
- Общая характеристика способов обмена генетической информацией у бактерий
- Микра / Лекции / Скорина / Лекция 21
- Горизонтальный обмен генами
- Читайте также
- Перекресты. Расстояния между генами
- Обмен веществ
- Микробам — горизонтальный перенос, высшим организмам — половое размножение
- Горизонтальный перенос генов и симбиоз
- Обмен витаминов
- Глава 18. Обмен гликогена
- Обмен жирных кислот
- Обмен кетоновых тел
- Глава 26. Обмен нуклеотидов
- Обмен железа
- 2.3. Обмен веществ и энергии
- ОБМЕН УГЛЕВОДОВ
- 7.4. Углеродный обмен в биосфере
- 7.6. Азотный обмен
- 16. Обмен веществ и превращение энергии. Энергетический обмен
- 13. Гений, управляющий генами
Общая характеристика способов обмена генетической информацией у бактерий
У бактерий, как и у высших организмов, существуют механизмы, которые обеспечивают перекомбинацию генетического материала между родственными, но генетически неидентичными клетками. В результате такого обмена возникает потомство с новой комбинацией наследственных признаков – рекомбинанты. Однако способы обмена генетической информацией и процессы формирования рекомбинантов у бактерий отличаются от существующих у эукариот, что вносит своеобразие в рекомбинационный анализ генетической организации бактериальной хромосомы.
Известны три механизма передачи генетического материала у бактерий: конъюгация, трансформация и трансдукция.
Конъюгация – это направленный перенос генетического материала из клетки-донора в клетку-реципиент при непосредственном физическом контакте между ними.
Трансформация – способ передачи генетической информации от клеток-доноров к клеткам-реципиентам с помощью химически чистой ДНК.
Трансдукция – это перенос генетического материала из клеток-доноров в клетки-реципиенты с помощью бактериофагов.
Основная отличительная черта всех трех механизмов – это однонаправленный перенос от одной клетки (донора) к другой (реципиенту) части генетического материала (рис.5). Вследствие такого обмена возникает не полноценная диплоидная зигота, а частично диплоидная (меродиплоидная) зигота, или, так называемая, мерозигота. В мерозиготе генетический материал реципиентной клетки представлен целой хромосомой, а материал донорной клетки только фрагментом хромосомы или частью генома.
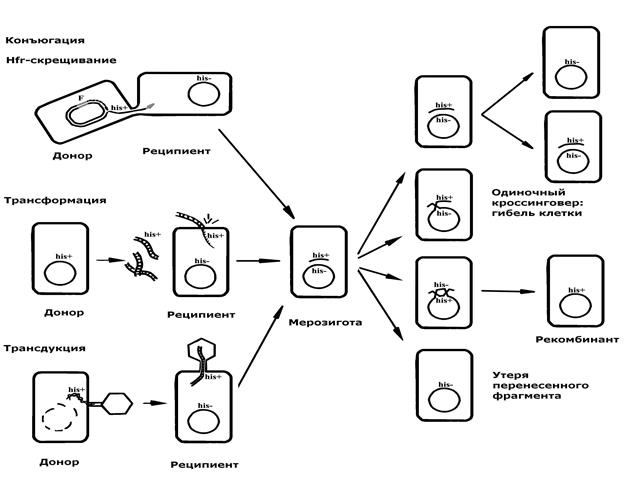 |
Рис. 5. Механизмы передачи генетического материала у бактерий и судьба перенесенного фрагмента
Судьба перенесенного фрагмента донорной ДНК в мерозиготе неоднозначна и зависит от многих факторов. Обычно он не способен к самостоятельной редупликации в клетке реципиента, так как не является репликоном, но некоторое время может в ней сохраняться и даже функционировать (транскрибироваться). В этом случае фрагмент ДНК донора при каждом делении мерозиготы будет передаваться лишь одной из дочерних клеток (однолинейная передача). Однако такое состояние не стабильно и может привести либо к утере фрагмента, либо к включению его в хромосому реципиентной клетки за счет рекомбинации (кроссинговера или конверсии) с ее гомологичной последовательностью.
Важно отметить ту особенность рекомбинации у бактерий, что только четное число обменов между хромосомой реципиента и переданным фрагментом донора дает полноценную хромосому. Единичный или нечетный кроссинговер приводит к размыканию хромосомы и дупликации концевых фрагментов (рис.5). Следовательно, элементарным регистрируемым актом рекомбинации в мерозиготе является двойной кроссинговер. Результатом двойного (или четного) обмена является реципрокная интеграция донорного фрагмента в реципиентную хромосому и участка реципиентной хромосомы в донорный фрагмент. Так как донорный фрагмент обычно не реплицируется и в дальнейшем теряется, регистрируется только интеграция в хромосому. Включенный фрагмент (или его участок) будет реплицироваться совместно с хромосомой реципиента и передаваться потомству. В результате в популяции реципиентных клеток появятся стабильные рекомбинанты, получившие некоторые признаки донора. Отмеченная особенность рекомбинации приводит к нереципрокности и уменьшенному вдвое выходу рекомбинантов.
Следует также напомнить, что бактериальные клетки в различных условиях могут содержать несколько копий геномных молекул ДНК. Поэтому рекомбинанты появляются не сразу, а после 3-4 последовательных делений, когда клетки становятся гаплоидными (несут одинаковые копии).
Размер перенесенного фрагмента хромосомы донора зависит от способа передачи генетического материала. Например, при трансдукции он ограничен объемом головки бактериофага, в которую может быть ошибочно упакована ДНК клетки-хозяина. При трансформации длина переносимой молекулы очень мала и зависит от способа приготовления препарата ДНК и специфики конкретной системы. Только при конъюгации размер переносимого генетического материала может значительно колебаться и в редких случаях достигать размера полного генома.
