
Способы откладки яиц насекомых




Фаза яйца
Типы яиц и способы кладки. Яйцо насекомого представляет собой крупную клетку (рис. 46, А) и, помимо протоплазмы и ядра, содержит также дейтоплазму, или желток, необходимый для питания и развития зародыша. Помимо того, в яйце насекомых нередко находятся симбиотические микроорганизмы, получаемые от матери трансовариально, т. е. через яичники.
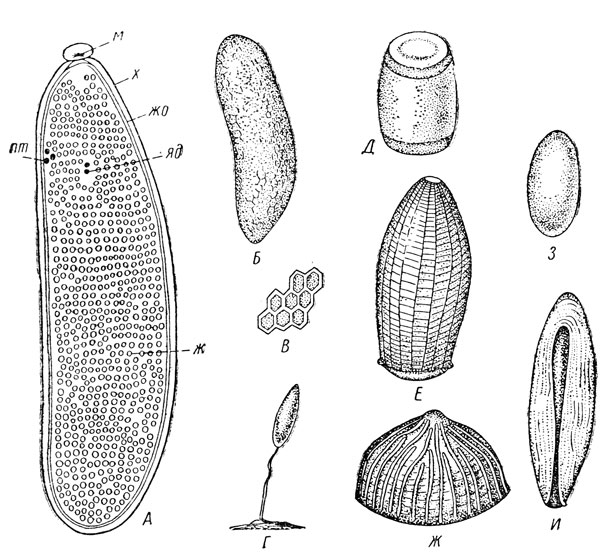
Рис. 46. Яйцо и его типы. А — строение яйца мухи; Б — яйцо саранчового; В — участок хориона яйца саранчового при сильном увеличении; Г — яйцо листоблошки; Д — клопа; Е — бабочки-белянки; Ж — бабочки-совки; З — жука-листоеда; И — капустной мухи (по Богданову-Катькову, Иммсу и др.): м — микропиле, х — хорион, жо — желточная оболочка, яд — ядро, пт — полярные тельца, ж — желток
Яйцо снаружи покрыто хорионом — оболочкой, возникшей за счет выделений фолликулярного эпителия. Хорион нередко имеет явственную микроскульптуру, которая часто очень характерна и тогда может служить надежным признаком для различения родов и даже видов насекомых по фазе яйца. Под хорионом лежит истинная, или желточная, оболочка яйца, но возможны и другие окружающие яйцо образования. На поверхности хориона часто обнаруживается микропиле — отверстие, служащее для прохождения сперматозоидов при оплодотворении; нередко около микропиле скульптура хориона усложняется. В яйце есть передний и задний концы, которые соответствуют положению зародыша; в яичнике передний полюс яйца направлен к голове матери, соответственно располагается спинная и брюшная сторона яйца.
Размеры и внешний вид яйца весьма разнообразны (рис. 46, Б, Г-И). У некоторых тлей, трипсов, мельчайших перепончатокрылых яйца имеют всего 0,02-0,03 мм длины, тогда как у крупных саранчовых — могут достигать 8-10 мм и более. Внешне они могут быть гладкими, без макроскульптуры (саранчовые, тли и др.), либо с резкой ребристой или иной скульптурой (совки и белянки из чешуекрылых, палочники и др.); в некоторых случаях они снабжены крышечкой (клопы-щитники). По форме яйца могут быть овальными (многие жуки и бабочки), удлиненными (саранчовые, кузнечики, мухи), полушаровидными (совки), бутылковидными (дневные бабочки), боченковидными (некоторые клопы) и пр., в ряде случаев яйцо может быть снабжено стебельком или ножкой (у медяниц, сетчатокрылых — золотоглазок, хальцид и др.).
Откладываются яйца также разнообразно — поодиночке, группами, открыто или погруженными в субстрат, либо защищенные различными способами. Наиболее обычный тип открытой кладки — откладка яиц на поверхности листьев и других частей растений; при этом яйца прикреплены к субстрату выделениями придаточных половых желез. При закрытой кладке яйца оказываются погруженными в ткани растений или находятся в почве. Защищенная кладка характеризуется образованием какого-либо общего прикрытия или общей оболочки. Так, самка непарного шелкопряда группу отложенных яиц перемешивает с войлочком из снятых с конца брюшка волосков, а яблонная моль покрывает группу яиц на ветке яблони щитком из выделений придаточных половых желез. У тараканов и богомоловых яйца заключены в яйцевую капсулу — оотеку, образовавшуюся из секрета придаточных желез. У большинства саранчовых яйца закладываются группой в почву и сопровождающий секрет не только обволакивает их, но может цементировать прилегающие частицы почвы; образуется своеобразная защищенная кладка — кубышка (рис. 86).
