянии (компетентности), которое достигается специальными методами обработки бактериальной популяции.
При трансформации передаются единичные (чаще 1) признаки. Трансформация является самым объективным свидетельством связи ДНК или ее фрагментов с тем или иным фенотипическим признаком, поскольку в реципиентную клетку вводится чистый препарат ДНК.
Трансдукция — обмен генетической информацией у бактерий путем передачи ее от донора к реципиенту с помощью умеренных (трансдуцирующих) бактериофагов.
Трансдуцирующие фаги могут переносить 1 или более генов (признаков).
• специфической — переносится всегда один и тот же ген;
• неспецифической — передаются разные гены.
Это связано с локализацией трансдуиируюших фагов в геноме донора:
• в случае специфической трансдукции они располагаются всегда в одном месте хромосомы;
• при неспецифической их локализация непостоянна.
Конъюгация — обмен генетической информацией у бактерий путем передачи ее от донора к реципиенту при их прямом контакте. После образования между донором и реципиентом конъюгационного мостика одна нить ДНК-донора поступает по нему в клетку-реципиент. Чем дольше контакт, тем большая часть донорской ДНК может быть передана реципиенту.
Основываясь на прерывании конъюгации через определенные промежутки времени, можно определить порядок расположения генов на хромосоме бактерий — построить хромосомные карты бактерий (произвести картирование бактерий).
Источник
Механизмы передачи генетического материала у бактерий: конъюгация, трансдукция, трансформация.
![]()
![]()
1. Трансформация – передача генетической информации через выделенную из клетки-донора ДНК. 1928г. Ф Гриффит (опыт с вирулентнотью пневмококков)
Трансформирующей активностью обладает только двунитчатая высокоспирализованная ДНК. В клетку-реципиент проникает только одна нить ДНК, другая – в клеточной мембране подвергается деградации с освобождением энергии, необходимой для проникновения в клетку второй нити ДНК.
Интеграция с хромосомой требует наличия гомологичных участков с трансформирующей ДНК.
Процесс трансформации зависит от компетентности клетки-реципиента и состояния трансформирующей донорской ДНК.
Компетентность клеток бактерий (способность воспринимать трансформирующую ДНК) зависит от присутствия в ЦПМ особых белков, обладающих специфическим аффинитетом к ДНК. Компетентность зависит от фазы роста бактериальной культуры, вида бактерий. Может увеличиваться при обработке культуры раствором хлорида кальция при пониженной температуре.
Трансформация является основным методом генной инженерии, используемым при конструировании рекомбинантных штаммов с заданным геномом.
2. Трансдукция – передача бактериальной ДНК посредством бактериофага. 1952г. Дж. Ледебург и Н. Циндер (опыты с сальмонеллами)
· Общая (неспецифическая) трансдукция – перенос бактериофагом фрагмента любой части бактериальной хромосомы. Феномен неспецифической трансдукции может быть использован для картирования бактериальной хромосомы.
● Специфическая трансдукция – фаговая ДНК интегрирует в бактерию с образованием профага. При исключении ДНК фага из бактериальной хромосомы захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы. Специфическая трансдукция может служить механизмом переноса вирулентных генов среди бактерий при условии, что эти гены локализованы в непосредственной близости от мест интеграции профага.
● Абортивная трансдукция. При абортивной трансдукции внесенный фрагмент ДНК донора не встраивается в хромосому реципиента, а остается в цитоплазме и там самостоятельно функционирует. Впоследствии он передается одной из дочерних клеток (т.е. наследуется однолинейно) и затем теряется в потомстве.
3. Конъюгация – однонаправленный перенос генетического материала (хромосомной и плазмидной ДНК) от донора к реципиенту при непосредственном контакте клеток.
Донор – бактерия, содержащая конъюгативную плазмиду или конъюгативный транспозон, способная передавать ДНК реципиенту путем горизонтального переноса генов.
