
Способы опыления голосеменных растений




4.4. Опыление у голосеменных
Опыление у голосеменных занимает как бы промежуточное положение между простым рассеиванием (микро-)спор у низших растений и собственно опылением. Как и в случае опыления, возможности прорастания ограниченны: если пыльцевое зерно не достигнет микропиле совместимого семязачатка в женском стробиле, то оно не сможет прорасти и образовать мужские половые клетки.
Из-за отсутствия рыльца воспринимающей поверхностью «женского цветка» голосеменных является микропиле семязачатка или примыкающая к нему чешуйка стробила. Мы не располагаем данными, свидетельствующими о том, что эти воспринимающие поверхности избирательно взаимодействуют с пыльцевым зерном или с пыльцевой трубкой. Следовательно, несовместимость у голосеменных, если она вообще существует, обнаруживается на гаплоидной стадии: между ядрами или между мужским ядром и женским эндоспермом, а не при взаимодействии гаплоида и диплоида, столь важном в системах несовместимости у покрытосеменных. Если рассматривать опыление в широком смысле слова, включая в него как опыление у голосеменных, так и опыление у покрытосеменных * , то приходится признать, что оно имеет полифилетическое происхождение, т. е. что переход от микроспоры к пыльце происходил независимо в различных развивающихся линиях сосудистых споровых, в том числе и тех, которые дали начало современным высшим растениям: голосеменным и покрытосеменным.
* ( Это мы считаем вполне допустимым, но возражаем против идеи включить также перенос микроспор у сосудистых споровых. В макроспорангии имеются структуры, очень похожие, например, на рыльце (см. Maheshwari, 1960), но их происхождение совершенно иное. Это, конечно, не означает, что невозможны одинаковые механизмы переноса.)
Поскольку хвойные преобладают в количественном отношении по сравнению с остальными голосеменными и поскольку для хвойных характерно, если не присуще им исключительно, ветроопыление, можно легко прийти к неверному выводу о том, что все голосеменные опыляются ветром. Исходя из возможности полифилетического происхождения голосеменных, следует не меньшее, а быть может, и большее значение придавать способам опыления в других порядках, помимо хвойных. У Ginkgo такой же способ опыления, как и у хвойных, хотя оплодотворение может произойти даже после отделения макроспорангия.
У саговниковых (Cycadales) по разрозненным наблюдениям по крайней мере некоторые виды опыляют насекомые; пока не ясно, случайные ли это наблюдения или постоянно присущий некоторым видам способ опыления. Известно, что жуки, питающиеся пыльцой, посещают мужские стробилы Encephalartos и Zamia integrifolia (Wester, 1910), а пчелы небольших размеров собирают пыльцу Macrozamia [разновидность М. tridentata (Willd.); Regel по Schuster, 1932]. Поскольку эти растения двудомные (посещение насекомыми только мужских шишек не приводит к опылению), спрашивается, зачем насекомым посещать женские шишки? На стробилах Macrozamia наблюдали представителей полужесткокрылых (клопов). Опылительная капля может быть аттрактантом для мелких насекомых; вместе с тем очень сомнительно, что и подъем температуры, наблюдаемый только в мужских стробилах, может служить аттрактантом в тропиках. У некоторых саговников и мужские, и женские шишки имеют очень сильный запах. У отдельных видов этот запах очень неприятен для человека, и, напротив, он очень приятен у Macrozamia; именно поэтому растения этого рода посещаются пчелами. По данным Рэтри (Rattray, 1913), жуки посещают мужские шишки Encephalartos, привлекаемые запахом, а в женских откладывают яйца. Можно думать, что механизм опыления в данном случае соответствует механизму опыления некоторых покрытосеменных, в котором принимают участие полувредные насекомые, подобные Hadena или Chiastochaeta (см. разд. 8.5). Наконец, следует упомянуть, что Бэйрд (Baird, 1938), хотя и придерживается мнения, что пыльца саговников рассеивается ветром от одного стробила к другому, однако считает, что перенос пыльцы внутрь женской шишки Macrozamia (см. выше) зависит от насекомых, так как при опылении ветром пыльцевое зерно не может проникнуть в микропиле, расположенное слишком глубоко. Насекомых обнаруживали внутри шишек обоих полов. Таким образом, имеется достаточно оснований, чтобы признать определенную роль насекомых в опылении саговниковых, но окончательного вывода сделать нельзя, поскольку никому не приходилось непосредственно наблюдать, как насекомые переносят пыльцу от мужского стробила к женскому. Норстог и Стевенсон (Norstog, Stevenson, 1980) наблюдали, что личинки жуков-долгоносиков живут в мужских шишках (культивируемой) Zamia furfuracea; взрослые насекомые покидают их, будучи при этом покрыты пыльцой. Позже они внедряются между чешуйками женских шишек, куда не проникает ветер. В этом случае опять не ясен тип аттрактации, но факт завязывания большого числа семян вполне очевиден.