Дата добавления: 2015-02-07 ; просмотров: 2399 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник
Микра / Лекции / Скорина / Лекция 21
ТЕМА 21 «ХАРАКТЕРИСТИКА СПОСОБОВ ГЕНЕТИЧЕСКОГО ОБМЕНА У БАКТЕРИЙ»
1. Способы генетического обмена у бактерий
2. Бактериальная трансформация
3. Бактериальная конъюгация
4. Бактериальная трансдукция
1. Способы генетического обмена у бактерий

Рис. 1 — Схема мерозиготы
Трансформация – перенос генетической информации, при котором ДНК, выделенная из клетки-донора, поступает в клетку-реципиент.
Явление трансформации было открыто Ф. Гриффитом в 1928 г. в опытах на пневмококках (Streptococcus pneumoniae). В классической работе О. Эвери, К. Мак-Леода и М. МакКарти, опубликованной в 1944 году, было установлено, что трансформирующим началом является ДНК.
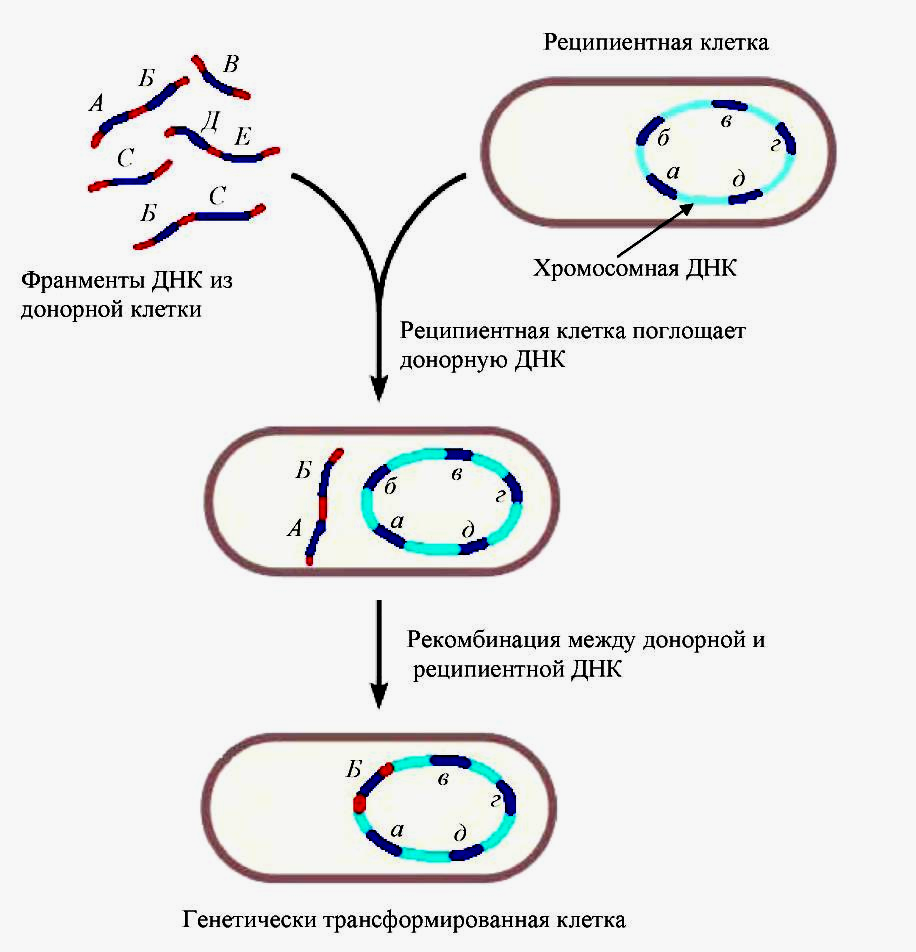
Рис. 2 — Схема процесса трансформации
Трансформация имеет практическое использование:
• для картирования бактериальной хромосомы;
• для конструирования промышленно-полезных штаммов микроорганизмов;
• для введения в геном бактерий определенных маркеров или элиминирования нежелательных мутаций;
• как один из этапов получения трансгенных растений;
• может выступать в качестве модели в различных генетических и молекулярно-биологических экспериментах на изолированной ДНК
Конъюгация – генетический обмен, который сопровождается переносом генетической информации от клетки-донора к клетке-реципиенту, он происходит при их непосредственном контакте.