Реципиент – любая бактериальная клетка, способная принимать ДНК от бактерии-донора путем горизонтального переноса генов.
Механизм передачи конъюгативных плазмид между Гр- бактериями:
1. Образование межклеточного контакта — конъюгативных пилей
2. Сайт-специфическое разрезание одной цепочки ДНК
3. Сборка особой структуры — релаксосомы для переноса ДНК
4. Перемещение разделенных цепей ДНК в клетку реципиента
5. Синтез комплементарных цепей ДНК
6. Терминация переноса ДНК и разделение конъюгирующих клеток
Биологическое значение конъюгации хорошо видно на примере распространения резистентности бактерий к антибиотикам. Устойчивость к антибиотикам бактерия может получить в результате мутации, что происходит 1 раз на каждые 106 клеточных делений. Изменившаяся генетическая информация может быстро распространяться среди сходных бактерий посредством конъюгации.
1.Цели и задачи МГК. Показания к направлению.
Цель медико-генетической консультации – установление степени генетического риска в обследуемой семье и разъяснение супругам результатов медико-генетического заключения.
Генетический риск – это вероятность появления в потомстве наследственной патологии. Различают низкую степень риска – до 5 %, среднюю степень – до 10 %, повышенную – до 20 % и высокую степень риска – больше 20 %. В зависимости от тяжести медицинских и социальных последствий данной патологии (например, синдром Дауна и синдром Патау) средняя, повышенная и высокая степени риска являются показаниями для прерывания беременности. Решение о прерывании беременности принимают супруги. Врач дает только рекомендации.
В задачи медико-генетических консультаций также входят:
— консультирование семей и больных с наследственной патологией;
— пренатальная диагностика врожденных и наследственных заболеваний;
— помощь врачам других специальностей в постановке диагноза, если необходимы генетические методы исследования;
— ведение территориального регистра семей с наследственной и врожденной патологией и их диспансерное наблюдение;
— постоянная пропаганда медико-генетических знаний среди населения.
Показания для направления семьи в медико-генетическую консультацию:
— наличие наследственной патологии у нескольких членов семьи;
— первичное невынашивание беременности;
— отставание ребенка в физическом и умственном развитии;
— рождение первого ребенка с пороками развития;
— первичная аменорея (отсутствие месячных) при недоразвитии вторичных половых признаков;
— кровное родство супругов.
2.
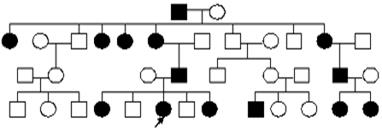
Сцепленный с х хромосомой доминантный тип наследования.
1.Кишечная амеба, Entamoeba coli сходна по морфологии с дизентерийной. Ее локализация — просвет толстого кишечника человека. Образует трофозоиты и цисты. Зрелые цисты кишечной амебы (размеры 13–25 мкм) содержат 8 ядер. Трофозоиты не выделяют протеолитические ферменты и не повреждают стенку кишечника. Непатогенна.
Царство – Protista
Подцарство – Protozoa
Тип – Sarcomastigophora
Подтип – Sarcodina (от греч. sarcodes – состоящий из мяса).
Надклассу – Rhizopoda (корненожек).
Класс – Lobosea
Подкласс — Gimnamoebia
Отряд – Amoebina (т.е. с лопастевидными псевдоподиями «голые» амебы).
Семейство – Endamoebidae
Род – Entаmоeba
Вид — Entаmоeba histolytica
2. Ixodes persulcatus таежный клещ- временные эктопаразиты и специфические переносчики возбудителей трансмиссивных болезней человека и животных (более 20 бактериальных и вирусных инфекций), поддерживают природные очаги чумы, бруцеллеза и туляремии
Токсономия
Царство – Animalia
Подцарство –Metazoa
Тип – ARTHROPODA
Класс — ARACHNOIDEA
Отряды: Acarina
Семейства – Ixodidae
Морфологические особенности : размеры тела от 2 до 30 мм. Отсутствуют дорзальный щиток и глаза, ротовой аппарат расположен вентрально и не виден со спинной стороны. Тело имеет краевой рант.
