
- Энергетические процессы в мышце
- Содержание
- Энергетические процессы в мышце [ править | править код ]
- АТФ [ править | править код ]
- Креатинфосфат [ править | править код ]
- Энергетический метаболизм скелетных мышц [ править | править код ]
- Алактатные механизмы [ править | править код ]
- Гликолиз [ править | править код ]
- Окислительное фосфорилирование [ править | править код ]
- Восстановление и кислородная задолженность [ править | править код ]
- Энергообеспечение мышечной деятельности
- Общая характеристика аэробной системы энергообеспечения
- Лактатная система энергообеспечения.
- Алактатная система энергообеспечения.
- Общая характеристика систем энергообеспечения мышечной деятельности.
Энергетические процессы в мышце
Содержание
Энергетические процессы в мышце [ править | править код ]
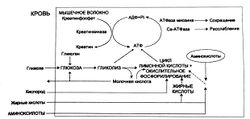
Естественно, что для совершения мышечного движения требуется энергия. В организме человека существуют разные источники энергии, которые последовательно включаются один за другим. Рассмотрим каждый из них.
АТФ [ править | править код ]
Универсальным источником энергии в живом организме является молекула АТФ, которая образуется в цитратном цикле Кребса. Под действием фермента АТФазы молекула АТФ гидролизуется, отсоединяя фосфатную группу в виде ортофосфорной кислоты (Н3РО4), и превращается в АДФ, при этом высвобождается энергия.
Головка миозинового мостика при контакте с актином обладает АТФазной активностью и соответственно возможностью расщеплять АТФ и получать энергию, необходимую для движения.
 Количества АТФ, которое содержится в мышце, достаточно для выполнения движений в течение 2-5 первых секунд.
Количества АТФ, которое содержится в мышце, достаточно для выполнения движений в течение 2-5 первых секунд.
Креатинфосфат [ править | править код ]
Запас молекул АТФ в мышце ограничен, поэтому расход энергии при работе мышцы требует постоянного его восполнения, это происходит за счет креатинфосфата. Креатинфосфат обладает способностью отсоединять фосфатную группу и превращаться в креатин, присоединяя фосфатную группу к АДФ, которая превращается в АТФ.
АДФ + креатинфосфат = АТФ + креатин.
Эта реакция получила название – реакции Ломана. Именно поэтому креатин имеет большое значение в бодибилдинге.
Надо заметить, что креатин эффективен только при выполнении анаэробных (силовых) упражнений, так как креатинфосфата достаточно примерно на 2 минуты интенсивной работы, затем подключаются другие источники энергии. Соответственно, в лёгкой атлетике приём креатина как добавки для увеличения атлетических показателей малоэффективен.
Запасы креатинфосфата в волокне не велики, поэтому он используется в качестве источника энергии только на начальном этапе работы мышцы, до момента активизации других более мощных источников – анаэробного и затем аэробного гликолиза. По окончании работы мышцы реакция Ломана идет в обратном направлении, и запасы креатинфосфата в течение нескольких минут восстанавливаются.
Энергетический метаболизм скелетных мышц [ править | править код ]
Алактатные механизмы [ править | править код ]
КФ обеспечивает запас энергии фосфата для ресинтеза АТФ из АДФ при наступлении сократительной деятельности (рис. 3):
КФ + АДФ Креатинкиназа К + АТФ (1)
В состоянии покоя мышечные волокна наращивают концентрацию КФ до пяти раз больше, чем АТФ. В начале сокращения, когда концентрация АТФ начинает падать, а АДФ повышаться вследствие ускорения разложения АТФ, массовая активность способствует образованию АТФ из КФ.
Хотя образование АТФ из КФ происходит быстро, требуя одной единственной ферментативной реакции (1), количество АТФ, которое может быть получено в результате этого процесса, ограничено начальной концентрацией КФ. Мышечные волокна также содержат миокиназу, которая катализирует образование одной молекулы АТФ и одной молекулы АМФ из двух молекул АДФ. АТФ и КФ, вместе взятые, могут обеспечить максимальную силу в течение 8—10 с. Таким образом, энергия, полученная от фосфагенной системы, используется для коротких всплесков максимальной мышечной активности, необходимых в легкой и тяжелой атлетике (забег на 100 м, толкание ядра или поднятие тяжестей).
