
Регистрация потенциала покоя
Как можно обнаружить мембранный потенциал покоя?
1. методом повреждения
2. методом внутриклеточного отведения (рис. ).
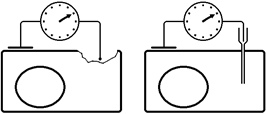
Рис. . Методы регистрации мембранного потенциала покоя: слева – метод повреждения, справа – метод внутриклеточного отведения[Б34] .
Студенты на этот вопрос часто отвечают так: методами внутриклеточного и внеклеточного отведения. Типичная ошибка! Методом внеклеточного отведения можно зарегистрировать возбуждение участка мембраны, сравнив его потенциал с потенциалом мембраны в состоянии покоя, но не потенциал покоя. Но об этом позже.
Метод повреждения чаще используется при регистрации потенциала покоя на макропрепарате – мышце, нерве (рис. ).
Рис. . Схема регистрации мембранного потенциала покоя методом повреждения на макропрепарате.
Впервые таким способом регистрировали потенциал покоя в середине XIX века Дюбуа-Реймон (на срезе нерва[Б35] ) и Маттеучи (на срезе мышцы). Этот же метод лежит в основе второго опыта Гальвани[Б36] , в этом случае мышечно-нервный препарат выполняет роль регистрирующего прибора.
Методом повреждения достаточно точно измерить потенциал покоя невозможно.
Чтобы измерить потенциал покоя и тем более проследить его изменения, вызываемые каким-либо воздействием на клетку, применяют технику внутриклеточных микроэлектродов (рис. ).
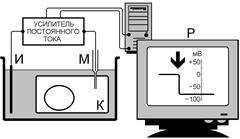
Рис. . Схема измерения потенциала покоя клетки с помощью внутриклеточного электрода.
К – объект исследования – клетка, М – микроэлектрод, И – индифферентный электрод, Р – регистрирующее устройство. Момент прокола на кривой «разность потенциалов – время» показан стрелкой.
Микроэлектрод представляет собой микропипетку, т.е. тонкий капилляр, вытянутый из стеклянной трубочки. Диаметр его кончика около 0,5 мкм[Б37] . Микропипетку заполняют солевым раствором (обычно 3 М KС1), погружают в него металлический электрод (хлорированную серебряную проволочку) и соединяют с электроизмерительным прибором[Б38] , снабженным усилителем постоянного тока[Б39] .
Микроэлектрод устанавливают над исследуемым объектом, например скелетной мышцей, а затем при помощи микроманипулятора — прибора, снабженного микрометрическими винтами, вводят внутрь клетки[Б40] . При удачном введении микроэлектрода мембрана плотно охватывает его кончик и клетка сохраняет способность функционировать в течение нескольких часов, не проявляя признаков повреждения[Б41] .
Микроэлектрод является активным (референтным). Электрод сравнения (индифферентный) обычных размеров погружают в нормальный солевой раствор, в котором находится исследуемая ткань[Б42] .
До прокола мембраны микроэлектродом разность потенциалов между активным и индифферентным электродом равна нулю (рис. и рис. А). Kак только микроэлектрод прокалывает поверхностную мембрану клетки, регистрируется разность потенциалов между поверхностью и содержимым клетки, равная потенциалу покоя клетки[Б43] . На рис. показано, что продвижение микроэлектрода внутри протоплазмы (рис. B, C, D) или изменение положения электрода внутри клетки (рис. E) на показаниях вольтметра не сказываются[Б44] . После прокола (рис. F) микроэлектродом мембраны на противоположной стенке клетки от входа в клетку электрода разность потенциалов вновь не регистрируется. Это свидетельствует о том, что разница потенциал действительно определяется между цитоплазмой и окружающим клетку наружным раствором[Б45] .
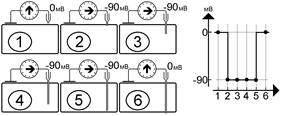
[Б47] Рис. . Результаты измерения разности потенциала микроэлектродным методом при разном расположении активного электрода.
У различных клеток мембранный потенциал покоя варьирует от –50 до –90 мВ[Б48] .
Источник
Вопрос 1 Мембранный потенциал или потенциал покоя- методы регистрации, величина, природа.