3. лентец широкий Diphyllobothrium latum — биогельминт, возбудитель дифиллоботриоза. Очаги этого заболевания встречаются по всему земному шару, но приурочены к местностям с крупными водными бассейнами.
Царство – Animalia
Подцарство –Metazoa
Тип – Plathelminthes
Класс — CESTODA
Род – — Diphyllobothrium
Вид — Diphyllobothrium latum
Морфологические особенности: длина тела достигает 10–18 м. На сколексе есть две присасывательные щели — ботрии, которыми паразит прикрепляется к слизистой оболочке кишечника. Размер проглоттид в ширину больше, чем в длину (рис. 37). Зрелые проглоттиды содержат открытую розетковидную матку.
4. самец анкилостомы — Кривоголовка, Ancylostoma duodenale — геогельминт, возбудитель анкилостомоза. Заболевание распространено в странах с субтропическим и тропическим климатом.
Царство – Animalia
Подцарство –Metazoa
Тип – NEMATHELMINTHES
Класс — NEMATODA
Род – Ancylostoma
Вид — Ancylostoma duodenale
Морфологические особенности (рис. 46): самка имеет длину 10–13 мм, самец — 8–10 мм. Тело красноватого цвета. На головном конце находится воронкообразная ротовая капсула с четырьмя кутикулярными зубцами. Задний конец тела самца имеет копулятивную сумку, напоминающую по форме колокол, состоящую из двух больших боковых лопастей и маленькой средней.
5. лямблия интестиналис — возбудитель лямблиоза. Паразитирует только у человека. Заболевание распространено повсеместно.
Морфологические особенности:имеет грушевидную форму с заостренным задним концом (рис. 10). Размер тела 10–18 мкм. На переднем конце имеется 4 пары жгутиков. Есть 2 опорных стержня (аксостили), разделяющие тело на 2 симметричные половины, имеющие по 1 ядру и присасывательному диску. Питание осмотическое, размножение бесполое (продольное деление надвое). Цисты овальной или округлой формы, размером 10–14 мкм.
Вид — Lamblia intestinalis

Билет
1. Специфичность – уникальная последовательность нуклеотидов для каждого структурного гена.
2. Целостность – как функциональная единица (программирование синтеза белка) ген неделим.
3. Дискретность – в составе гена имеются субъединицы: мутон – субъединица, которая отвечает за мутацию; рекон – отвечает за рекомбинацию. Минимальная их величина – пара нуклеотидов.
4. Стабильность – гены относительно устойчивы (стабильны). Частота самопроизвольной мутации одного гена составляет примерно 1 : 10-5 на поколение.
5. Лабильность – устойчивость генов не абсолютная, они могут изменяться, мутировать.
6. Плейотропия – множественное действие гена (один ген отвечает за несколько признаков).
7. Экспрессивность – степень фенотипического проявления гена. Она определяется факторами среды и влиянием других генов.
8. Пенетрантность – частота проявления гена: отношение (в процентах) числа особей, имеющих данный признак, к числу особей, имеющих данный ген.
По функциям гены классифицируют на структурные и функциональные. Структурные гены содержат информацию о белках-ферментах, гистонах, о последовательности нуклеотидов в разных видах РНК. Функциональные гены оказывают влияние на работу структурных генов. Функциональными являются гены-модуляторы и гены-регуляторы. Гены-модуляторы – это ингибиторы, интенсификаторы, модификаторы. Они усиливают, ослабляют или изменяют работу структурных генов. Регулируют работу структурных генов гены-регуляторы и гены-операторы.