Гликолиз [ править | править код ]
Хотя метаболизм по гликолитическому пути производит лишь небольшое количество АТФ из каждой усвоенной единицы глюкозы, он может обеспечить быстрый синтез большого количества АТФ при наличии достаточного количества ферментов и субстрата. Этот процесс может также происходить в отсутствие кислорода:
Глюкоза анаэробный быстрый гликолиз 2 АТФ + 2 лактата (2)
Глюкоза для гликолиза поступает либо из крови, либо из запасов гликогена. Когда исходным материалом выступает гликоген, из одной единицы потребленной глюкозы в результате фосфоролитического гликогенолиза образуется три молекулы АТФ. По мере того, как мышечная активность становится интенсивнее, для анаэробного расщепления гликогена мышц требуется все больше и больше АТФ, и, соответственно, увеличивается производство молочной кислоты. Анаэробный гликолиз может обеспечить энергию на 1,3-1,6 мин максимальной мышечной активности.
Образование молочной кислоты понижает уровень pH в мышечных волокнах. Это препятствует действию ферментов и вызывает боль, если удаление молочной кислоты происходит слишком медленно по сравнению с ее образованием.
Окислительное фосфорилирование [ править | править код ]
При умеренном уровне физической нагрузки, например, при беге на 5000 м или марафоне, большая часть АТФ, используемого для сокращения мышц, образуется путем окислительного фосфорилирования. Окислительное фосфорилирование позволяет высвободить из глюкозы гораздо больше энергии по сравнению с отдельно взятым анаэробным гликолизом:
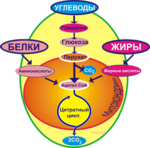
Жиры катаболизируются только с помощью окислительных механизмов, при этом выделяется много энергии. Аминокислоты тоже могут быть метаболизированы подобным образом. Три метаболических пути образования АТФ для сокращения и расслабления мышц показаны на рис. 3.
В течение первых 5
10 мин умеренной физической нагрузки главным потребляемым «топливом» является собственный гликоген мышц. В течение следующих 30 мин доминирующими становятся переносимые кровью вещества; глюкоза крови и жирные кислоты вносят примерно одинаковый вклад в потребление мышцами кислорода. По истечении этого периода все более важную роль приобретают жирные кислоты. Важно подчеркнуть взаимодействие между анаэробными и аэробными механизмами в образовании АТФ во время физической нагрузки. Вклад анаэробного образования АТФ больше при краткосрочной нагрузке высокой интенсивности, в то время как при более продолжительных нагрузках низкой интенсивности преобладает аэробный метаболизм.
Восстановление и кислородная задолженность [ править | править код ]
После того как физическая нагрузка закончилась, поглощение кислорода все еще остается выше нормы (табл.). С недавнего времени для обозначения кислородной задолженности используется также термин «избыточное потребление кислорода после физической нагрузки». Сначала его уровень очень высок, пока тело восстанавливает запасы КФ и АТФ, возвращая тканям запасенный кислород, а затем в течение еще одного часа потребление идет на более низком уровне, пока удаляется молочная кислота. Поэтому ранние и последние фазы кислородной задолженности называют соответственно алактатной и лактатной кислородной задолженностью. Повышение температуры тела также говорит о более высокой скорости метаболизма и росте потребления кислорода.
Чем продолжительнее и интенсивнее физическая нагрузка, тем больше времени занимает восстановление. Например, на восстановление после полного истощения гликогена мышц зачастую требуется несколько дней, а не секунд, минут или часов, необходимых для восстановления запасов КФ и АТФ и удаления молочной кислоты. Физическая нагрузка большой интенсивности, вероятно, приводит к микротравмам мышечных волокон, и их восстановление занимает некоторое время.
Компоненты кислородной задолженности. После длительной, тяжелой физической нагрузки дыхание остается выше нормы для удовлетворения повышенной потребности в кислороде
Восстановление запасов кислорода в тканях(около 1 л)
Восстановление уровней креатинфосфата и других богатых энергией фосфатов (около 1-1,5 л)
Удаление молочной кислоты путем глюконеогенеза и другими путями (до 12 л)
Стимуляция метаболизма вследствие повышения уровня адреналина (около 1 л)
Дополнительное потребление кислорода в дыхательных мышцах и сердце (около 0,5 л)
Общее усиление метаболизма вследствие более высокой температуры тела*
Q10 — повышение температуры на 10 °С может удвоить скорость метаболизма, если клетки могут справляться с такими изменениями температуры
Источник
Энергообеспечение мышечной деятельности
Общая характеристика аэробной системы энергообеспечения
Аэробная система энергообеспечения значительно уступает алактатной и лактатной по мощности энергопродукции, скорости включения в обеспечение мышечной деятельности, однако многократно превосходит по ёмкости и экономичности (табл. 1).