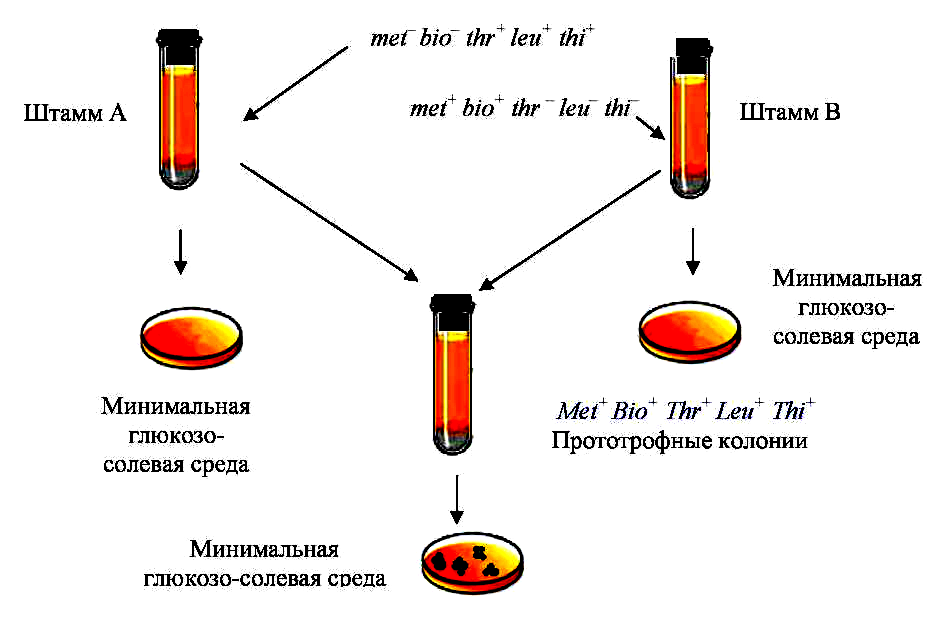
Рис. 3 — Схематическое изображение классического опыта по скрещиванию
ауксотрофных мутантов, проведенного Дж. Ледербергом и Э. Татумом
Явление конъюгации было открыто Дж. Ледербергом и Э. Татумом в 1946 г. в экспериментах с полиауксотрофными штаммами бактерий E. сoli (рис. 3). В 1949 г. Б. Дэвис получил дополнительные данные, которые также доказывали, что для образования прототрофов необходим контакт родительских клеток (рис.4).

Рис. 4 — Схема эксперимента Б. Дэвиса
Позднее У. Хейс показал, что существуют бактерии мужского и женского типа и вклад их в конъюгацию не равнозначен. Рекомбинанты наследуют большинство своих признаков от реципиента, а от донора получают только отдельные фрагменты генома.
Женская бактериальная клетка, обозначается как F — -штамм [бактерии]. (F- fertility). Это бактериальная клетка, которая не содержит F-фактора и участвует в конъюгации в качестве рецепиента; рекомбинация может происходить только в Ж.б.к.
Мужская бактериальная клетка, F + -штамм [бактерии]. Обозначается как бактериальная клетка, которая несет половой F-фактор и участвует в процессе конъюгации в качестве донора генетического материала; рекомбинация в М.б.к. никогда не происходит.
F-фактор представляет собой внехромосомную кольцевую двухцепочечную молекулу ДНК, которая автономно реплицируется, его относят к плазмидам. При конъюгации частота передачи F-фактора близка к 100%. Таким образом, клетки-реципиенты превращаются в потенциальных доноров.
В зависимости от состояния F-фактора различают два типа донорных клеток:
— F + -доноры, у которых F-фактор находится в автономном от хромосомы состоянии. При скрещивании обычно передается только F-фактор;
— Доноры Hfr— типа (high frequency of recombination (высокая частота рекомбинации), у которых F-фактор интегрирован в хромосому. При скрещивании передаются хромосомные гены. Интеграция F-фактора в бактериальную хромосому обратима.
F- факторы, которые содержат фрагменты хромосомной ДНК, получили название F́ — факторы (прим). Такие факторы могут нести в своем составе один ген – это малые F́-факторы, если несут до половины бактериальной хромосомы – это большие. F́- факторы с высокой эффективностью передаются при конъюгации клеткам – реципиентам, и переносят при этом бактериальные гены, которые включены в их состав. Такой тип передачи генов получил название сексдукции, или F- дукции.

Рис. 5 — Микрофотография конъюгирующих клеток E. coli
Конъюгация используется в следующих направлениях:
1. Передача многих генетических маркеров из одних клеток в другие. Показано, что при конъюгации вся хромосома бактерий E. coli передается за 100 мин.
2. Метод конъюгационного скрещивания удобен для картирования хромосомы. Он был первым методом, который использовался для этих целей. Карта хромосомы у бактерий строится в минутах (рис. 6).
3. Изучение генетического аппарата у бактерий.
4. Конъюгация эффективно происходит в природе и поэтому является одним из факторов изменчивости бактерий.

Рис. 6 — Генетическая карта E. coli
Трансдукция – перенос генетической информации (хромосомных генов или плазмид) от клетки-донора к клетке-реципиенту; происходит при участии бактериофагов.
Трансдукция была открыта Дж. Ледербергом и Н.Циндером в 1952 у Salmonella typhimurium и фага Р22.
Принято выделять два типа трансдукции: 1) общая (генерализованная, неспецифическая) (рис. 7); 2) специфическая (или ограниченная) (рис. 8).
При общей трансдукции может переноситься любой фрагмент бактериальный хромосомы с частотой 10 –5 –10 –6 . Количество бактериальной ДНК, которое может переноситься фагом, обычно составляет 1–2 % от всей клеточной ДНК. Исключение составляет бактериофаг РBS1 B. subtilis, который может трансдуцировать до 8 % генома хозяина. В процессе общей трансдукции бактериальный вирус является только «пассивным» переносчиком генетического материала бактерий и содержит только фрагменты бактериальной ДНК. Механизм рекомбинации у трансдуцируемых бактерий соответствует общепринятой схеме (гомологичной рекомбинации).