![]()
![]()
МП, или потенциал покоя, — это разность потенциалов между наружной и внутренней поверхностями мембраны в условиях покоя. В среднем у клеток возбудимых тканей он достигает 50—80 мВ, со знаком «—» внутри клетки. Обусловлен преимущественно ионами калия. Как известно, в клетках возбудимых тканей концентрация ионов калия достигает 150 ммоль/л, в среде — 4—5 ммоль (ионов калия намного больше в клетке, чем в среде). Поэтому по градиенту концентрации калий может выходить из клетки, и это происходит с участием калиевых каналов, часть которых открыта в условиях покоя. В результате из-за того, что мембрана непроницаема для анионов клетки (глутамат, аспартат, органические фосфаты), на внутренней поверхности клетки образуется избыток отрицательно заряженных частиц, а на наружной — избыток положительно заряженных частиц. Возникает разность потенциалов. Эта ситуация описывается уравнением Нернста:
П — ПХ Л* Кв ** (формула другая)
Чем выше концентрация калия в среде — тем меньше это отношение, тем меньше величина мембранного потенциала. Однако расчетная величина, как правило, ниже реальной, Например, по расчетам МП должен быть -90 мВ, а реально -70 мВ. Это расхождение обусловлено тем, что ионы натрия и хлора тоже вносят свой вклад в создание МП. В частности, известно, что натрия больше в среде (140 ммоль/л против 14 ммоль/л внутриклеточной). Поэтому натрий может войти в клетку. Но большая часть натриевых каналов в условиях покоя закрыта. Поэтому в клетку входит лишь небольшая часть ионов натрия. Но и этого достаточно, чтобы хотя бы частично компенсировать избыток анионов. Ионы хлора, наоборот, входят в клетку (частично) и вносят отрицательные заряды. В итоге величина мембранного потенциала определяется в основном калием, а также натрием и хлором. Для того чтобы МП поддерживался на постоянном уровне, необходимо поддержание ионного гетерогенитета — ионной асимметрии. Для этого, в частности, служит калий-на- триевый насос (и хлорный), который восстанавливает ионную асимметрию, особенно после акта возбуждения. Доказательством калиевой природы МП является наличие зависимости: чем выше концентрация калия в среде, тем меньше величина МП. Для дальнейшего изложения важно понятие; деполяризация (уменьшение МП, например, от минус 90 мВ до минус 70 мВ) и гиперполяризация — противоположное явление.
Вопрос 2 Газообмен между альвеолярным воздухом и кровью. Обмен газов как результирующая трех процессов – вентиляции легких, перфузии легких (легочного кровотока) и диффузии газов.
Обмен газов в легких определяется такими факторами как вентиляция легких, перфузия, т. е. интенсивность кровотока через легкие, и процесс диффузии.
Диффузия газов осуществляется через легочную мембрану — это многослойный «пирог», представленный пленкой сурфактанта, эпителием альвеолы, интерстицием (2 основные мембраны), эндотелием капилляра и слоем плазмы. Толщина этой мембраны около 1 мкм. При патологии, например, при отеке легких или при фиброзах легких, толщина мембраны может существенно возрастать, что нарушает процесс диффузии газов. В цепом, у человека площадь легочной мембраны, через которую осуществляется диффузия, в норме составляет 50—80 м2. При резекции легких, при ателектазе, при пневмонии эта площадь значительно уменьшается, что снижает диффузионные процессы.
Процесс диффузии описывается первым законом диффузии Фнка: диффузионный поток, т. е. количество вещества, проходящего через площадь слоя А за единицу времени, — прямо пропорционален градиенту давления (Р, —Р2) и площади А и обратно пропорционален толщине слоя (1).