Принято считать, что у гнетовых (Gnetales) преобладает ветроопыление. Однако хорошо известно, что у нескольких видов, например у Ephedra campylopoda и Welwitschia, в мужских стробилах имеется один стерильный семязачаток (см. Porsch, 1958; мы не согласны с утверждением Бормана (Bornmann, 1972) о том, что Welwitschia опыляется ветром). Очевидно, его единственная функция — выделение (сладкой) опылительной капли для привлечения насекомых. У этих растений наблюдается совершенно иное явление, чем у саговниковых, у которых возможный аттрактант, а именно пыльца для питающихся пыльцой жуков, находится в мужской шишке, в связи с чем трудно объяснить причину привлечения насекомых к женской шишке. У гнетовых возможный аттрактант, а именно опылительная капля, образуется в женском стробиле, в мужском, по-видимому, нет ничего, что привлекало бы насекомых, и рудиментарный семязачаток выполняет эту функцию. Если принять общее филогенетическое правило, что органы никогда не возникают как рудименты, то следовательно, Ephedra и Welwitschia должны были произойти от растений с гермафродитными стробилами, которые позднее стали однополыми, подобно цветкам многих других ветроопыляемых растений. Некоторые виды Ephedra (см. Mehra, 1950) и Welwitschia остались более примитивными и до сих пор (частично?) опыляются насекомыми (Ephedra отмечена в работе Ordetz, 1952). Мужские стробилы Е. campylopoda обладают запахом фруктов. Эти примеры еще раз свидетельствуют о том, что опыление насекомыми должно быть более древним, чем разделение полов между отдельными особями и, возможно, также, что последнее появилось в ответ на вторичное ветроопыление, как это было у многих высших растений. По некоторым наблюдениям, у Gnetum насекомые собирают липкую пыльцу на ярких сладко пахнущих ночью мужских стробилах. В женских стробилах этого растения образуются сладковатые опылительные капельки (см. Hendryck, 1953). Несомненно, что было бы крайне интересно найти в стробилах этой группы голосеменных такие же свидетельства первичного гермафродитизма и опыления насекомыми, однако при этом следует соблюдать большую осторожность и не допускать поспешных обобщений; хвойные весьма отдаленно связаны с Gnetales, и их однополость может быть первичной, например однополые стробилы развились у их предков до того, как появились другие признаки, выделившие эти растения в голосеменные. Возможная первичная энтомофилия у саговниковых и гнетовых не исключает первичной анемофилии у гинкго и хвойных — быть может, единственной первичной анемофилии. В конце концов филогенетические связи между этими группами и покрытосеменными так неясны, что в отношении последней группы можно делать какие-либо заключения только по аналогии, но есть основания полагать, что образование нектара, заключенного в нектарниках цветка покрытосеменных, не первично.