Таблица № 1. Энергообеспечение мышечной работы
| Источники | Пути образования | Время активации до максимального уровня | Срок действия | Продолжительность максимального выделения энергии |
|---|---|---|---|---|
| Лактатные анаэробные | АТФ, креатинфосфат | 0 c | До 30 с | До 10 с |
| Лактатные анаэробные | Гликолиз с образованием лактата | 15 – 20 с | От 15 – 20 с до 6 мин | От 30 с до 1 мин 30 с |
| Аэробные | Окисление углеводов и жиров кислородом воздуха | 90 – 180 с | До нескольких часов | 2 – 5 мин и более |
Аденозинтрифосфат или Аденозинтрифосфорная кислота — нуклеозид трифосфат, имеющий большое значение в обмене энергии и веществ в организмах. АТФ — универсальный источник энергии для всех биохимических процессов, протекающих в живых системах, в частности для образования ферментов.
Особенностью аэробной системы является то, что образование АТФ в клеточных органелах-митохондриях, находящихся в мышечной ткани происходит при участии кислорода, доставляемого кислородтранспортной системой. Это предопределяет высокую экономичность аэробной системы, а достаточно большие запасы гликогена в мышечной ткани и печени, а также практически неограниченные запасы липидов – её ёмкость.
В наиболее упрощенном виде деятельность аэробной системы энергообеспечения осуществляется следующим образом: на первом этапе в результате сложных процессов происходит преобразование как гликогена, так и свободных жирных кислот (СЖК) в ацетил-кофермент А (ацетил-КоА) – активную форму уксусной кислоты, что обеспечивает протекание всех последующих процессов энергообразования по единой схеме. Однако до момента образования ацетил-КоА окисление гликогена и СЖК происходит самостоятельно.
Все многочисленные химические реакции, происходящие в процессе аэробного ресинтеза АТФ, можно разделить на три типа:
система транспорта электронов (рис. 7).

Рис. 7. Этапы реакций ресинтеза АТФ в аэробном процессе
Первым этапом реакций является аэробный гликолиз, в результате которого осуществляется расщепление гликогена с образованием СО2 и Н2О. Протекание аэробного гликолиза происходит по той же схеме, что и протекание рассмотренного выше анаэробного гликолиза.
В обоих случаях в результате химических реакций гликоген преобразуется в глюкозу, а глюкоза – в пировиноградную кислоту с ре-синтезом АТФ. В этих реакциях кислород не участвует. Присутствие кислорода обнаруживается в дальнейшем, когда при его участии пировиноградная кислота не превращается в молочную кислоту, а затем в лактат, что имеет место в процессе анаэробного гликолиза, а направляется в аэробную систему, конечными продуктами которой оказывается углекислый газ (СО2), выводимый из организма легкими, и вода (рис. 8)

Рис. 8. Схематическое протекание анаэробного и аэробного гликолиза
Расщепление 1 моля гликогена на 2 моля пировиноградной кислоты происходит с выделением энергии, достаточной для ресинтеза 3 молей АТФ: Энергия + 3АДФ + Фн → 3АТФ.
Из образовавшейся в результате расщепления гликогена пировиноградной кислоты сразу выводится СО2, превращая её из трёхуглеродного соединения в двухуглеродное, которое сочетаясь с коферментом А, образует ацетил- КоА, который включается во второй этап аэробного образования АТФ – цикл лимонной кислоты или цикл Кребса.
В цикле Кребса протекает серия сложных химических реакций, в результате которых происходит окисление пировиноградной кислоты – выведение ионов водорода (Н+) и электронов (е-), которые в итоге попадают в систему транспорта кислорода и участвуют в реакциях ресинтеза АТФ на третьем этапе, образуя СО2, который диффундируется в кровь и переносится в легкие, из которых и выводится из организма. В самом цикле Кребса образуется только 2 моля АТФ (рис. 9).