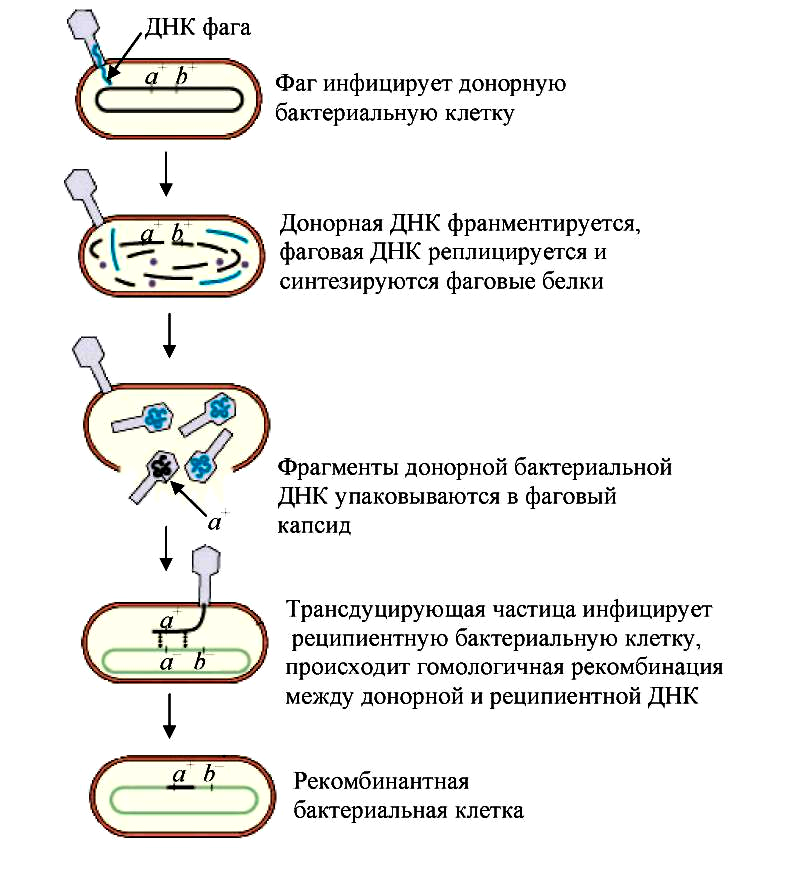
Рис. 7 — Схема генерализованной трансдукции
Специфическая трансдукция была открыта в 1956 г. М. Морзе и супругами Е. и Дж. Ледерберг.
Для специфической трансдукции характерно:
1) каждый трансдуцирующий фаг передает только определенную, строго ограниченную область бактериальной хромосомы;
2) фаг не только переносит генетический материал, но и обеспечивает его включение в бактериальную хромосому;
3) вирус включает фрагмент ДНК бактерий-доноров в свой геном и передает ее в ДНК бактерий-реципиентов в результате встраивания в хромосому.
Наиболее известным примером специфической трансдукции является трансдукция, которая выполняется фагом λ, который способен заражать клетки бактерий E. coli с последующей интеграцией его ДНК в геном бактерий.

Рис. 8 — Схема специфической трансдукции
Использовать трансдукцию можно в следующих направлениях:
• для трансдуцирования плазмид и коротких фрагментов хромосомы донора;
• для конструирования штаммов заданного генотипа;
• для точного картирования бактериальных генов, установления порядка их расположения в оперонах, что осуществляют с помощью комплементационного теста.
Источник
Горизонтальный обмен генами
Горизонтальный обмен генами
У одноклеточных организмов, понятное дело, нет разделения на соматические и половые клетки. Их единственная клетка является одновременно и половой, и соматической, и любые произошедшие в ней изменения генов беспрепятственно и неизбежно передаются потомкам. А гены у одноклеточных организмов изменяются довольно часто. И это не только мутации. У них очень широко распространен так называемый горизонтальный обмен генетическим материалом.
Три способа горизонтального обмена генами у бактерий:
конъюгация: две бактерии соединяются при помощи специальных белковых трубочек — конъюгационных пилей, и бактерия-донор передает бактерии-реципиенту часть своего генома;
вирусная трансдукция: вирусы, переходя из одной бактерии в другую, могут прихватывать с собой куски бактериального генома;
естественная трансформация: иногда бактерия просто «всасывает» фрагменты ДНК из окружающей среды и при определенных условиях встраивает их в свой геном. Как мы помним из главы «Великий симбиоз», этот способ межвидового генетического обмена мог сыграть важную роль в становлении эукариотической клетки.
Когда бактерия встраивает в свою единственную кольцевую хромосому кусочки чужого генома, она меняет свои свойства, то есть фактически превращается в другой организм. Новые свойства — «приобретенные признаки», — естественно, передаются потомству. В предельном случае возможна даже полная замена собственного генома бактерии чужим геномом. Если последний получен от другого вида бактерий, происходит нечто совершенно чудесное: бактериальная клетка в одночасье меняет свою видовую принадлежность. Микроб, относящийся к виду А, трансформируется в микроба вида Б. Самое удивительное, что это не чисто теоретические рассуждения, а экспериментально доказанный факт. Он был установлен в 2007 году исследователями из института Крейга Вентера (США).
Первая в мире операция по пересадке генома позволила превратить один вид бактерий в другой. Ученые из Института Крейга Вентера в течение последних 10 лет уверенно идут к великой цели — созданию искусственных микроорганизмов с заданными свойствами. Практическое значение этих работ может оказаться огромным. Например, планируется создание микробов, которые будут в больших количествах производить дешевое топливо. Генеральная идея состоит в том, чтобы установить минимальный набор генов, необходимый для жизнеобеспечения бактерии, добавить туда гены, кодирующие полезные функции, например, синтез водорода, искусственно синтезировать спроектированный геном и внедрить его в живую бактерию. Ее собственный геном при этом должен быть каким-то образом удален.
Работы ведутся в основном с бактериями рода Mycoplasma.