Генотип всех соматических клеток организма одного вида одинаков. Но клетки разных тканей отличаются друг от друга. Вероятно, это связано с тем, что в них работают разные блоки генов. Область проявления действия данного гена называется полем его действия (например, распределение волосяного покрова на теле человека). Как правило, гены, детерминирующие
определенные признаки, «работают» непостоянно (например, гены,
определяющие синтез половых гормонов); их функция значительно снижается
с возрастом. Период функционирования гена называется временем его действия.
По месту действия гены подразделяют подразделяют на три группы:
1) функционирующие во всех клетках;
2) функционирующие в клетках одной ткани;
3) специфичные для одного типа клеток.
Первичными функциями генов являются хранение и передача наследственной информации. Гены выполняют в клетке две основные функции.
Гетеросинтетическая функция – это программирование биосинтеза белка в клетке.
Аутосинтетическая функция – репликация ДНК (самоудвоение ДНК).
Репликация ДНК проходит в синтетический период интерфазы митоза. Синтез молекулы ДНК – полуконсервативный: одна цепочка материнская («старая»), на ней собирается новая цепочка – «дочерняя». Новая цепочка собирается по принципу комплементарности. Основной фермент
синтеза – ДНК – полимераза (А. Коренберг, 1956).
Спираль ДНК раскручивается и делится на две цепочки, каждая выполняет роль матрицы. Репликация начинается сразу в нескольких точках молекулы ДНК. Участок ДНК от точки начала одной репликации до точки начала другой называется репликоном. Хромосомы эукариот имеют много репликонов, нуклеоид бактерий – 1 репликон. ДНК – полимераза в репликоне может двигаться вдоль материнской нити только в направлении от конца 3′ к концу 5′. Поэтому сборка дочерних нитей ДНК идет антипараллельно – в противоположных направлениях. Процесс во всех репликонах проходит одновременно. Участок репликации называется репликационной вилкой. В
каждой вилке новые цепи ДНК одновременно собирают несколько ДНК – полимераз.
В каждой репликационной вилке ДНК-полимераза может постепенно и непрерывно собирать одну новую цепь ДНК (так как она движется в одном направлении). Вторая цепь – дочерняя – синтезируется короткими участками по 150-200 нуклеотидов под действием ДНК-полимеразы, которая движется в обратном направлении. Эти участки называются фрагментами Оказаки. Все
синтезированные участки полинуклеотидной цепи соединяются ферментом лигазой.
Весь геном клетки реплицируется 1 раз в митотическом цикле.
2. Гаметогенез – процесс образования половых клеток. В гаметогенезе различают 4 стадии: размножения, роста, созревания и формирования. В стадии размножения происходит деление гаметогоний митозом. В стадии роста гаметоциты I порядка достигают размеров, характерных для половых клеток. В стадии созревания после редукционного деления образуются гаметоциты II порядка, а после эквационного – яйцеклетки и сперматиды, преобразующиеся в сперматозоиды.
Отличия сперматогенеза и овогенеза:
1. При сперматогенезе более интенсивно и продолжительно идет размножение. За время половой жизни мужчины образуется около 500 млрд. сперматозоидов. На 5-ом месяце эмбриогенеза в зачатке женской половой железы насчитывается 6 млн. овогоний. От момента полового созревания до прекращения гаметогенеза созревает 300-400 овоцитов.
2. Стадия роста более выражена при овогенезе, что связано с накоплением желтка.
3. Неравномерное распределение цитоплазмы при мейозе (стадия созревания) характерно для овогенеза. В при сперматогенезе из 1 гаметотоцита I-го порядка образуются 4 сперматиды, при овогенезе – 1 яйцеклетка и 3 направительных (редукционных) тельца.
4. Стадия формирования выражена только при сперматогенезе: сперматиды приобретают форму, характерную для сперматозоидов данного вида.
Сперматозоиды имеют мелкие размеры и подвижны. Они состоят из головки, шейки и хвостика. В головке находится ядро с гаплоидным набором хромосом и небольшое количество цитоплазмы с КГ (акросома). В цитоплазме шейки сосредоточены центросома и митохондрии, вырабатывающие энергию для движения. ЯЦО – 1:0,5.