Во всяком случае ясно, что проблемы, связанные с выяснением функций опылительной капли, не так просты, как кажется. По словам Мак-Уильямса (McWilliams, 1958), «поглощение микропилярной жидкости под воздействием пыльцы трудно объяснить». Дойл (Doyle, 1943) утверждает, что опылительная капля образуется у примитивных хвойных, тогда как у более продвинутых типов (Abies, Tsuga) воспринимающая функция переходит к выступам микропиле. Это может означать, что в процессе филогенеза данной группы закрепилась анемофилия, а первоначально существовавшая энтомофилия была утрачена. Однако Дойл и Кэм (Doyle, Кат, 1944) пришли к заключению, что нуцеллус недоступен для переносимой ветром пыльцы A. homolepis. Поэтому в процессе оплодотворения возникает пыльцевая трубка-посредник, растущая на поверхности семенной чешуи.
Источник
Голосеменные
Спешу обрадовать, мы добрались до изучения семенных растений! К ним относятся голосеменные и покрытосеменные (цветковые). До этого размножение шло только с помощью спор: у мхов, папоротников, хвощей и плаунов — высших споровых растений. Настало время открыть новую интересную главу этой книги, посвященную растениям, которые размножаются с помощью удивительного изобретения природы — семени.
Голосеменные — распространенная древняя группа растений, включающая небольшое число видов. Главной особенностью данной группы являются «голо» (то есть открыто) лежащие семяпочки и, в дальнейшем, развивающиеся из них семена. Иными словами, у голосеменных растений отсутствуют замкнутые вместилища для семян.

На примере типичного представителя — сосны обыкновенной, относящейся к классу хвойных, поговорим о характерных чертах данного класса и голосеменных растений в целом.
Общие признаки
Все голосеменные представлены древесными формами: деревьями и кустарниками. Травы отсутствуют.
Хвоинки (хвоя) — игольчатые видоизменения листьев. Сохраняются долгие годы, у некоторых сосен до 45 лет. Хвоя лиственниц опадает ежегодно.

Древесина голосеменных обладает большим запасом механической прочности. Это связано с ее особенностями: она состоит из трахеид с окаймленными порами, паренхима развита слабо. Либриформ (древесные волокна) и настоящие сосуды отсутствуют (исключение — гнетовые, имеют сосуды). Клетки-спутницы во флоэме также отсутствуют.
В древесине и коре имеются каналы, заполненные смолой. Однако, есть исключения — у гинкго смола не образуется вовсе.
Несколько веков назад в России целенаправленно создавались и охранялись, так называемые, корабельные рощи. Это, прежде всего, требовалось для флота, так как мачты кораблей изготавливали из сосен, отвечающих всем требованиям — корабельных (гладкий, твердый и прочный прямой ствол с минимальным количеством сучков и смолы).

Семяпочки и развивающиеся из них семена лежат «голо», открыто, для них нет закрытых вместилищ, отсутствует завязь. В сравнении с высшими споровыми растениями, размножение семенами ставит голосеменных на более высокий уровень организации.
Голосеменным растениям для размножения не требуется вода, опыление у них происходит с помощью ветра. Этот процесс перестал быть зависимым от капельно-жидкой среды, как было у мхов и у папоротников. Благодаря этому голосеменные получили большое преимущество и смогли расселиться по всей Земле, в том числе в засушливых районах. Они господствовали в юрском периоде, когда климат стал более сухим и жарким.

Обитают голосеменные в местах с холодным климатом и достаточным количеством влаги. Имеются виды, обитающие в жарких странах: растение вельвичия удивительная обитает в пустынях южной Африки.
Строение и жизненный цикл
Жизненный цикл голосеменных состоит из чередования бесполого поколения — спорофита (диплоиден, 2n), и полового поколения — гаметофита (гаплоиден, n). Господствует (доминирует) в цикле спорофит (2n) — это взрослое растение сосны.