где К — коэффициент диффузии (коэффициент Крога), или
диффузионная проводимость, который зависит от природы газа. Для углекислого газа он в 20—25 раз выше, чем для кислорода. Закон Фнка показывает факторы, от которых зависит скорость диффузии. В клинической практике широко используется понятие «диффузионная способность легких» (ДСЛ): это отношение объема газа,
продиффундировавшего через легочную мембрану за 1 минуту в расчете на 1 мм рт. ст. градиента давления. Контакт крови с альвеолярным воздухом происходит за 0,3—0,7 с. За этот период времени осуществляется полное выравнивание парциальных давлений (напряжений) газа. Учитывая, что процесс связывания кислорода гемоглобином протекает еще быстрее — за несколько милисекунд, — то процесс диффузии и насыщения крови кислородом и отдача углекислого газа происходит в полном объеме за 0,3—0,7 с. Соотношение между объемом вентиляции и объемом перфузии является важным фактором газообмена. В легких, несмотря на существование альвеолярно-капиллярного рефлекса, регулирующего соотношение между вентилируемыми альвеолами и процессом их перфузии, возникает ситуация, при которой имеет место не только анатомическое мертвое пространство, но и альвеолярное мертвое пространство, т. е. имеются вентилируемые альвеолы, лишенные кровотока. Поэтому при расчете объема вентиляции легких учитывается объем физиологического мертвого пространства (суммы анатомического и альвеолярного мертвых пространств). В норме объем вентиляции легких составляет примерно 0,8 от объема перфузии легких (80%). В положении «лежа» в силу гидростатического давления легкое равномерно снабжается кровью: верхушки, средние области и основание получают примерно одно и то же количество крови. В положении «сидя» верхушки легких снабжаются кровью хуже (примерно на 15% меньше, чем в положении «лежа»), а стоя — на 25% меньше. Таким образом, перфузия легких максимальна в положении «лежа». Поэтому при необходимости увеличить диффузию газов у человека его следует перевести в позу «лежа». Отметим, что наиболее опасным состоянием для газообмена является отек легких: при этом газам приходится проходить через легочную мембрану, насыщенную водой. Известно, что С02 в жидкости диффундирует примерно в 13000 раз, а кислород — в 300000 раз медленнее, чем в газовой среде.

Вопрос 3 Проприоцептивная сенсорная система, или двигательный анализатор. Рецепторный, проводниковый и корковый отделы проприоцептивного анализатора.
Источник
2_1 Мембранный потенциал покоя кратко (доцент Сазонов В.Ф.)
Введение
Зачем нам нужно знать, что такое потенциал покоя?
Что такое «животное электричество»? Откуда в организме берутся «биотоки»? Как живая клетка, находящаяся в водной среде, может превратиться в «электрическую батарейку»?
— На эти вопросы мы сможем ответить, если узнаем, как клетка за счёт перераспределения электрических зарядов создаёт себе электрический потенциал на мембране.
Как работает нервная система? С чего в ней всё начинается? Откуда в ней берётся электричество для нервных импульсов?
— На эти вопросы мы также сможем ответить, если узнаем, как нервная клетка создаёт себе электрический потенциал на мембране.
Итак, понимание того, как работает нервная система, начинается с того, что надо разобраться, как работает отдельная нервная клетка — нейрон.
А в основе работы нейрона с нервными импульсами лежит перераспределение электрических зарядов на его мембране и изменение величины электрических потенциалов. Но чтобы потенциал изменять, его нужно для начала иметь. Поэтому можно сказать, что нейрон, готовясь к cвоей нервной работе, создаёт на своей мембране электрический потенциал, как возможность для такой работы.
Таким образом, наш самый первый шаг к изучению работы нервной системы — это понять, каким образом перемещаются электрические заряды на нервных клетках к как за счёт этого на мембране появляется электрический потенцила. Этим мы и займёмся, и назовём этот процесс появления электрического потенциала у нейронов — формирование потенциала покоя.
Определение
В норме, когда клетка готова к работе, у неё уже есть электрический заряд на поверхности мембраны. Он называется мембранный потенциал покоя.
Потенциал покоя — это разность электрических потенциалов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя. Его средняя величина составляет -70 мВ (милливольт).
«Потенциал» — это возможность, он сродни понятию «потенция». Электрический потенциал мембраны — это её возможности по перемещению электрических зарядов, положительных или отрицательных. В роли зарядов выступают заряженные химические частицы — ионы натрия и калия, а также кальция и хлора. Из них только ионы хлора заряжены отрицательно (-), а остальные — положительно (+).
Таким образом, имея электрический потенциал, мембрана может перемещать в клетку или из клетки указанные выше заряженные ионы.
Важно понимать, что в нервной системе электрические заряды создаются не электронами, как в металлических проводах, а ионами — химическими частицами, имеющими электрический заряд. Электрический ток в организме и его клетках — это поток ионов, а не электронов, как в проводах. Обратите также внимание на то, что заряд мембраны измеряется изнутри клетки, а не снаружи.
Если говорить уж совсем примитивно просто, то получается, что снаружи вокруг клетки будут преобладать «плюсики», т.е. положительно заряженные ионы, а внутри — «минусики», т.е. отрицательно заряженные ионы. Можно сказать, что внутри клетка электроотрицательна. И теперь нам всего лишь надо объяснить, как это так получилось. Хотя, конечно, неприятно сознавать, что все наши клетки — отрицательные «персонажи». ((
Сущность
Сущность потенциала покоя — это преобладание на внутренней стороне мембраны отрицательных электрических зарядов в виде анионов и недостаток положительных электрических зарядов в виде катионов, которые сосредотачиваются на её наружной стороне, а не на внутренней.