Яйцеклетка крупная, круглая и неподвижная, имеет оболочку, большое количество цитоплазмы, все органоиды, кроме клеточного центра и гаплоидное ядро. В цитоплазме содержатся желток –питательные вещества, необходимые для развития зародыша. ЯЦО – 1: 500.
• изолецитальные (желтка мало, он равномерно располагается в клетке) – у ланцетника и млекопитающих;
• резко телолецитальные (желтка много, он расположен на вегетативном полюсе клетки, а на анимальном полюсе располагаются цитоплазма и ядро) – у рептилий и птиц;
• умеренно телолецитальные у рыб и земноводных;
• центролецитальные (желтка мало, он располагается в центре клетки) – у насекомых.
3. Голандрический тип наследования.
— болеют только мужчины;
— у больного отца больны все сыновья.
Бычий цепень

Царство – Animalia
Подцарство – Metazoa
Тип – Plathelminthes
Класс – Cestoda
Род – Taeniarhynchus
Вид — Taeniarhynchus saginatus
Мед.значение возбудитель тениаринхоза
Диагностические признаки: зрелый членик бычего невооруженного цепня, членик имеет разветвленную матку,боковых разветвлений от 17-35
2.Малярийный плазмодий
Царство – Protista
Подцарство – Protozoa
Тип – APICOMPLEXA
Класс — Sporozoa (от spora – семя)
Отряд — Coccidiida (истинные кокцидии)
Подотряду – Haemosporina
Род – Plasmodium
Вид – Plasmodium vivax
Мед.значение: малярия
Диагностические признаки:стадия амебоидного шизонта
3.Лейшмания 
Царство – Protista
Подцарство – Protozoa
Тип – Sarcomastigophora
подтипу Mastigophora
классу Zoomastigophora
отряду Kinetoplastida
семейству Leischmaniatidae
Род Leischmania
Вид — Leischmania donovani — висцеральным лейшманиозом
Вид – Leischmania tropica — кожный лейшманиоз
Вид – Leischmania Mexicana — кожный лейшманиоз Нового Света
Вид – Leischmania brasiliensis — кожно-слизистого лейшманиоза Нового Света
Мед.значение:
Висцеральный лейшманиоз (черная болезнь, лихорадка дум-дум, кала-азар, детский\лейшманиоз). Кожный лейшманиоз (пендинка, восточная язва).
Диагностические признаки:
есть 2 формы — промастигота (имеет жгутик, отходящий от кинетопласта, и размеры до 10–20 мкм) и амастигота (безжгутиковая округлая или овальная форма, размер — 3–5 мкм). Все возбудители лейшманиоза морфологически сходны, но имеют биохимические и антигенные различия.
4.Яйцо аскариды
Царство – Animalia
Подцарство –Metazoa
Тип – NEMATHELMINTHES
Класс – NEMATODA
Род – Ascaris
Вид — Ascaris lumbricoides
Мед.значение:геогель-минт, возбудитель аскаридоза.
Диагностические признаки:Диагностика кишечного аскаридоза — обнаружение яиц аскарид в фекалиях. Яйца (60 × 45 мкм) овальной или округлой формы, покрыты тремя оболочками. Наружная оболочка бугристая
5.Головная вошь

Царство -Animalia
Вид — Pediculus humanus capitis
Мед.значение:педикулез
Диагностические признаки:Задний конец тела самца закруглен, у самки — раздвоен. Ротовой аппарат колюще-сосущего типа.
Билет-14
1.Уровни упаковки генетического материала (нуклеосомный супернуклеосомный хроматидный метафазной хромосомы)
Ответ: Упаковка ДНК включает несколько уровней
1) Первый уровень заключается в образовании нуклеосомной нити. Нуклеосома состоит из октамера белков-гистонов, обвитого 2 витками ДНК. Октамер (греч. octo – восемь, meros — часть) состоит из 8 глобулярных белков-гистонов (парные Н2А, Н2В, Н3, Н4). Нуклеосомы разделены короткими участками линкерной ДНК. С октамером и частично с участком линкера связывается гистон H1. Цепь нуклеосом называется нуклеосомной нитью. Она имеет диаметр
10нм. ДНК укорачивается в 6-7 раз.