Голосеменные относятся к разноспоровым, как и все семенные растения. Они образуют разные споры: крупные женские (мегаспоры) и мелкие мужские (микроспоры). Образуются они в спорангиях, расположенных на спорофиллах, которые собраны в стробилы (шишки) — от лат. strobilus — сосновая шишка.
Мужские шишки (стробилы)
К концу весны у основания молодых побегов образуются мужские шишки (стробилы) — мелкие, собранные в тесные группы, желтого цвета. Чешуи мужских шишек представляют собой микроспорофиллы. Микроспорофиллы — гомологи тычинок, которые крепятся к оси каждой шишки спирально, с нижней стороны, и имеют два пыльцевых мешка — микроспорангия.
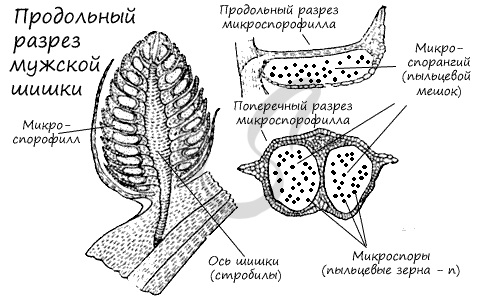
Образование мужского гаметофита
Из материнских клеток (2n) в микроспорангии путем мейоза образуются 4 микроспоры (n). Строение микроспоры следующее: она покрыта экзиной (от гр.exo снаружи, вне) — наружная оболочка, изнутри интиной (от лат. intus внутри) — внутренней оболочкой. В составе микроспоры имеются также два воздухоносных мешка, образованных в результате отслоения экзины от интины и возникновения полости между ними.
Микроспора делится, не покидая спорангия, преобразуется в заросток. При делении из ядра микроспоры образуются две клетки. Одна из них превращается в две заростковые клетки (протоллиальные — от греч. проталлиум — заросток) — быстро отмирают и исчезают. Их функция до конца не изучена.
Из другой клетки в ходе митоза также образуются две: антеридиальная, из которой развиваются мужские половые клетки — спермии (неподвижные, без жгутиков в отличие от сперматозоидов), и более крупная вегетативная клетка, из которой в дальнейшем формируется пыльцевая трубка.

Мужской гаметофит сильно упрощен, антеридии отсутствуют. Формируется он прямо внутри микроспоры, которая в итоге превращается в пыльцевое зерно. Совокупность пыльцевых зерен называется пыльца.
При вскрытии (нарушении целостности) микроспорангия, или пыльцевого мешка, пыльца высыпается во внешнюю среду и достигает женской шишки, где, в результате опыления, внутри семязачатка происходит дальнейшее развитие мужского гаметофита.
Образование женского гаметофита
На тех же самых соснах, где расположены мужские шишки, лежат и женские. Весной на верхушке молодого побега появляются мелкие (около 5 мм) красноватые шишки — это женские шишки (стробилы). Состоят они из оси (стержня) , на котором располагаются две чешуи: кроющая и семенная. На верхней стороне у основания семенной чешуи лежат два семязачатка.
Кроющая чешуя представляет собой видоизмененный лист, в его пазухе находится семенная чешуя. Семенная чешуя — видоизмененный боковой побег.