Внутри клетки — «отрицательность», а снаружи — «положительность».
Такое положение вещей достигается с помощью трёх явлений: (1) поведения мембраны , (2) поведения положительных ионов калия и натрия и (3) соотношения химической и электрической силы.
1. Поведение мембраны
В поведении мембраны для потенциала покоя важны три процесса:
1) Обмен внутренних ионов натрия на наружные ионы калия. Обменом занимаются специальные транспортные структуры мембраны: ионные насосы-обменники. Таким способом мембрана перенасыщает клетку калием, но обедняет натрием.
2) Открытые калиевые ионные каналы. Через них калий может как заходить в клетку, так и выходить из неё. Он выходит в основном.
3) Закрытые натриевые ионные каналы. Из-за этого натрий, выведенный из клетки насосми-обменниками, не может вернуться в неё обратно. Натриевые каналы открываются только при особых условиях — и тогда потенциал покоя нарушается и смещается в сторону нуля (это называется деполяризацией мембраны, т.е. уменьшением полярности).
2. Поведение ионов калия и натрия
Ионы калия и натрия по-разному перемещаются через мембрану:
2) Через постоянно открытые калиевые каналы калий выходит из клетки, но может и возвращаться в неё обратно через них же.
3) Натрий «хочет» войти в клетку, но «не может», т.к. каналы для него закрыты.
3. Соотношение химической и электрической силы
По отношению к ионам калия между химической и электрической силой устанавливается равновесие на уровне — 70 мВ.
1) Химическая сила выталкивает калий из клетки, но стремится затянуть в неё натрий.
2) Электрическая сила стремится затянуть в клетку положительно заряженные ионы (как натрий, так и калий).
Формирование потенциала покоя
Попробую рассказать коротко, откуда берётся мембранный потенциал покоя в нервных клетках — нейронах. Ведь, как всем теперь известно, наши клетки только снаружи положительные, а внутри они весьма отрицательные, и в них существует избыток отрицательных частиц — анионов и недостаток положительных частиц — катионов.
И вот тут исследователя и студента поджидает одна из логических ловушек: внутренняя электроотрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а наоборот — из-за потери некоторого количества положительных частиц (катионов).
И поэтому сущность нашего рассказа будет заключаться не в том, что мы объясним, откуда берутся отрицательные частицы в клетке, а в том, что мы объясним, каким образом в нейронах получается дефицит положительно заряженных ионов — катионов.
Куда же деваются из клетки положительно заряженные частицы? Напомню, что это ионы натрия — Na + и калия — K + .
Натрий-калиевый насос
А всё дело заключается в том, что в мембране нервной клетки постоянно работают насосы-обменники, образованные специальными белками, встроенными в мембрану. Что они делают? Они меняют «собственный» натрий клетки на наружный «чужой» калий. Из-за этого в клетке оказывается в конце концов недостаток натрия, который ушёл на обмен. И в то же время клетка переполняется ионами калия, который в неё натащили эти молекулярные насосы.
Чтобы легче было запомнить, образно можно сказать так: «Клетка любит калий!» (Хотя об истинной любви здесь не может идти и речи!) Поэтому она и затаскивает калий в себя, несмотря на то, что его и так полно. Поэтому она невыгодно обменивает его на натрий, отдавая 3 иона натрия за 2 иона калия. Поэтому она тратит на этот обмен энергию АТФ. И как тратит! До 70% всех энергозатрат нейрона может уходить на работу натрий-калиевых насосов. Вот что делает любовь, пусть даже не настоящая!
Кстати, интересно, что клетка не рождается с потенциалом покоя в готовом виде. Например, при дифференцировке и слиянии миобластов потенциал их мембраны изменяется от -10 до -70 mV, т.е. их мембрана становится более электроотрицательной, она поляризуется в процессе дифференцировки . А в экспериментах на мультипотентных мезенхимальных стромальных клетках (ММСК) костного мозга человека искусственная деполяризация ингибировала дифференцировку клеток (Fischer-Lougheed J., Liu J.H., Espinos E. et al. Human myoblast fusion requires expression of functional inward rectifier Kir2.1 channels. Journal of Cell Biology 2001; 153: 677-85; Liu J.H., Bijlenga P., Fischer-Lougheed J. et al. Role of an inward rectifier K+ current and of hyperpolarization in human myoblast fusion. Journal of Physiology 1998; 510: 467-76; Sundelacruz S., Levin M., Kaplan D.L. Membrane potential controls adipogenic and osteogenic differentiation of mesenchymal stem cells. Plos One 2008; 3 ).