Микоплазмы — довольно обширная (около 180 видов) группа паразитических бактерий, вызывающих всевозможные болезни у растений, животных и человека. Микоплазмы обладают рядом уникальных свойств, которые делают их весьма удобным объектом для подобных исследований. Геномы микоплазм очень малы — от 600 до 1400 тыс. пар нуклеотидов — и хорошо изучены. На сегодняшний день полностью прочтены геномы 14 видов микоплазм. В отличие от подавляющего большинства других бактерий с маленькими геномами микоплазмы не являются облигатными внутриклеточными паразитами. Они могут жить вне хозяйских клеток, поэтому их можно выращивать обычным образом на питательной среде. Правда, среда должна быть весьма богатой: микоплазмы очень требовательны в этом отношении, поскольку у них отсутствуют гены, необходимые для синтеза многих жизненно важных веществ. Наконец, у микоплазм нет жесткой клеточной стенки, характерной для большинства бактерий. Клетки микоплазм окружены лишь тонкой и эластичной мембраной. Это сильно облегчает обмен наследственным материалом между клетками.
Изучая геномы микоплазм, Крейг Вентер и его коллеги уже очень близко подошли к пониманию того, что должен представлять собой «минимальный геном» будущих искусственных микробов. Синтез искусственных фрагментов генома уже налажен, синтез целого бактериального генома — дело недалекого будущего. Биологи давно научились внедрять в бактерий отдельные фрагменты геномов. В этом ученым большую помощь оказывают имеющиеся у микробов естественные механизмы для обмена генетическим материалом. Однако до сих пор никому не удавалось пересадить целый геном в живую бактериальную клетку.
В июне 2007 года Крейг Вентер и его сотрудники сообщили о первой успешной трансплантации целого генома от одного вида бактерий другому. Правда, ученые пока сами не до конца понимают, как им это удалось и пройдет ли этот номер с другими видами бактерий. Сделано было следующее. Ученые выделили геном из бактерии Mycoplasma mycoides, которая вызывает пневмонию у коров. Геном этого микроба, как и у большинства бактерий, представляет собой одну кольцевую молекулу ДНК. Геном был тщательно очищен от посторонних примесей, в том числе от белков, и добавлен в культуру бактерий Mycoplasma capricolum, возбудителей козьего полиартрита. Предварительно в геном M. mycoides были внесены особые метки, в том числе гены устойчивости к антибиотикам. По этим меткам можно потом определить, успешно ли прошла трансплантация.
Спустя недолгое время среди клеток Mycoplasma capricolum появились бактерии с признаками Mycoplasma mycoides. Обработав культуру бактерий антибиотиком, ученые уничтожили тех микробов, которые не вобрали в себя чужую ДНК, а оставшихся подвергли тщательному изучению. По всем признакам это были самые настоящие M. mycoides. Ни генов, ни белков, характерных для исходного вида Mycoplasma capricolum, у них обнаружить не удалось. Антитела, избирательно реагирующие на поверхностные белки Mycoplasma capricolum, не прикреплялись к этим микробам, в отличие от антител, распознающих поверхностные белки Mycoplasma mycoides.
Все это свидетельствует о том, что пересадка генома полностью удалась. Авторы предполагают, что бактерии «проглатывали» чужую молекулу ДНК, и в первый момент в них, вероятно, содержались оба генома вместе. Когда такая клетка делилась, одна из дочерних клеток получала геном Mycoplasma capricolum, а другая — геном Mycoplasma mycoides. Последующая обработка антибиотиком уничтожила клетки первого типа.
Дальнейшие исследования покажут, можно ли проделывать подобную манипуляцию с другими бактериями и другими геномами. Не исключено, что вобрать в себя целый чужой геном способны только микробы, не имеющие клеточной стенки, — в этом случае микоплазмы, скорее всего, и впредь останутся единственными объектами для таких экспериментов. Так или иначе, проделанная работа сильно приблизила Крейга Вентера к его заветной цели — созданию искусственного микроба. По-видимому, эта цель может быть достигнута уже через несколько лет. Кстати сказать, в США сейчас активно дискутируются этические и юридические проблемы, связанные с близящимся созданием искусственных организмов. Самые горячие споры идут по вопросу о том, можно ли будет эти организмы патентовать.
(Источник: Lartigue C. et al. Genome Transplantation in Bacteria: Changing One Species to Another // Science. 2007. V. 317. P. 632–638.)
У многоклеточных горизонтальный обмен генами между неродственными организмами играет гораздо меньшую роль. Вместо него развились более совершенные механизмы перемешивания и перекомбинирования наследственной информации, связанные с половым размножением. По сути дела это тот же самый горизонтальный обмен, но только замкнутый в пределах вида (разные особи смешивают свои гены в потомстве, но с представителями других видов обмен генами резко ограничен). К тому же половые железы у животных действительно ограждены от влияний внешней среды особым «вейсмановским» барьером, через который могут проникать только очень немногие вещества, в основном небольшие молекулы.
Тем не менее многоклеточные животные и растения время от времени заимствуют гены у микроорганизмов, например, у паразитических или симбиотических бактерий. Поскольку эти бактерии, в свою очередь, могут заимствовать гены у своих хозяев, а также переходить от одного хозяина к другому, они могут служить посредниками при переносе генетического материала между разными видами хозяев. По-видимому, это происходит чрезвычайно редко. Надежные экспериментальные подтверждения переноса генов от бактерий к многоклеточным были получены только недавно, и их пока очень мало. Но нужно иметь в виду, что редкость события вовсе не обязательно означает, что его роль в эволюции мала и незначительна. Ведь ключевые эволюционные преобразования сами по себе являются весьма редкими событиями — это, что называется, «штучный товар». Роль горизонтального переноса генов в эволюции многоклеточных еще предстоит оценить, и некоторые косвенные данные свидетельствуют о том, что она может быть весьма велика.