Таким образом, для насекомых характерно большое разнообразие типов яиц и способов их откладки. Это делает возможным определение по фазе яйца многих групп насекомых до семейства, рода и даже вида.
Эмбриональное развитие. Эмбриональное развитие начинается с дробления ядра и передвижения образовавшихся дочерних ядер с небольшими участками протоплазмы к периферии яйца. Здесь из массы дочерних ядер образуется сплошной слой клеток — бластодерма. В дальнейшем бластодерма дифференцируется на зародышевую и внезародышевую зоны; клетки последней не участвуют в образовании зародыша, тогда как клетки зародышевой зоны начинают делиться более интенсивно и образуют с брюшной стороны яйца зародышевую полосу.
Продолжающееся размножение клеток бластодермы приводит к впячиванию срединной части зародышевой полосы с последующим смыканием над местом впячивания наружного его слоя. Так возникает дифференциация этой полосы на зародышевые слои — эктодерму и мезодерму; что касается внутреннего зародышевого слоя, или энтодермы, то у крылатых насекомых он образуется за счет оставшихся в яйце неиспользованными ядер дробления. Таким образом, у насекомых начальный этап дробления яйца не сопровождается образованием морулы; объясняется это богатством яйца насекомых инертным желтком, не участвующим в дроблении яйца.
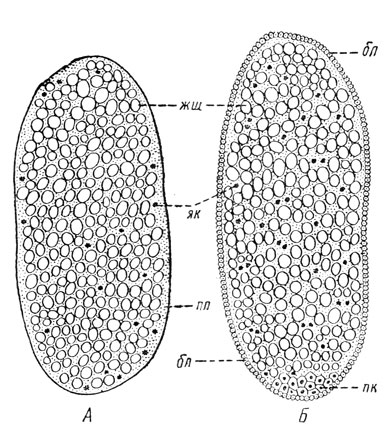
Рис. 47. Дробление яйца жука-листоеда. А — начало дробления; Б — в конце дробления (по Иммсу); жш — желточные шары, пл — плазма, як — клетки, возникшие из дробящегося ядра, бл — бластодерма, пк — половые клетки
Развитие зародыша сопровождается также бластокинезом, образованием зародышевых оболочек и сегментацией (рис. 48). Бластокинез представляет собой перемещение зародыша к новым, еще не усвоенным участкам желтка в яйце. Протекает он практически одновременно с образованием зародышевых оболочек; по А. Г. Шарову, у насекомых наблюдаются два основных способа бластокинеза. У ортоптероидных (Orthopteroidea) и у насекомых с полным превращением (Holometabola) зародыш разрастается по длине яйца, но его головная часть остается в передней части яйца; спереди и сзади внезародышевая бластодерма образует над зародышем нарастающую складку. Обе складки сливаются и образуют над зародышем две зародышевые оболочки — наружную, или серозу, и внутреннюю, или амнион. Зародыш оказывается под защитой названных оболочек и даже изолируется от контакта с ними. Несомненно, это создает более надежные условия для нормального развития эмбриона, особенно если яйца оказываются отложенными открыто, т. е. в виде незащищенной кладки.
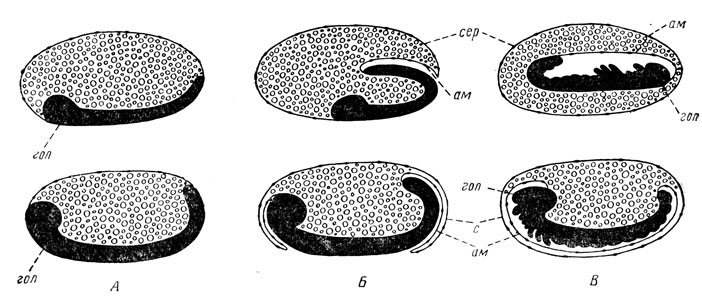
Рис. 48. Способы образования зародышевых оболочек и бластокинез у насекомых (по Шарову). А — зародыш до образования оболочек; Б — начальный этап их образования; В — завершение образования зародышевых оболочек. Верхний ряд — древнекрылые и гемиптероидные насекомые, нижний ряд — ортоптероидные и с полным превращением: гол — головной отдел зародыша, ам — амнион, с — сероза
У древнекрылых (Palaeoptera) и гемиптероидных (Hemipteroidea) указанной выше нарастающей складки бластодермы не образуется, а у зародыша хвостовой отдел загибается внутрь яйца, погружается в желток и увлекает за собой весь зародыш. Головной отдел его оказывается в задней части яйца и здесь же происходит смыкание зародышевых оболочек; сам зародыш оказывается глубоко погруженным в желток яйца.