Образно говоря, можно выразиться так:
Создавая потенциал покоя, клетка «заряжается любовью».
Это любовь к двум вещам:
1) любовь клетки к калию,
2) любовь калия к свободе.
Как ни странно, но результат этих двух видов любви — пустота!
Именно она, пустота, создаёт в клетке отрицательный электрический заряд — потенциал покоя. Точнее, отрицательный потенциал создают пустые места, оставшиеся от убежавшего из клетки калия.
Итак, результат деятельности мембранных ионных насосов-обменников таков:
Натрий-калиевый ионный насос-обменник создаёт три потенциала (возможности):
1. Электрический потенциал — возможность затягивать внутрь клетки положительно заряженные частицы (ионы).
2. Ионный натриевый потенциал — возможность затягивать внутрь клетки ионы натрия (и именно натрия, а не какие-нибудь другие).
3. Ионный калиевый потенциал — возможновть выталкивать из клетки ионы калия (и именно калия, а не какие-нибудь другие).
1. Дефицит натрия (Na + ) в клетке.
2. Избыток калия (K + ) в клетке.
Можно сказать так: ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Именно из-за получившегося дефицита натрия в клетку теперь «полезет» этот самый натрий снаружи. Так всегда ведут себя вещества: они стремятся выравнять свою концентрацию во всём объёме раствора.
И в то же время в клетке получился избыток ионов калия по сравнению с наружной средой. Потому что насосы мембраны накачали его в клетку. И он стремится уравнять свою концентрацию внутри и снаружи, и поэтому стремится выйти из клетки.
Тут ещё важно понять, что ионы натрия и калия как бы «не замечают» друг друга, они реагируют только «на самих себя». Т.е. натрий реагирует на концентрацию натрия же, но «не обращает внимания» на то, сколько вокруг калия. И наоборот, калий реагирует только на концентрацию калия и «не замечает» натрий. Получается, что для понимания поведения ионов в клетке надо по-отдельности сравнивать концентрации ионов натрия и калия. Т.е. надо отдельно сравнить концентрацию по натрию внутри и снаружи клетки и отдельно — концентрацию калия внутри и снаружи клетки, но не имеет смысла сравнивать натрий с калием, как это часто делается в учебниках.
По закону выравнивания концентраций, который действует в растворах, натрий «хочет» снаружи войти в клетку. Но не может, так как мембрана в обычном состоянии плохо его пропускает. Его заходит немножко и клетка его опять тут же обменивает на наружный калий. Поэтому натрий в нейронах всегда в дефиците.
А вот калий как раз может легко выходить из клетки наружу! В клетке его полно, и она его удержать не может. Так вот он и выходит наружу через особые белковые дырочки в мембране (ионные каналы).
Анализ
От химического — к электрическому
А теперь — самое главное, следите за излагаемой мыслью! Мы должны перейти от движения химических частиц к движению электрических зарядов.
Калий заряжен положительным зарядом, и поэтому он, когда выходит из клетки, выносит из неё не только себя, но и «плюсики» (положительные заряды). На их месте в клетке остаются «минусы» (отрицательные заряды). Это и есть мембранный потенциал покоя!
Мембранный потенциал покоя — это дефицит положительных зарядов внутри клетки, образовавшийся за счёт утечки из клетки положительных ионов калия.
Заключение

Рис. Схема формирования потенциала покоя (ПП). Автор благодарит Попову Екатерину Юрьевну за помощь в создании рисунка.
Составные части потенциала покоя
Потенциал покоя — отрицательный со стороны клетки и состоит как бы из двух частей.
1. Первая часть — это примерно -10 милливольт, которые получаются от неравносторонней работы мембранного насоса-обменника (ведь он больше выкачивает «плюсиков» с натрием, чем закачивает обратно с калием).
2. Вторая часть — это утекающий всё время из клетки калий, утаскивающий положительные заряды из клетки. Он дает большую часть мембранного потенциала, доводя его до -70 милливольт.
Калий перестанет выходить из клетки (точнее, его вход и выход сравняются) только при уровне электроотрицательности клетки в -90 милливольт. Но этому мешает постоянно подтекающий в клетку натрий, который тащит с собой свои положительные заряды. И в клетке поддерживается равновесное состояние на уровне -70 милливольт.