Рис. 9. Схематическое изображение окисления углеродов в цикле Кребса
Третий этап протекает в цепи транспорта электронов (дыхательной цепи). Реакции, происходящие с участием коферментов, в общем виде сводятся к следующему. Ионы водорода и электроны, выделяемые в результате реакций, протекавших в цикле Кребса и в меньшей мере в процессе гликолиза, транспортируются к кислороду, чтобы в результате образовать воду. Одновременно выделяемая энергия в серии сопряженных реакций используется для ресинтеза АТФ. Весь процесс, происходящий по цепи передачи электронов кислороду называется окислительным фосфорилированием. В процессах, происходящих в дыхательной цепи, потребляется около 90 % поступающего к клеткам кислорода и образуется наибольшее количество АТФ. В общей сложности окислительная система транспорта электронов обеспечивает образование 34 молекул АТФ из одной молекулы гликогена.
Усвоение и абсорбция углеводов в кровоток происходит в тонком кишечнике. В печени они превращаются в глюкозу, которая в свою очередь может быть превращена в гликоген и депонируется в мышцах и печени, а также используется различными органами и тканями в качестве источника энергии для поддержания деятельности. В организме здорового с достаточным уровнем физической подготовленности мужчины с массой тела 75 кг содержится 500 – 550 г углеводов в виде гликогена мышц (около 80 %), гликогена печени (примерно 16 – 17 %), глюкозы крови (3 – 4 %), что соответствует энергетическим запасам порядка 2000 – 2200 ккал.
Гликоген печени (90 – 100 г) используется для поддержания уровня глюкозы в крови, необходимого для обеспечения нормальной жизнедеятельности различных тканей и органов. При продолжительной работе аэробного характера, приводящей к истощению запасов мышечного гликогена, часть гликогена печени может использоваться мышцами.
Следует учитывать, что гликогеновые запасы мышц и печени могут существенно увеличиваться под влиянием тренировки и пищевых манипуляций, предусматривающих углеводное истощение и последующее углеводное насыщение. Под влиянием тренировки и специального питания концентрация гликогена в печени может увеличиться в 2 раза. Увеличение количества гликогена повышает его доступность и скорость утилизации при выполнении последующей мышечной работы.
При продолжительных физических нагрузках средней интенсивности образование глюкозы в печени возрастает в 2 – 3 раза по сравнению с образованием её в состоянии покоя. Напряженная продолжительная работа может привести к 7 – 10-кратному увеличению образования глюкозы в печени по сравнению с данными, полученными в состоянии покоя.
Эффективность процесса энергообеспечения за счет жировых запасов определяется скоростью протекания липолиза и скоростью кровотока в адипозной ткани, что обеспечивает интенсивную доставку свободных жирных кислот (СЖК) к мышечным клеткам.
Если работа выполняется с интенсивностью 50 – 60 % VO2max, отмечается максимальный кровоток в адипозной ткани, что способствует максимальному поступлению в кровь СЖК. Более интенсивная мышечная работа связана с интенсификацией мышечного кровотока при одновременном уменьшении кровоснабжения адипозной ткани и, следовательно, с ухудшением доставки СЖК в мышечную ткань.
Хотя в процессе мышечной деятельности липолиз разворачивается, однако уже на 30 – 40-й минутах работы средней интенсивности её энергообеспечения в равной мере осуществляется за счет окисления как углеводов, так и липидов. Дальнейшее продолжение работы, приводящее к постепенному исчерпанию ограниченных углеводных ресурсов, связано с увеличением окисления СЖК. Например, энергообеспечение второй половины марафонской дистанции в беге или шоссейных велогонках (более 100 км) преимущественно связано с использованием жиров.
Несмотря на то что использование энергии от окисления липидов имеет реальное значение для обеспечения выносливости только при продолжительной мышечной деятельности, начиная уже с первых минут работы с интенсивностью, превышающей 60 % VO2max, отмечается освобождение из триацилглицеридов СЖК, их поступление и окисление в сокращающихся мышцах. Через 30 – 40 мин после начала работы скорость потребления СЖК возрастает в 3 раза, а после 3 – 4 часов работы – в 5 – 6 раз.
Внутримышечная утилизация триглицеридов существенно возрастает под влиянием тренировки аэробной направленности. Эта адаптационная реакция проявляется как в быстроте развертывания процесса образования энергии за счет окисления СЖК, поступивших из трицеридов мышц, так и в возрастании их утилизации из мышечной ткани.