2) Второй уровень упаковки – образование хроматиновой фибриллы. При этом нуклеосомная нить закручивается в спираль диаметром 30 нм. ДНК укорачивается в 40 раз. В дальнейшей упаковке хроматина ведущую роль играют негистоновые белки.
3) Третий уровень упаковки ДНК – хромомерный. Хроматиновые фибриллы образуют петли – хромомеры, диаметром около 100-150 нм. ДНК укорачивается в 600 раз. Нитчатая хроматиновая структура со средней толщиной 100-200 нм получила название хромонемы. Хромонему можно наблюдать на стадии начальной конденсации хромосом в профазе митоза и при деконденсации хромосом в телофазе.
4) Четвертый уровень — хромосомный. Заключается в более плотной упаковке петель и образовании «конденсированных участков хромосом» диаметром около 700 нм. Максимальной степени конденсации хромосомы достигают в метафазе митоза/меойза.
2. Формы размножения, их характеристика. Эволюция форм полового процесса.
Размножение – свойство живых организмов воспроизводить себе подобных. Выделяют две основные формы размножение: бесполое и половое. Бесполое размножение способствует сохранению наибольшей приспособленности в ней меняющихся условиях обитания, т.к. образуются генетически точные копии родителей.
Клеточные источники наследственной информации-многоклеточные: одна или несколько соматических клеток родителя. Одноклеточные: клетка организма как целое. Родители – одна особь. Потомство – генетически точная копия родителей при отсутствии соматических мутаций,т.е. клон организмов. Главный клеточный механизм — митоз.
Эволюционное значение – поддержание максимальной приспособленности к мало меняющимся условиям обитания. Усиливает роль стабилизирующего естественного отбора.
Половое – Клеточные источники наследственной информации:родителям образуют половые клетки, специализированные к выполнению функций размножения.
Родитель представлен в потомке исходно одной клеткой. Родители – как правило две особи.
Потомство – генетическое отличие от обоих родителей. Главный клеточный механизм – Мейоз.
Эволюционное значение – за счёт генетического разнообразия создает предпосылки к освоению разнообразных условий обитания. Усиливает творческую роль естественного отбора.
Бесполое размножение. Деление путём митоза лежит в основе бесполого размножения организмов. Деление надвое приводит к возникновению из одного родительского организма двух дочерних. Оно является преобладающей формой и у прокариот и простейших. У прокариот – это деление нуклеотида с последующим делением клетки.
Половое размножение. Половое размножение характеризуется наличием полового процесса, при котором происходит слияние гаплоидных половых клеток (гамет). Гаметы образуются в результате особого вида деления клеток – Мейоз. Половой процесс одноклеточных происходит по типу конъюгации и копуляция.
Конъюгация – это половой процесс, заключающийся во временном объединение двух особей и обмене частями их ядерного аппарата,а также небольшим количеством цитоплазмы.
Копуляция – это половой процесс у одноклеточных организмов, при котором две особи приобретают половые различия, т.е. превращаются в гаплоидные гаметы в результате размножения мейозом.
3. 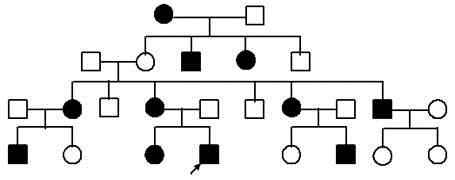
Тип наследования: аутосомно доминантный

 — АА,Аа остальные аа
— АА,Аа остальные аа
4 1.Циста лямблии 
Царство – Protista
Подцарство – Protozoa
Тип – Sarcomastigophora
подтипу Mastigophora
классу Zoomastigophora
отряду Diplomonadida
семейству Hexamitidae.