Примерно одновременно с разрастанием зародыша и образованием его оболочек начинается его сегментация. Вначале на головном отделе появляются зачатки глаз в виде пары выступов. В области будущей переднегруди возникает центр сегментации; от него вперед отчленяются головные сегменты, а назад — сегменты груди и брюшка. С брюшной стороны этих сегментов затем появляются выросты — зачатки парных конечностей: на голове — усиков и ротовых частей, на груди и немного позже на брюшке — зачатки ног. При этом в головном отделе обнаруживается явственная сегментация, но вопрос о числе сегментов остается спорным, так как одни авторы насчитывают их 5, а другие 4 или иное число.
Как показал еще в 1913 г. итальянский ученый А. Берлезе, развитие сегментации и конечностей эмбриона сопровождается прохождением трех стадий: протоподной, полиподной и олигоподной (рис. 49). Протоподная стадия характеризуется отсутствием или слабой сегментацией, а парные придатки слабо намечены лишь на голове и груди; полиподная стадия отличается уже явственной сегментацией брюшка и появлением на нем зачатков конечностей, которые в заключительной олигоподной стадии исчезают, сохраняясь лишь на голове и груди. Однако эта схема имеет отклонения; например, у многих насекомых с полным превращением выпадает полиподная стадия, у некоторых паразитических перепончатокрылых существует лишь первая, т. е. протоподная стадия и пр.
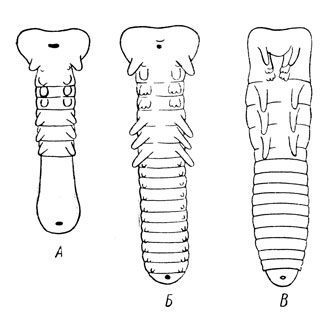
Рис. 49. Три стадии развития конечностей и сегментации зародыша: А — протоподная; Б — полиподная; В — олигоподная (из Шванвича)
Основы внутреннего строения начинают закладываться с образованием мезодермы и возникновением зародышевых слоев. Из эктодермы происходят все наружные покровы тела, затем путем впячивания образуются ротовое и анальное отверстия с зачатками передней и задней кишки, зачатки будущих трахей, концевые части половой системы и продольный тяж клеток — зачаток нервной системы. Энтодерма имеет вначале вид узкого срединного тяжа, но в последующем на его переднем и заднем конце появляются направленные друг к другу выступы, которые затем разрастаются, сливаются вместе и образуют замкнутую трубку, слившуюся с зачатками передней и задней кишки и содержащую захваченный при разрастании желток. Так образуется средняя кишка, точнее ее эпителий. Содержащийся в ней эмбриональный желток идет на построение органов зародыша, но не всегда полностью расходуется, и его остатки могут сохраниться в кишечнике вылупившейся личинки. Мезодерма разрастается в пространстве между эктодермой и энтодермой и дает начало мышечной системе, включая и мышечный слой кишечника, жировому телу, спинному сосуду, оболочке половых желез.
Что касается развития собственно половых желез, т. е. яичников и семенников, то первоначально появляются клетки полового зачатка, которые обособляются еще на ранних стадиях развития яйца — иногда еще до дифференциации на зародышевые слои. В последующем эти зачатки половых клеток мигрируют в соответствующие части тела зародыша, покрываются клетками мезодермы и превращаются в половые железы. Столь раннее обособление полового зачатка свидетельствует об особых свойствах половых клеток как носителях наследственных свойств.
Вполне сформировавшийся зародыш заполняет все яйцо, часто характеризуется потемнением глаз, а также иногда и концов ротовых частей и готов к вылуплению; в сущности, это уже личинка. Она начинает совершать интенсивные движения, набирает в трахеи воздух, заглатывает амниотическую жидкость и тем самым увеличивает объем своего тела. Наконец личинка выходит из яйца наружу — происходит вылупление. При этом вылупляющаяся личинка прогрызает оболочку яйца — хорион, либо разрезает или пробуравливает ее специальным органом — пиловидным образованием на голове, шипом и пр.
У некоторых видов развившаяся в яйце осенью личинка остается в нем на зимовку; вылупление в этих случаях происходит весной, что обеспечивает личинке более благоприятные возможности для перезимовки и последующей жизни и питания. Зимовка сформировавших личинок в яйце наблюдается у непарного шелкопряда, некоторых совок и др.
В большинстве случаев развитие насекомого в фазе яйца продолжается недолго — от нескольких дней (у многих мух) до двух-трех недель. Но нередко продолжительность фазы яйца может быть более значительной, достигая 6-9 месяцев; это происходит в тех Случаях, когда яйца откладываются осенью и уходят на зимовку, либо при возникновении эмбриональной диапаузы — временной остановки развития зародыша.
Источник
Типы яиц насекомых, способы их кладки.