Не менее важным адаптационным эффектом тренированной мышечной ткани является повышение ее способности к утилизации жировых запасов. Так, после 12-недельной тренировки аэробной направленности способность к утилизации триглицеридов в работающих мышцах резко возрастала и достигала 40 %.
Роль белков для ре-синтеза АТФ не существенна. Однако углеродный каркас многих аминокислот может быть использован в качестве энергетического топлива в процессе окислительного метаболизма, что проявляется при продолжительных нагрузках средней интенсивности, при которых вклад белкового метаболизма в энергопродукцию может достичь 5 – 6 % общей потребности в энергии.
Благодаря значительным запасам глюкозы и жиров в организме и неограниченной возможности потребления кислорода из атмосферного воздуха, аэробные процессы, обладая меньшей мощностью по сравнению с анаэробными, могут обеспечивать выполнение работы в течении длительного времени (т. е. их ёмкость очень велика при очень высокой экономичности).
Исследования показывают, что, например в марафонском беге за счет использования мышечного гликогена работа мышц продолжается в течении 80 мин. Определённое количество энергии может быть мобилизовано за счет гликогена печени. В сумме это может обеспечить 75 % времени, необходимого для преодоления марафонской дистанции. Остальная энергия образуется в результате окисления жирных кислот. Однако скорость их диффузии из крови в мышцы ограничена, что лимитирует производство энергии за счёт этих кислот. Энергии, продуцируемой вследствие окисления СЖК, достаточно для поддержания интенсивности работы мышц на уровне 40 – 50 % VO2max. В то время, как сильнейшие марафонцы способны преодолевать дистанцию с интенсивностью, превышающей 80 – 90 % VO2max, что свидетельствует о высоком уровне адаптации аэробной системы энергообеспечения, позволяющем не только обеспечить оптимальное сочетание использования углеводов, жиров, отдельных аминокислот и метаболитов для производства энергии, но и экономное расходование гликогена.
Таким образом, вся совокупность реакций, обеспечивающих аэробное окисление гликогена, выглядит следующим образом:
На первом этапе в результате аэробного гликолиза образуется пировиноградная кислота и ре-синтезируется некоторое количество АТФ.
На втором, в цикле Кребса, производится СО2, а ионы водорода (Н+) и электроны (е-) вводятся в систему транспорта электронов также с ре-синтезом некоторого количества АТФ.
И наконец, заключительный этап связан с образованием Н2О из Н+, е- и кислорода с высвобождением энергии, используемой для ре-синтеза подавляющего количества АТФ.
Жиры и белки, используемые в топлива для ре-синтеза АТФ, также проходят через цикл Кребса и систему транспорта электронов (рис. 10).

Рис. 10. Схематическое изображение функционирования аэробной системы энергообеспечения
Лактатная система энергообеспечения.
В лактатной системе энергообеспечения ресинтез АТФ происходит за счёт расщепления глюкозы и гликогена при отсутствии кислорода. Этот процесс принято обозначать как анаэробный гликолиз. Анаэробный гликолиз является значительно более сложным химическим процессом по сравнению с механизмами расщепления фосфогенов в алактатной системе энергообеспечения. Он предусматривает протекание серии сложных последовательных реакций, в результате которых глюкоза и гликоген расщепляются до молочной кислоты, которая в серии сопряжённых реакций используется для ресинтеза АТФ (рис. 2).

Рис. 2. Схематическое изображение процесса анаэробного гликолиз.
В результате расщепления 1 моля глюкозы образуется 2 моля АТФ, а при расщеплении 1 моля гликогена – 3 моля АТФ. Одновременно с высвобождением энергии в мышцах и жидкостях организма происходит образование пировиноградной кислоты, которая затем преобразуется в молочную кислоту. Молочная кислота быстро разлагается с образованием её соли – лактата.
Накопление молочной кислоты в результате интенсивной деятельности гликолитического механизма приводит к большому образованию лактата и ионов водорода (Н+) в мышцах. В результате, несмотря на действие буферных систем, постепенно снижается мышечный pH с 7,1 до 6,9 и даже до 6,5 – 6,4. Внутриклеточный pH, начиная с уровня 6,9 – 6,8 замедляет интенсивность гликолитической реакции восстановления запасов АТФ, а при pH 6,5 – 6,4 расщепление гликогена прекращается. Таким образом, именно повышение концентрации молочной кислоты в мышцах ограничивает расщепление гликогена в анаэробном гликолизе.