Род — Lamblia (Giardia)
Вид — Lamblia (Giardia) intestinalis
Мед.значение: возбудитель лямблиоза.
Диагностические признаки: круглое формы, и он внутри имеет жгутики, 2 оболочки
2.Трипаносома
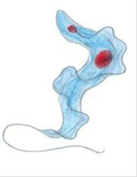
Царство – Protista
Подцарство – Protozoa
Тип – Sarcomastigophora
классу Zoomastigota
Род-трипоносома
вид-трипоносома бруцеи
Мед.значение:трипаносомозы(американский,африканский). Сонная болезнь
Диагностические признаки: Тело изогнутое, сплющенное в одной плоскости, сужено на обоих концах, имеет жгутик, который идет по краю ундулирующей мембраны
3.Аскарида
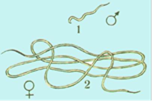
Ascaris lumbricoides.-лат.
Царство – Animalia
Подцарство –Metazoa
Тип – NEMATHELMINTHES
Класс – NEMATODA
Род – Ascaris
Вид — Ascaris lumbricoides
Мед.значение:
миграционный аскаридоз,кишечного аскаридоза
Диагностические признаки:Длина самки достигает 40 см, самца — 25 см. Живые паразиты бело-розового цвета. Тело цилиндрическое, заостренное на концах. На переднем конце тела есть кутикулярные губы.
4.Вошь платяная

Царство -Animalia
Вид – Pediculus HUMANUS HUMANUS
Мед.значение: педикулез, осложнение: колтун-поражение волосистой части головы, вшивый сыпной тиф
Диагностические признаки: более крупные размеры, чем у головной вши (до 4,7 мм), менее глубокие вырезки по краю брюшка и слабо выраженную пигментацию
5.Эхинококк
Царство – Animalia
Подцарство –Metazoa Тип – Plathelminthes
Вид — Echinococcus granulosus
Мед.значение: возбудитель эхинококкоза.
Диагностические признаки: половозрелая форма имеет длину 3–5 мм. Сколекс снабжен присосками и хоботком с двумя рядами крючьев. Шейка короткая. Стробила состоит из 3–4 проглоттид. Предпоследняя проглоттида гермафродитная, последняя — зрелая. Матка разветвленная, закрытая, содержит до 5000 яиц (рис. 35)
1
1 этап. Транскрипция ДНК. На транскрибируемой цепи ДНК с помощью ДНК-зависимой РНК-полимеразы достраивается комплементарная цепь мРНК. Молекула мРНК является точной копией нетранскрибируемой цепи ДНК с той разницей, что вместо дезоксирибонуклеотидов в ее состав входят рибонуклеотиды, в состав которых вместо тимина входит урацил.
2 этап. Процессинг (созревание) мРНК. Синтезированная молекула мРНК (первичный транскрипт) подвергается дополнительным превращениям. В большинстве случаев исходная молекула мРНК разрезается на отдельные фрагменты. Одни фрагменты –- интроны– расщепляются до нуклеотидов, а другие – экзоны – сшиваются в зрелую мРНК. Процесс соединения экзонов «без узелков» называется сплайсинг.
Сплайсинг характерен для эукариот и архебактерий, но иногда встречается и у прокариот Существует несколько видов сплайсинга. Сущность альтернативного сплайсинга заключается в том, что одни и те же участки исходной мРНК могут быть и интронами, и экзонами. Тогда одному и тому же участку ДНК соответствует несколько типов зрелой мРНК и, соответственно, несколько разных форм одного и того же белка. Сущность транс–сплайсинга заключается в соединение экзонов, кодируемых разными генами (иногда даже из разных хромосом), в одну зрелую молекулу мРНК.