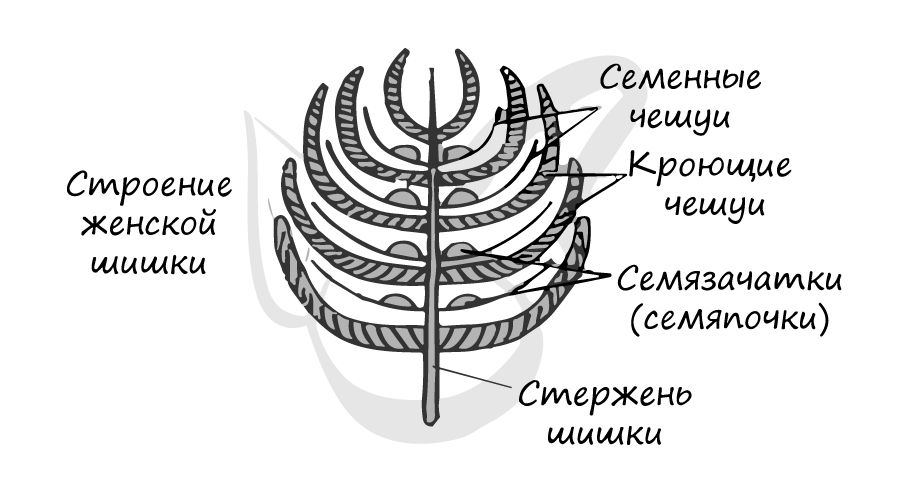
Женские шишки (стробилы)
Именно открыто расположенные семязачатки (семяпочки) служат причиной, по которой этот отдел растений называется — голосеменные.
В женских шишках, в отличие от мужских, каждая чешуя гомологична целой мужской шишке (стробилу). То есть одна чешуя — целой мужской шишке, а не отдельным ее микроспорофиллам (чешуям)!
Молодой семязачаток состоит из нуцеллуса, интегумента и фуникулуса. Нуцеллус (от лат. nucella — орешек) — центральная часть семяпочки, соответствующая мегаспорангию. Интегумент (от лат. integumentum покрывало) — покров семяпочки, вырастающий из ее центральной части — нуцеллуса. В зрелом семени интегумент преобразуется в семенную кожуру. Фуникулус (от лат. funiculus канатик, верёвка) или семяножка — часть семязачатка, соединяющая его с мегаспорофиллом (семенным чешуями).
На интегументе около вершины располагается микропиле (пыльцевход) — через него после опыления пыльцевая трубка проникает в нуцеллус. Между интегументом и нуцеллусом имеется густая жидкость, выступающая из микропиле. Подсыхая, она втягивается внутрь семязачатка и затягивает вместе с собой пыльцу, осевшую на ней.
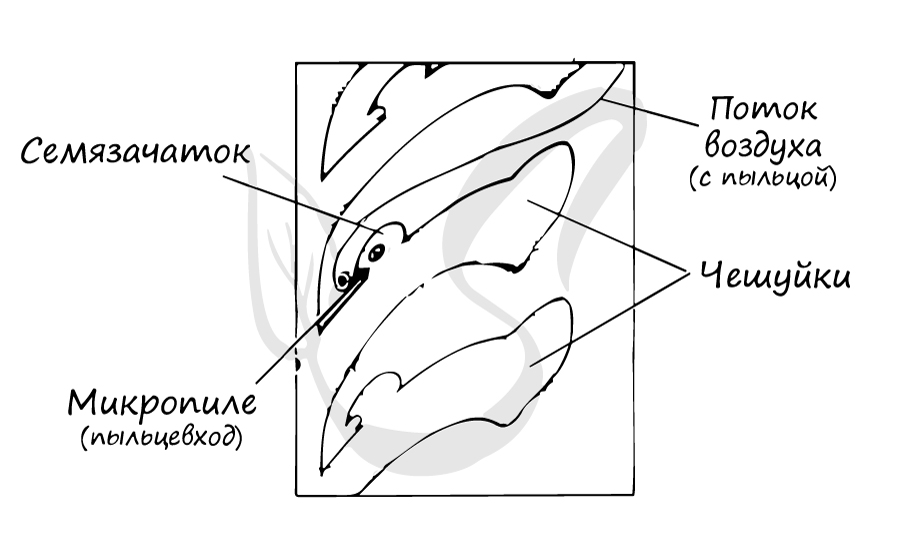
Образование женского гаметофита
В средней части обособляется спорогенная клетка (2n) (археспориальная — от греч. arche начало и sporá семя). В результате ее митотического деления образуются материнские клетки спор — спороциты (2n), однако и сама археспориальная клетка может выступать в роле спороцита, минуя стадию митоза. Спороциты (2n) делятся мейозом на четыре гаплоидные (n) мегаспоры.