Яйцо насекомых представляет собой крупную клетку. протоплазма (заполняет всю внутреннюю часть яйца), снаружи покрыто хорионом – оболочкой, которая предохраняет от различных повреждений. Под харионом лежит желточная оболочка на поверхности хориона обнаруживается микропиле – (отверстие служащее для оплодотворения
2. Размеры и внешний вид яйца очень разнообразны:
У тлей, трипсов, филоксеры от 0,02 – 0,03 мм длины, у саранчи 8-10 мм., у кузнечиков до 15 мм., 8-10 мм. у медведки. По форме:
1. Серповидные — саранча
2. Боченковидные — клопы
3. Веретеновидные — мухи
4. Бутылковидные — белянки
5. Полушаровидные — совки
6. Со стебельком — у златоглазки, медяниц
7. Лепешковидные — луговой мотылек. По окраске: цвет зависит от вида насекомого. Свежеотложенные яйца имеют молочно-белую окраску, а спустя некоторое время приобретают окраску свойственную данному виду: оранжевая – колорадский жук, желтоватые яйца у капустной белянки, белые у мухи. Откладывают яйца так же разнообразно:
1. Поодиночке – яблонная плодожорка, пилильщики
2. Группами или кучками — колорадский жук
Если яйца откладываются определенными группами, то такие группы называются кладками яиц.
Кладки бывают: 1. Правильными рядами . (крестоцветные клопы). 2. В виде колечек на однолетних побегах: кольчатый шелкопряд.
Кладки бывают открытого типа – на поверхности листьев и других частях растений (колорадский жук)
Прикрываются яйца пушком из брюшка самки — непарный шелкопряд, златоглазка.
В форме кубышки — саранча.
Собранные в мошковидную оболочку – или особую капсулу — оотеку (тараканы).
Насекомые могут откладывать разное количество яиц, которое зависит от вида насекомого и от условий: плодовитость ком. От 200 до 3000 штук; медведка от 150 до 300, тля от 2 до 10 яиц.
80. свекловичная блошка и меры борьбы с ней. Свекловичные блошки — cистематическое положение: отряд жуки, или жесткокрылые, семейство листоеды (Chrysomelidae).
На свекле вредят несколько видов блошек, имеющих сходные особенности развития, но различные зоны вредоносности и спектр повреждаемых культур. Обыкновенная свекловичная блошка — Chaetocnema concinna Marsh, распространена повсеместно, круг кормовых растений помимо свеклы включает лен, хмель, различные виды маревых, гречишных и капустных. Южная свекловичная блошка — СЛ. breviuscula Fald. повреждает свеклу и маревые сорняки в Центрально-Черноземном, Поволжском, Северо-Кавказском, Уральском и Западно-Сибирском регионах. На юге европейской части России свекле вредит западная свекловичная блошка — СЛ. tibiales 111. Широко распространенная корнеплодная блошка — Psylloides cupreata Duft. повреждает свеклу, брюкву и турнепс в Западно-Сибирском и Дальневосточном регионах.
Жуки свекловичных блошек мелкие, длиной 1,5—2,5 мм; тело овальное, выпуклое, черное с различными металлическими оттенками: от медно-бронзового до сине-зеленого; антенны 11-члени- ковые; задние ноги прыгательные. Видовые отличия блошек основаны на строении переднеспиыки и пунктировке головы.
Зимуют имаго на поверхности почвы под растительными остатками. Обычные места зимовок — краевые участки лесополос и другой древесно-кустарниковой растительности. Весной перезимовавшие жуки начинают дополнительное питание на маревых и гречишных сорняках, позже заселяют всходы свеклы. В этот период блошки наносят наиболее серьезные повреждения растениям, выгрызая на листьях многочисленные мелкие ямки и сквозные отверстия. Листовая пластинка при этом подсыхает и крошится. Вредоносность жуков особенно усиливается в жаркую сухую погоду и часто приводит к массовой гибели всходов свеклы.
После спаривания самки откладывают по 4—12 яиц в поверхностный слой почвы вблизи кормовых растений. Плодовитость блошек составляет 200—240 яиц. Эмбриональный период длится 11 — 13 дней, после чего отродившиеся червеобразные личинки начинают питаться на корнях маревых и гречишных растений, не нанося им существенного вреда. Личинки развиваются не более 1 мес, окукливаются свекловичные блошки в почве. Вылетевшие в конце лета жуки недолго питаются на сорных и культурных растениях, после чего переселяются в места зимовки. У обыкновенной и корнеплодной блошек за год развивается одно поколение, в южных регионах у западной и южной свекловичных блошек может развиваться второе поколение.

На численность свекловичных блошек оказывают влияние многие виды паразитических нематод, вызывающие гибель личинок или значительное снижение плодовитости у самок. В популяциях блошек часто отмечаются массовые эпизоотии, вызываемые энтомопатогенными грибами.
Источник