В отличие от алактатной системы энергообеспечения, мощность которой достигает максимальных показателей уже на первой секунде работы, процесс активизации гликолиза разворачивается значительно медленнее и достигает высоких величин энергопродукции только на 5 – 10 секундах работы. Мощность гликолитического процесса значительно уступает мощности креатинфосфокиназного механизма, однако является в несколько раз более высокой по сравнению с возможностями системы аэробного окисления. В частности, если уровень энергопродукции АТФ за счёт распада КФ составляет 9 – 10 ммоль/кг с.м.т./с (сырая масса ткани), то при подключении гликолиза объем производимой АТФ может увеличиться до 14 ммоль/кг с.м.т./с. За счёт использования обоих источников ресинтеза АТФ в течении 3-минутной интенсивной работы мышечная система человека способна вырабатывать около 370 ммоль/кг с.м.т. При этом на долю гликолиза приходится не менее 80 % общей продукции. Максимальная мощность лактатной анаэробной системы проявляется на 20 – 25-й секундах работы, а на 30 – 60-й секундах гликолитический путь ресинтеза АТФ является основным в энергообеспечении работы.
Ёмкость лактатной анаэробной системы обеспечивает её превалирующее участие в энергопродукции при выполнении работы продолжительность до 30 – 90 с. При более продолжительной работе роль гликолиза постепенно снижается, однако остаётся существенной и при более продолжительной работе – до 5 – 6 мин. Общее количество энергии, которое образуется за счёт гликолиза, наглядно может быть оценено и по показателям лактата крови после выполнения работы, требующей предельной мобилизации лактатной системы энергообеспечения. У нетренированных людей предельная концентрация лактата в крови составляет 11 – 12 ммоль/л. Под влиянием тренировки ёмкость лактатной системы резко возрастает и концентрация лактата в крови может достигать 25 – 30 ммоль/л и выше.
Максимальные величины энергообразования и лактата в крови у женщин на 30 – 40 % ниже по сравнению с мужчинами такой же спортивной специализации. Юные спортсмены по сравнению со взрослыми отличаются невысокими анаэробными возможностями. максимальная концентрация лактата в крови при предельных нагрузках анаэробного характера у них не превышает 10 ммоль/кг, что в 2 – 3 раза ниже, чем у взрослых спортсменов.
Таким образом, адаптационные реакции лактатной анаэробной системы могут протекать в различных направлениях. Одним из них является увеличение подвижности гликолитического процесса, что проявляется в значительно более быстром достижении его максимальной производительности (с 15 – 20 до 5 – 8 с). Вторая реакция связана с повышением мощности анаэробной гликолитической системы, что позволяет ей продуцировать значительно большее количество энергии в единицу времени. Третья реакция сводится к повышению ёмкости системы и, естественно общего объема продуцируемой энергии, вследствие чего увеличивается продолжительность работы, преимущественно обеспечиваемая за счет гликолиза.
Максимальное значение лактата и pH в артериальной крови в процессе соревнований по некоторым видам спорта представлены на рис. 3.

Рис.3. Максимальные значения лактата и pH в артериальной крови у спортсменов, специализирующихся в различных видах спорта: а – бег (400, 800 м); б – скоростной бег на коньках (500, 1000м); в – гребля (2000 м); г – плавание 100 м; д – бобслей; е – велогонки (100 км)
(Eindemann, Keul, 1977)
Они дают достаточно полное представление о роли лактатных анаэробных источников энергии для достижения высоких спортивных результатов разных видах спорта и об адаптационных резервах системы анаэробного гликолиза.
При выборе оптимальной продолжительности работы, обеспечивающей максимальную концентрацию лактата в мышцах, следует учитывать, что максимальное содержание лактата отмечается при использовании предельных нагрузок, продолжительность которых колеблется в пределах 1 – 6 мин. Увеличение продолжительности работы связано с уменьшением концентрации лактата в мышцах.
Для выбора оптимальной методики повышения анаэробных возможностей важно проследить особенности накопления лактата при прерывистой работе максимальной интенсивности. Например, одноминутные предельные нагрузки с четырёх минутными паузами приводят к постоянному увеличению лактата в крови (рис. 4) при одновременном снижении показателей кислотно-основного состояния (рис. 5).