Обратите внимание на то, что для создания потенциала покоя нужны затраты энергии. Эти затраты производятся ионными насосами, которые обменивают «свой» внутренний натрий (ионы Na + ) на «чужой» внешний калий (K + ). Вспомним, что ионные насосы являются ферментами АТФазами и расщепляют АТФ, получая из неё энергию на указанный обмен ионов разного типа друг на друга.Тут очень важно понять, что с мембраной «работают» сразу 2 потенциала: химический (концентрационный градиент ионов) и электрический (разность электрических потенциалов по разные стороны мембраны). Ионы перемещаются в ту или иную сторону под действием обеих этих сил, на которые и тратится энергия. При этом один из двух потенциалов (химический или электрический) уменьшается, а другой увеличивается. Разумеется, если рассматривать электрический потенциал (разность потенциалов) отдельно, то не будут учитываться «химические» силы, перемещающие ионы. И тогда может сложиться неверное впечатление о том, что энергия на движение ионо берётся как бы ниоткуда. Но это не так. Необходимо рассматривать обе силы: химическую и электрическую. При этом крупные молекулы с отрицательными зарядами, находящиеся внутри клетки играют роль «статистов», т.к. их не перемещают через мембрану ни химические, ни электрические силы. Поэтому эти отрицательные частицы обычно и не рассматривают, хотя они существуют и именно они обеспечивают отрицательную сторону разности потенциалов между внутренней и наружной сторонами мембраны. А вот шустрые ионы калия, как раз способны к перемещению, и именно их утечка из клетки под действием химических сил создаёт львиную долю электрического потенциала (разности потенциалов). Ведь именно ионы калия перемещают на наружную сторону мембраны положительные электрические заряды, будучи положительно заряженными частицами.
Так что всё дело в натрий-калиевом мембранном насосе-обменнике и последующем вытекании из клетки «лишнего» калия. За счёт потери положительных зарядов при этом вытекании внутри клетки нарастает электроотрицательность. Она-то и есть «мембранный потенциал покоя». Он измеряется внутри клетки и составляет обычно -70 мВ.
Выводы
Говоря образно, «мембрана превращает клетку в «электрическую батарейку» с помощью управления ионными потоками».
Мембранный потенциал покоя образуется за счёт двух процессов:
1. Работа калий-натриевого насоса мембраны.
Новая гипотеза механизма работы Na,K-АТФазы рассматривается здесь: Механизм натрий-калиевого насоса
Работа калий-натриевого насоса, в свою очередь, имеет 2 следствия:
1.1. Непосредственное электрогенное (порождающее электрические явления) действие ионного насоса-обменника. Это создание небольшой электроотрицательности внутри клетки (-10 мВ).
Виноват в этом неравный обмен натрия на калий. Натрия выбрасывается из клетки больше, чем поступает в обмен калия. А вместе с натрием удаляется и больше «плюсиков» (положительных зарядов), чем возвращается вместе с калием. Возникает небольшой дефицит положительных зарядов. Мембрана изнутри заряжается отрицательно (примерно -10 мВ).
1.2. Создание предпосылок для возникновения большой электроотрицательности.
Эти предпосылки — неравная концентрация ионов калия внутри и снаружи клетки. Лишний калий готов выходить из клетки и выносить из неё положительные заряды. Об этом мы скажем сейчас ниже.
2. Утечка ионов калия из клетки.
Из зоны повышенной концентрации внутри клетки ионы калия выходят в зону пониженной концентрации наружу, вынося заодно положительные электрические заряды. Возникает сильный дефицит положительных зарядов внутри клетки. В итоге мембрана дополнительно заряжается изнутри отрицательно (до -70 мВ).
Финал
Калий-натриевый насос создает предпосылки для возникновения потенциала покоя. Это — разность в концентрации ионов между внутренней и наружной средой клетки. Отдельно проявляет себя разность концентрации по натрию и разность концентрации по калию. Попытка клетки выравнять концентрацию ионов по калию приводит к потере калия, потере положительных зарядов и порождает электроотрицательность внутри клетки. Эта электроотрицательность составляет большую часть потенциала покоя. Меньшую его часть составляет непосредственная электрогенность ионного насоса, т.е. преобладающие потери натрия при его обмене на калий.
Видео: Мембранный потенциал покоя (Resting membrane potential)
Источник