3 этап. Трансляция мРНК. Трансляция (как и все матричные процессы) включает три стадии: инициацию (начало), элонгацию (продолжение) и терминацию (окончание).
Транскрипция
В ядре клетки на одной из цепей ДНК (кодирующий) синтезируется и-РНК с участием фермента РНК-полимеразы. Она «переписывает» порядок расположения нуклеотидов в молекуле ДНК (по правилу комплементарности). Этот процесс называется транскрипцией. Единица транскрипции эукариот называется транскриптоном. Он состоит из неинформативной зоны и информативной зоны. Неинформативная, или акцепторная, зона включает промотор, инициатор и блок генов-операторов. Информативная зона содержит структурный ген, который заканчивается терминатором. В составе структурного гена находятся интроны – неинформативные участки ДНК и экзоны – информативные участки. Работу транскриптона регулирует блок генов-регуляторов. На основе их информации синтезируется несколько белков-репрессоров, которые блокируют гены-операторы. Так же как и у оперона, считывание информации со структурного гена происходит, когда в клетку попадают индукторы. В данном случае индукторами являются вещества сложного строения (например, гормоны), для расщепления которых необходимо несколько ферментов. Индукторы освобождают гены-операторы от белков-репрессоров. На одной из нитей ДНК по принципу комплементарности синтезируется и-РНК, но она снимает информацию со всего транскриптона и представляет собой про-и-РНК. В ядре под действием экзо- и эндонуклеаз происходит процессинг про-и-РНК – разрушение неинформативной части и расщепления ее на фрагменты. И-РНК, соответствующая экзонам, образуется в результате сплайсинга (сшивания) информативных участков ферментами лигазами. После таких преобразований и-РНК поступает в цитоплазму на рибосомы, где синтезируются белки-ферменты для разрушения индукторов. Когда индукторы разрушены, восстанавливается блокирование генов-операторов белками-репрессорами, и транскриптон выключается.
2.
Причины развития врожденных пороков:
— генетические (различные мутации);
— экзогенные (действие факторов среды);
— мультифакториальные (совместное действие факторов 1-й и 2-й групп);
Эмбриопатии возникают в период с 16-го дня по 10-ю неделю после оплодотворения. В этот период образования зачатков всех важных органов (органогенез) происходит усиленный процесс дифференцировки клеток и тканей; зародыш обладает высокой чувствительностью к действию повреждающих факторов. Под их влиянием возникают пороки развития (истинные уродства), задержка развития или гибель зародыша. Беременность нередко завершается выкидышем, рождением ребенка с уродствами или мертворож-дением.
Фетопатии — заболевания и функциональные расстройства, возникающие у плода под влиянием экзогенных воздействий в период с 11-й недели беременности до рождения. В зависимости от патогенеза и этиологического фактора различают инфекционные и неинфекционные фетопатии.
Инфекционные фетопатии возникают под влиянием любого инфекционного агента (от вирусов до простейших и грибов) и приводят к врожденному сифилису, туберкулезу, герпетическим инфекциям).
Неинфекционные фетопатии (гипотрофия и гипоксия) развиваются в результате плацентарной недостаточности и обменных нарушений у плода при патологии беременности. К неинфекционным фетопатиям относятся гемолитическая болезнь плода; врожденный генерализованный кальциноз артерий, фибробластоз миокарда, диабетическая, тиреотоксическая, алкогольная фетопатии.
3.
Ответ:
-Необходимо рекомендовать использование других методов пренатальной диагностики: серологическое обследование, исследование содержания других эмбрионспецифических белков, ультрасонографию, амниоцентез с последующим биохимическим и цитогенетическим исследованием клеток плода.
-Повышенный уровень a-фетопротеина может быть как при патологии плода (угрожающий выкидыш, внутриутробная гибель плода, резус-несовместимость матери и плода, грыжа пупочного канатика, дефекты нервной трубки, врожденный нефроз, крестцово-копчиковая тератома), так и при многоплодной беременности.
Источник