Рис. 4. Изменение концентрации лактата в крови в процессе прерывистой максимальной нагрузки (одноминутные упражнения с интенсивностью 95 %, разделенные периодами отдыха длительностью 4 мин) (Hermansen, Stenswold, 1972)

Рис. 5. Изменение pH крови при прерывистом выполнении одноминутных нагрузок максимальной интенсивности (Hollman, Hettinger, 1980)
Аналогичный эффект отмечается и при выполнении 15 – 20-секундных упражнений максимальной мощности с паузами около 3 минут (рис. 6).

Рис. 6. Динамика биохимических изменений у спортсменов при повторном выполнении кратковременных упражнений максимальной мощности (Н. Волков и др., 2000)
Алактатная система энергообеспечения.
Эта система энергообеспечения является наименее сложной, отличается высокой мощностью освобождения энергии и кратковременностью действия. Образование энергии в этой системе происходит за счёт расщепления богатых энергией фосфатных соединений – аденозинтрифосфата (АТФ) и креатинфосфата (КФ). Энергия, образующаяся в результате распада АТФ, в полной мере включается в процесс энергообеспечения работы уже на первой секунде. Однако уже на второй секунде выполнение работы осуществляется за счёт креатинфосфата (КФ), депонированного в мышечных волокнах и содержащего богатые энергией фосфатные соединения. Расщепление этих соединений приводит к интенсивному высвобождению энергии. Конечными продуктами расщепления КФ являются креатин (Кр) и неорганический фосфат (Фн). Реакция стимулируется ферментом креатинкиназа и схематически выглядит следующим образом:

Энергия, высвобождаемая при распаде КФ, является доступной для процесса ресинтеза АТФ, поэтому за быстрым расщеплением АТФ в процессе мышечного сокращения незамедлительно следует его ресинтез из АДФ и Фн с привлечением энергии, высвобождаемой при расщеплении КФ:

Ещё одним механизмом алактатной системы энергообеспечения является так называемая миокиназная реакция, которая активизируется при значительном мышечном утомлении, когда скорость расщепления АТФ существенно превышает скорость её ресинтеза. Миокиназная реакция стимулируется ферментом миокиназа и заключается в переносе фосфатной группы с одной молекулы на другую и образованием АТФ и аденозинмонофосфата (АМФ):

Аденозинмонофосфат (АМФ), являющийся побочным продуктом миокиназной реакции, содержит последнюю фосфатную группу и в отличие от АТФ и АДФ не может быть использован в качестве источника энергии. Миокиназная реакция активизируется в условиях, когда в силу утомления другие пути ресинтеза АТФ исчерпали свои возможности.
Запасы КФ не могут быть восполнены в процессе выполнения работы. Для его ресинтеза может быть использована только энергия, высвобождаемая в результате распада АТФ, что оказывается возможным лишь в восстановительном периоде после окончания работы.
Алактатная система, отличаясь очень высокой скорость освобождения энергии, одновременно характеризуется крайне ограниченной ёмкостью. Уровень максимальной алактатной анаэробной мощности зависит от количества фосфатов (АТФ и КФ) в мышцах и скорости их использования. Под влиянием тренировки спринтерского характера показатели алактатной анаэробной мощности могут быть значительно повышены. Под влиянием специальной тренировки мощность алактатной анаэробной системы может быть увеличена на 40 -80 %. Например, спринтерская тренировка в течении 8 недель бегунов привела к увеличению содержания АТФ и КФ в скелетной мышце в состоянии покоя примерно на 10 %.
Под влиянием тренировки в мышцах не только увеличивается количество АТФ и Кф, но и существенно возрастает способность мышечной ткани к их расщеплению. Ещё одной адаптационной реакцией, определяющей мощность алактатной анаэробной системы, является ускорение ресинтеза фосфатов за счет повышения активности ферментов, в частности креатинфосфокиназы и миокиназы.
Под влиянием тренировки существенно возрастают и показатели максимальной ёмкости алактатной анаэробной системы энергообеспечения. Емкость алактатной анаэробной системы под влиянием целенаправленной многолетней тренировки может возрастать в 2,5 раза. Это подтверждается показателями максимального алактатного О2-долга: у начинающих спортсменов он составляет 21,5 мл/кг, у спортсменов высокого класса может достигать 54,5 мл/кг.