Рассмотрим один из случаев переноса генов бактерий в геном многоклеточного животного, обнаруженный в 2007 году. В данном случае «донором» генетического материала была паразитическая бактерия вольбахия, а «реципиентом» — мушка дрозофила. Этот случай интересен тем, что в геном насекомого встроились не отдельные гены, а целый бактериальный геном.
Вольбахия — паразитическая бактерия, обитающая в клетках многих наземных и пресноводных членистоногих и круглых червей — филярий. Вольбахию называют микробом-манипулятором, поскольку она научилась при помощи специальных регуляторных белков управлять размножением и развитием своих хозяев. Например, она умеет превращать самцов в самок, избирательно убивать зародышей мужского пола, повышать плодовитость зараженных самок и даже делать бесплодными самок, которые ею не заражены. О том, как ей это удается, можно прочесть в популярных статьях: А. В. Марков. Антимужской микроб. http://elementy.ru/lib/164668, А. В. Марков, И. А. Захаров-Гезехус. Бактерия вольбахия — повелитель мух. http://evolbiol.ru/wolbachia.htm. Вольбахия «впрыскивает» регуляторные белки в цитоплазму хозяина при помощи модифицированного конъюгационного аппарата, то есть поступает примерно так же, как ее дальняя родственница агробактерия — природный генный инженер, о котором мы говорили в заключительной части главы «Управляемые мутации». Вольбахия паразитирует в клетках беспозвоночных уже более 100 миллионов лет, да и ее предки — альфапротеобактерии из группы риккетсиевых — тоже были внутриклеточными паразитами. За это время вольбахия и ее хозяева успели приспособиться друг к другу. В ряде случаев вольбахия даже повышает жизнеспособность своих хозяев, то есть выступает в роли полезного симбионта. При таком долгом и тесном сожительстве было бы даже странно, если бы какие-то фрагменты генома вольбахии время от времени не попадали в ядра клеток хозяина и не включались в хозяйский геном. Однако доказать это удалось лишь в 2007 году.

Яйцо осы Trichogramma kaykai с множеством бактерий Wolbachia (черные точки). Вольбахии концентрируются в удлиненном кончике яйца, из которого впоследствии разовьются органы размножения осы. Бактерии попадут в репродуктивные органы, затем — в яйцеклетки, обеспечив себе гарантированный переход в следующее поколение насекомых-хозяев.
Довольно часто в ходе выполнения проектов по прочтению геномов высших организмов (особенно насекомых) исследователи натыкались на фрагменты бактериальных последовательностей ДНК, но это обычно интерпретировалось как результат загрязнения: предполагали, что при выделении ДНК из клеток исследуемого организма в пробы попало небольшое количество бактериальной ДНК. И соответствующие участки ДНК просто не учитывались при «сборке» генома из прочтенных фрагментов.
В середине 2007 года группа американских ученых предприняла широкомасштабный анализ таких «загрязнений» с целью найти реальные случаи переноса генов вольбахии в геномы животных-хозяев[93]. Ученые выделяли ДНК из разных видов насекомых и круглых червей — филярий, а также анализировали накопленные в Генбанке (http://www.ncbi.nlm.nih.gov/Genbank/) данные по нуклеотидным последовательностям различных беспозвоночных. Надо сказать, что многие прочтенные «вчерне» геномы до сих пор не подвергались процедуре окончательной сборки. Они хранятся в компьютерных базах в виде набора разрозненных, частично перекрывающихся обрывков разной длины. Если перенос генов от внутриклеточных бактерий к хозяевам действительно имеет место, среди этих обрывков могут обнаружиться такие куски ДНК, которые содержат одновременно и эукариотические, и бактериальные участки. Именно такие обрывки и интересовали исследователей.
В результате для четырех видов насекомых и четырех видов филярий удалось получить бесспорные доказательства внедрения генов вольбахии в геном хозяина; еще у трех видов это можно предполагать с большой долей вероятности.
Наибольшее внимание авторы уделили тропической плодовой мушке Drosophila ananassae, потому что в геноме некоторых представителей этого вида обнаружились полные или почти полные копии генома вольбахии. Получается, что в ядрах клеток этих мушек содержится полная генетическая информация сразу о двух разных организмах!
Для проверки этого результата ученые провели целый ряд специальных тестов. Мушек вылечили от вольбахии антибиотиком и убедились, что лечение привело к полному исчезновению внутриклеточных паразитов. Из вылеченных мух снова выделили ДНК. Оказалось, что полный набор генов вольбахии по-прежнему присутствует в пробах.
Затем проверили наследуемость этих генов по мужской линии. Дело в том, что вольбахия, как и другие цитоплазматические бактерии (вспомним митохондрии!), передается потомству только по материнской линии, вместе с цитоплазмой яйцеклетки. В сперматозоиды вольбахия не проникает — они для этого слишком малы. Поэтому потомство зараженной самки всегда оказывается зараженным, потомство здоровой — здоровым, а от отца это не зависит. Однако если геном вольбахии действительно встроился в геном хозяина, то он должен передаваться по отцовской линии точно так же, как и по материнской, — вместе с ядерными хромосомами.