Три мегаспоры отмирают, остается одна, которая многократно делится митозом и формирует эндосперм — запасное питательное вещество. Обратите на этот факт особое внимание: у голосеменных эндосперм гаплоидный (n) и образуется до оплодотворения. Такой эндосперм называется — первичный, он соответствует женскому гаметофиту.
Как и мужской, женский гаметофит весьма упрощен и заключен внутри мегаспоры. На верхушке женского гаметофита (мегагаметофита) образуется архегоний с яйцеклеткой (n). У гнетовых архегонии отсутствуют.

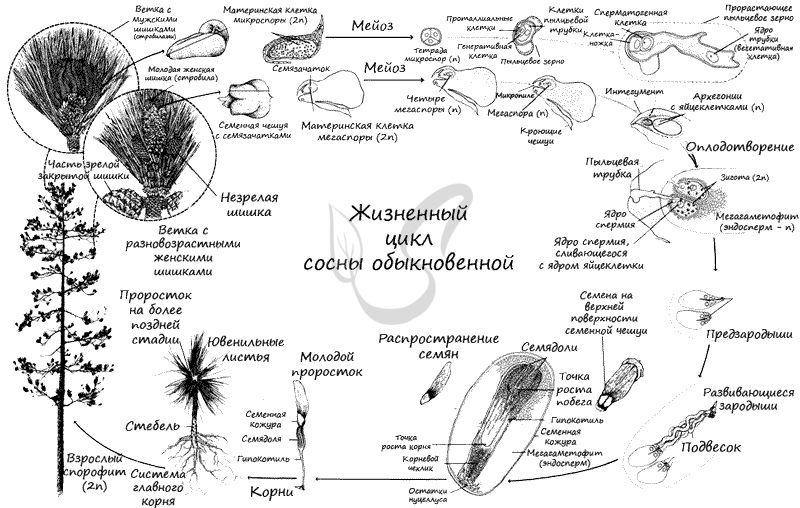
Жизненный цикл
На спорофите (2n) в микроспорангиях из материнских клеток (2n) путем мейоза образуются микроспоры (n). Из микроспоры формируется пыльцевое зерно. Пыльца (пыльцевые зерна (n)) с помощью ветра попадает в женские шишки, где улавливается густой жидкостью между интегументом и нуцеллусом, выступающей из микропиле. Жидкость засасывает пыльцу внутрь семязачатка на нуцеллус (в пыльцевую камеру). После того, как опыление произошло, микропиле зарастает. Чешуи шишки смыкаются и склеиваются смолой.
Семязачатки в этот момент еще не готовы к оплодотворению, так что от момента опыления до оплодотворения проходит около 13 месяцев. За это время в семязачатке формируется эндосперм, женская шишка увеличивается до 3-4 см и приобретает зеленую окраску.
Оказавшись на мегаспорангии, наружная оболочка пыльцевого зерна (экзина) разрывается, из вегетативной клетки в направлении архегония начинает расти пыльцевая трубка. Антеридиальная клетка делится на генеративную (спермагенную) и клетку-ножку антеридия (функция последней до сих пор не изучена). Спермагенная клетка попадает в пыльцевую трубку, а из нее — в архегоний.
Непосредственно перед оплодотворением спермагенная клетка делится на два спермия (n), один из которых отмирает, а другой сливается с яйцеклеткой (n). Образуется зигота (2n), из которой формируется и растет зародыш благодаря эндосперму — запасу питательных веществ.
Окончательно созревают семена к осени на второй год после опыления, к этому моменту женские шишки увеличиваются в размерах до 6 см. Зеленая окраска меняется на серую, чешуйки расходятся, и семена, образовавшиеся из семязачатков, высыпаются. Из семени прорастает взрослое растение — спорофит (2n). Цикл замыкается.