Увеличение ёмкости алактатной энергетической системы проявляется и в продолжительности работы максимальной интенсивности. Так, у лиц не занимающихся спортом, максимальная мощность алактатного анаэробного процесса, достигнутая через 0,5 – 0,7 с после начала работы, может удерживаться не более 7 – 10 с, то у спортсменов высшего класса, специализирующихся в спринтерских дисциплинах, она может проявляться в течение 15 – 20 с. При этом большая продолжительность работы сопровождается и значительно большей её мощностью, что обусловливается высокой скоростью распада и ресинтеза высокоэнергетических фосфатов.
Концентрация АТФ и КФ у мужчин и женщин практически одинакова – около 4 ммоль/кг АТФ и 16 ммоль/кг КФ. Однако общее количество фосфогенов, которые могут использоваться при мышечной деятельности, у мужчин значительно больше, чем у женщин, что обусловлено большими различиями в общем объеме скелетной мускулатуры. Естественно, что у мужчин значительно больше ёмкость алактатной анаэробной системы энергообеспечения.
В заключении следует отметить, что лица с высоким уровнем алактатной анаэробной производительности, как правило, имеют низкие аэробные возможности, выносливость к длительной работе. Одновременно у бегунов на длинные дистанции алактатные анаэробные возможности не только не сравнимы с возможностями спринтеров, но и часто уступают показателям, регистрируемым у лиц, не занимающихся спортом.
Общая характеристика систем энергообеспечения мышечной деятельности.
Энергия, как известно, представляет собой общую количественную меру, связывающую воедино все явления природы, разные формы движения материи. Из всех видов энергии, образующейся и использующейся в различных физических процессах(тепловая, механическая, химическая и др.)применительно к мышечной деятельности, основное внимание должно быть сконцентрировано на химической энергии организма, источником которой являются пищевые продукты и её преобразовании в механическую энергию двигательной деятельности человека.
Энергия, высвобождаемая во время расщепления пищевых продуктов, используется для производства аденозинтрифосфата (АТФ), который депонируется в мышечных клетках и является своеобразным топливом для производства механической энергии мышечного сокращения.
Энергию для мышечного сокращения даёт расщепление аденозинтрифосфата (АТФ) до аденозиндифосфата (АДФ) и неорганического фосфата (Ф). Количество АТФ в мышцах невелико и его достаточно для обеспечения высокоинтенсивной работы лишь в течении 1 – 2 с. Для продолжения работы необходим ресинтез АТФ, который производится за счет энерго отдающих реакций трёх типов. Восполнение запасов АТФ в мышцах позволяет поддерживать постоянный уровень его концентрации, необходимый для полноценного мышечного сокращения.
Ресинтез АТФ обеспечивается как в анаэробных, так и в аэробных реакциях с привлечением в качестве энергетических источников запасов креатинфосфата (КФ) и АДФ, содержащихся в мышечных тканях, а также богатых энергией субстратов (гликоген мышц и печени, запасы липозной ткани и др.). Химические реакции, приводящие к обеспечению мышц энергией протекают в трёх энергетических системах:
- анаэробной алактатной
- анаэробной лактатной (гликолитической)
- аэробной.
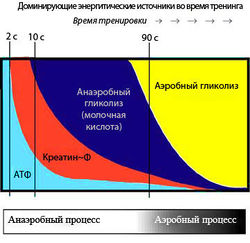
Образование энергии в первых двух системах осуществляется в процессе химических реакций, не требующих наличия кислорода. Третья система предусматривает энергообеспечение мышечной деятельности в результате реакций окисления, протекающих с участием кислорода. Наиболее общие представления о последовательности включения и количественных соотношениях в энергообеспечении мышечной деятельности каждой из указанных систем приведены на рис. 1.

Рис. 1. Последовательность и количественные соотношения процессов энергообеспечения мышечной деятельности у квалифицированных спортсменов в различных энергетических системах (схема): 1 – алактатной; 2 – лактатной; 3 – аэробной.
Возможности каждой из указанных энергетических систем определяются мощностью, т. е. скоростью освобождения энергии в метаболических процессах, и ёмкостью, которая определяется величиной и эффективностью использования субстратных фондов.
Источник