Чтобы проверить это, скрестили вылеченных самцов D. ananassae, в хромосомы которых встроился геном вольбахии, со здоровыми самками, в геноме которых гены вольбахии отсутствовали. В ДНК потомства обнаружились гены вольбахии, что и стало решающим доказательством горизонтального переноса генов от паразита к хозяину. Авторы также показали, что многие гены, заимствованные мухой у бактерии, активно работают (то есть транскрибируются, «считываются»).
Полученные результаты показывают, что межвидовой обмен генами может играть более существенную роль в эволюции животных, чем считалось ранее. Источниками нового генетического материала для животных могут служить не только вирусы и мобильные генетические элементы, что было известно и ранее, но и бактерии. Впрочем, пока трудно сказать, насколько широк спектр бактерий, гены которых могут быть заимствованы животными.
Горизонтальный перенос генов привлекает в последние десятилетия пристальное внимание ученых — и отнюдь не только потому, что в некоторых случаях он может приводить к «ламарковскому» наследованию. Широкое распространение горизонтального переноса в живой природе заставляет пересмотреть еще одно основополагающее положение классического дарвинизма и СТЭ, а именно — точку зрения об исключительно дивергентном характере эволюции. «Дивергенция» означает «расхождение». Долгое время эволюционная теория базировалась на представлении о том, что виды не могут обмениваться друг с другом наследственной информацией. Как только вид делится на два, потомки теряют способность скрещиваться друг с другом, между ними возникает репродуктивная изоляция. Так что после разделения они эволюционируют изолированно, сами по себе, по схеме «случайные мутации + естественный отбор».
Дарвиновская схема дивергенции. Классический взгляд на эволюцию отражен в знаменитой дарвиновской «схеме дивергенции». Это единственный рисунок, которым Чарльз Дарвин сопроводил свой великий труд о происхождении видов. Эволюционный процесс на нем представлен в виде ветвящегося древа. Исходный вид делится на насколько ветвей — новых видов. Каждая ветвь может делиться дальше, и так до бесконечности. Отсутствие поперечных перемычек между ветвями показывает, что каждый вид эволюционирует сам по себе. Он должен самостоятельно изобретать все полезные адаптации, он не может «посоветоваться» с другими видами, перенять их опыт, заимствовать их «открытия».
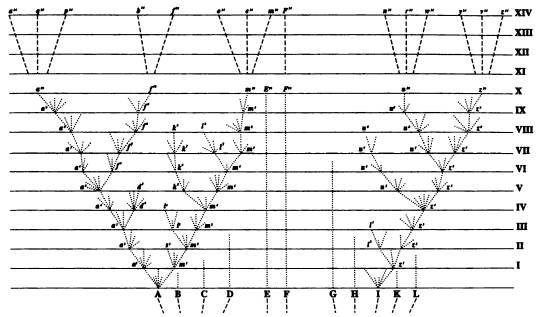
Классическая схема дивергенции по Дарвину имеет вид древа, ветви которого, раз разделившись, уже никогда более не сливаются.
Примерно так же выглядят практически все эволюционные реконструкции, публикуемые в научных статьях вплоть до настоящего времени.
Однако некоторые исследователи выражали сомнение в том, что при такой изолированной эволюции на основе мутаций и отбора жизнь успела бы за сравнительно недолгий срок своего существования (4 млрд лет) развиться от простейших форм до таких высокоорганизованных, как млекопитающие или насекомые.
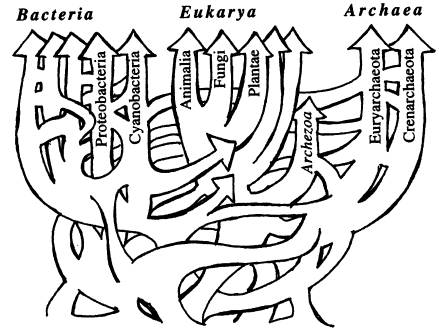
Схема эволюции жизни с учетом горизонтального переноса генов похожа не на дерево, а на запутанную сеть. Из статьи W. F. Doolittle. Phylogenetic Classification and the Universal Tree // Science. 1999. V. 284. P. 2124–2128.
С открытием горизонтального переноса генов между разными видами и даже царствами живых организмов ситуация изменилась, и эволюция предстает в несколько ином свете. Получается, что «удачные изобретения» одних видов в принципе доступны другим и могут быть ими заимствованы. В этом случае биосфера предстает единой информационной средой, в которой вирусы и различные мобильные генетические элементы (см. ниже) распространяют информацию примерно так же, как в человеческом обществе благодаря устной и письменной речи достижения и открытия одних людей становятся известными другим и могут ими использоваться.
Горизонтальный обмен генами вовсе не является бесконтрольным и неограниченным. Он не таков даже у прокариот, которые обмениваются генами относительно свободно и в каком-то смысле могут рассматриваться как единый, огромный и невероятно полиморфный вид. Уже у прокариот выработались различные механизмы выбора партнеров по обмену генами, и чаще всего такой обмен происходит все-таки между родственниками. Эукариоты выработали гораздо более эффективные и сложные адаптации для того, чтобы ограничивать и контролировать этот процесс. Важнейшими из этих адаптаций являются половое размножение и репродуктивная изоляция видов. Именно появление полового размножения и репродуктивной изоляции привело к формированию биологических систем нового типа — эндогамных видов. Эндогамия — избирательное скрещивание с себе подобными, со «своими», в отличие от экзогамии — скрещивания с «чужаками».
Однако межвидовая репродуктивная изоляция у эукариот все-таки не абсолютна. В частности, эукариоты тоже способны заимствовать чужие гены. Горизонтальный обмен фактически превращает биосферу в единую «лабораторию» по изобретению новых полезных наследственных признаков.