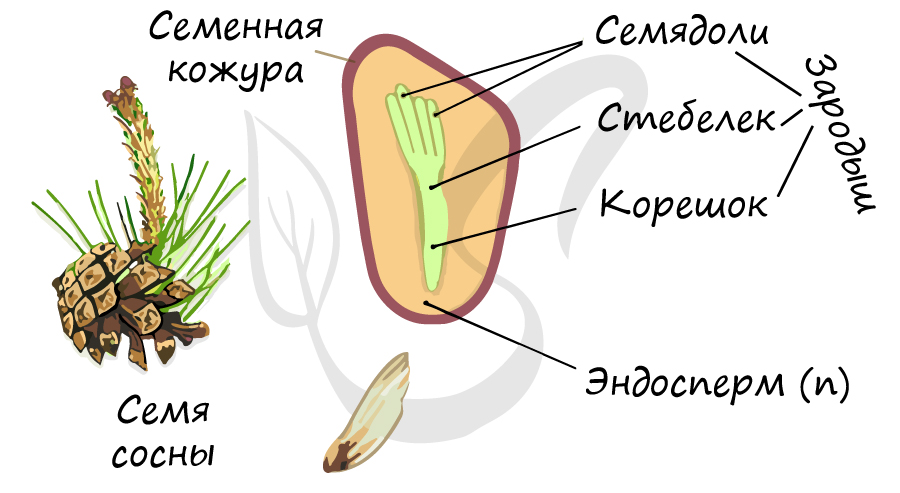
Строение семени
Семя голосеменных состоит из:
- Семенной кожуры
Семенная кожура, защищающая семя от пересыхания и неблагоприятных факторов внешней среды, образована разросшимся интегументом.
Зародыш (2n) формируется в результате митотического деления образовавшейся зиготы. Состоит из зародышевого корешка, стебелька и почечки.
Число семядолей у голосеменных различается — от 2 до 15. Семядоли имеют доступ к запасным питательным вещества (эндосперму).
Запас питательных веществ
Запасные питательные вещества накапливаются в эндосперме (n). Особенностью в строении семени голосеменных, по сравнению с семенем покрытосеменных (цветковых) является наличие гаплоидного эндосперма (n). Не забывайте, что эндосперм у голосеменных это производное мегагаметофита (n), исходя из этого становится понятно, почему ткань гаплоидна. У цветковых, в отличие от голосеменных, эндосперм триплоиден (3n).
Фитонциды
Фитонциды (от греч. phyton — растение и лат. caedo — убиваю) — образуемые растениями, биологически активные вещества, убивающие или приостанавливающие размножение других организмов, главным образом — микробов. Обычно выделяются растениями в газообразном виде, к примеру, аллицин у лука и чеснока. Наличие фитонцидов играет крайне важную роль в формировании устойчивости растения к грибным заболеваниям.
Фитонциды имеют медицинское значение, из них изготавливаются некоторые препараты. За лето гектар лиственного леса выделят 2 кг фитонцидов, хвойного — 5 кг, можжевельника — 30 кг! Санатории часто располагаются в сосновых борах, где наблюдается повышенная концентрация фитонцидов. Вдыхание такого воздуха очень полезно при заболеваниях дыхательной системы инфекционной природы (когда возбудителями являются бактерии, грибы).

Значение голосеменных
Трудно переоценить значение голосеменных для человека, они очень важны. Голосеменные — источники высококачественной древесины, продуктов ее переработки. Являются звеном в цепи питания (продуцентами), основой многих биоценозов. Хвойные растения в больших количествах выделяют фитонциды, имеющие медицинское значение. Из смолы хвойных получают канифоль, скипидар, лаки. Кедровых орехи — это семена нескольких видов растений из рода сосна, которые употребляют в пищу.

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Источник