Правда, эффективность работы этой эволюционной лаборатории резко снижается по мере роста сложности организмов и их приспособленности. Большая часть адаптаций у сложных организмов зависит не от одного-двух, а от множества генов, которые должны работать согласованно и которые влияют не только на данный признак, но и на множество других. Поэтому вероятность того, что привнесенный извне чужой ген окажется полезным, у высших организмов значительно меньше, чем у низших. Именно этим, по-видимому, объясняется тот факт, что многоклеточные организмы выработали эффективные, хотя и не абсолютные, средства защиты от горизонтального переноса.
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРес
Читайте также
Перекресты. Расстояния между генами
Перекресты. Расстояния между генами Обычно дрозофилы сероватые, но есть среди них «блондины»—золотисто-желтые. Это рецессивный признак, вызванный геном из первой хромосомы. Его обозначают буквой «у», от английского слова yellow — желтый. Раз есть рецессив, значит, есть и
Обмен веществ
Обмен веществ Наши болезни все те же, что и тысячи лет назад, но врачи подыскали им более дорогие названия. Народная мудрость — Повышенный уровень холестерина может наследоваться — Ранняя смертность и гены ответственны за утилизацию холестерина — Наследуется ли
Микробам — горизонтальный перенос, высшим организмам — половое размножение
Микробам — горизонтальный перенос, высшим организмам — половое размножение Рассмотренные примеры говорят о пользе скрещивания и перемешивания генов при половом размножении. Но у бактерий и архей вместо настоящего амфимиксиса работает горизонтальный перенос. Будет ли
Горизонтальный перенос генов и симбиоз
Горизонтальный перенос генов и симбиоз Дупликация и перестановки участков ДНК одного и того же генома открывают большие эволюционные возможности. Еще больше их дает комбинирование фрагментов разных, неродственных геномов[76]. Обмен генами между неродственными
Обмен витаминов
Обмен витаминов Ни один из витаминов не осуществляет свои функции в обмене веществ в том виде, в котором он поступает с пищей. Этапы обмена витаминов:1. всасывание в кишечнике с участием специальных транспортных систем;2. транспорт к местам утилизации или депонирования с
Глава 18. Обмен гликогена
Глава 18. Обмен гликогена Гликоген – основной резервный полисахарид в животных тканях. Он представляет собой разветвленный гомополимер глюкозы, в котором остатки глюкозы соединены в линейных участках ?-1,4-гликозидными связями, а в точках ветвления – ?-1,6- гликозидными
Обмен жирных кислот
Обмен жирных кислот Высвобождающиеся при липолизе жирные кислоты поступают в кровоток и транспортируются в связанном с сывороточными альбуминами состоянии. Поступление СЖК сопровождается появлением в плазме также и глицерола. Глицерол может участвовать в
Обмен кетоновых тел
Обмен кетоновых тел При голодании, длительной физической нагрузке и в случаях, когда клетки не получают достаточного количества глюкозы (желудочно-кишечные расстройства у детей, диета с низким содержанием углеводов, почечная глюкозурия, сахарный диабет), в жировой ткани
Глава 26. Обмен нуклеотидов
Глава 26. Обмен нуклеотидов Практически все клетки организма способны к синтезу нуклеотидов (исключение составляют некоторые клетки крови). Другим источником этих молекул могут быть нуклеиновые кислоты собственных тканей и пищи, однако эти источники имеют лишь
Обмен железа
Обмен железа В организме взрослого человека содержится 3–4 г железа, из этого количества около 3,5 г находится в плазме крови. Гемоглобин эритроцитов содержит примерно 68 % всего железа организма, ферритин – 27 % (резервное железо печени, селезенки, костного мозга), миоглобин
2.3. Обмен веществ и энергии
2.3. Обмен веществ и энергии Вся совокупность химических реакций, протекающих в живых организмах, называется обменом веществ, или метаболизмом. В результате этих реакций энергия, запасенная в химических связях, переходит в другие формы, т. е. обмен веществ всегда
ОБМЕН УГЛЕВОДОВ
ОБМЕН УГЛЕВОДОВ Следует еще раз подчеркнуть, что процессы, происходящие в организме, представляют собой единое целое, и только для удобства изложения и облегчения восприятия рассматриваются в учебниках и руководствах в отдельных главах. Это относится и к разделению на
7.4. Углеродный обмен в биосфере
7.4. Углеродный обмен в биосфере На всем земном шаре, по оценкам ученых (Лархер, 1978, с. 128), растения ежегодно связывают около 155 109 т углерода. Из этого количества на долю суши приходится 61 %, а гидросферы – 39 % от общего его количества. Очень высокая первичная продуктивность на
7.6. Азотный обмен
7.6. Азотный обмен Азот, углерод, кислород и водород являются основообразующими химическими элементами, без которых (хотя бы в пределах нашей солнечной системы) не возникла бы жизнь. Азот в свободном состоянии обладает химической инертностью и является самым
16. Обмен веществ и превращение энергии. Энергетический обмен
16. Обмен веществ и превращение энергии. Энергетический обмен Вспомните!Что такое метаболизм?Из каких двух взаимосвязанных процессов он состоит?Где в организме человека происходит расщепление большей части органических веществ, поступающих с пищей?Обмен веществ и
13. Гений, управляющий генами
13. Гений, управляющий генами Откройте любой свежий журнал — и увидите, насколько популярна эпигенетика. Только в этом году в ПабМед[135] перечислено больше пяти тысяч статей по эпигенетике. Конечно, эта отрасль исследований имеет и свой собственный журнал, «Эпигенетика»,
Источник



