
Подцарство простейшие
Простейшие — одноклеточные организмы. Безусловно, ни о каких тканях, органах не может идти и речи — но это совершенно не означает, что у простейших не идут процессы газообмена, выделения, транспорта питательных веществ — все они идут, но по-особенному.
У простейших одна клетка выполняет все функции целого организма, поэтому клетки имеют сложное строение. Клетки обладают всеми основными жизненными функциями: раздражимостью, размножением, обменом веществ.
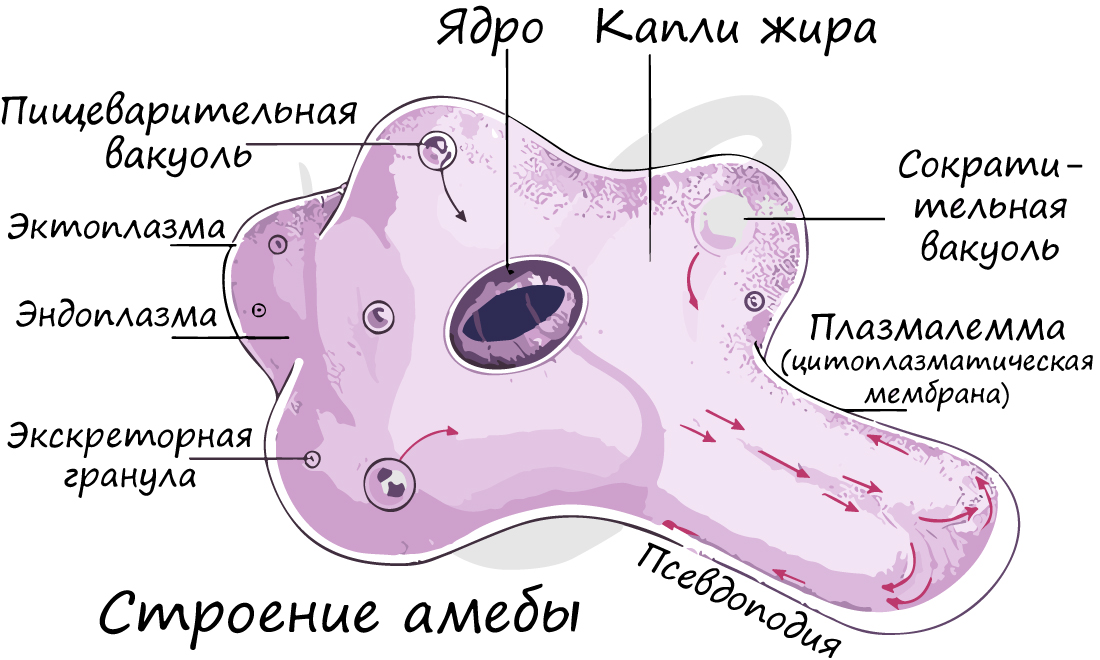
Строение клетки простейшего
Форма клетки простейших постоянная, окружена пелликулой — наружным, уплотненным слоем цитоплазмы, который поддерживает постоянную форму. У некоторых простейших (амеба, на рисунке выше) пелликула отсутствует и форма клетки непостоянная, растекающаяся.
Клетка простейших является эукариотической — имеет оформленное ядро, обособленное ядерной мембраной от цитоплазмы. В цитоплазме многих простейших выделяют эктоплазму (периферический наружный, более плотный слой цитоплазмы) и эндоплазму (внутренний зернистый слой цитоплазмы, менее плотный, подвижен).
Типичным для эукариотов является набор органоидов в клетке: митохондрии, эндоплазматический ретикулум (сеть), аппарат (комплекс) Гольджи, запасные питательные вещества (гликоген, жировые включения), рибосомы, лизосомы.
Сократительные вакуоли
Особенностью строения, является наличие в клетке простейших сократительных вакуолей, которые служат для поддержания осмотического давления. В клетку простейших постоянно поступает избыток воды, и, чтобы клетку не разорвало от повышенного давления, вода постоянно удаляется из клетки. Таким образом, функцию выделения выполняют сократительные вакуоли.

Работа сократительной вакуоли подчинена определенному механизму. Сначала лучистые канальцы, расположенные вокруг вакуоли, накапливают воду. При скоплении в них достаточно большого количества воды они изливают ее в центральную полость — сократительную вакуоль. Вакуоль сокращается и избыток воды удаляется из клетки во внешнюю среду, таким образом, разрыв клетки предотвращается.
Хемотаксис
Поскольку нервная система отсутствует, раздражимость у простейших осуществляется с помощью хемотаксиса. Хемотаксис — движение подвижных организмов под влиянием одностороннего раздражения химическими веществами. Хемотаксис может быть положительным (движение по направлению к химическому веществу) или отрицательным (движение в обратном направлении, от химического вещества).
Пищеварительная система также отсутствует, ее функция передана пищеварительным вакуолям. Тип питания — внутриклеточный, осуществляется с помощью фагоцитоза (от греч. phago — ем) — захват и переваривание твердых пищевых частиц, и пиноцитоза (от греч. pino — пью) — захват и транспортировка жидкости.
На рисунке ниже показаны стадии фагоцитоза. Фагоцитоз был открыт Мечниковым И.И., создателем фагоцитарной теории иммунитета. Отмечу, что адгезия (от лат. adhaesio — прилипание) — сцепление между клеткой и твердой пищевой частицей (другой клеткой, например бактерией), которую она собирается поглотить.

Дыхание
Очевидно, что органов дыхания у простейших нет. Простейшие дышат всей поверхностью клетки.
Размножение
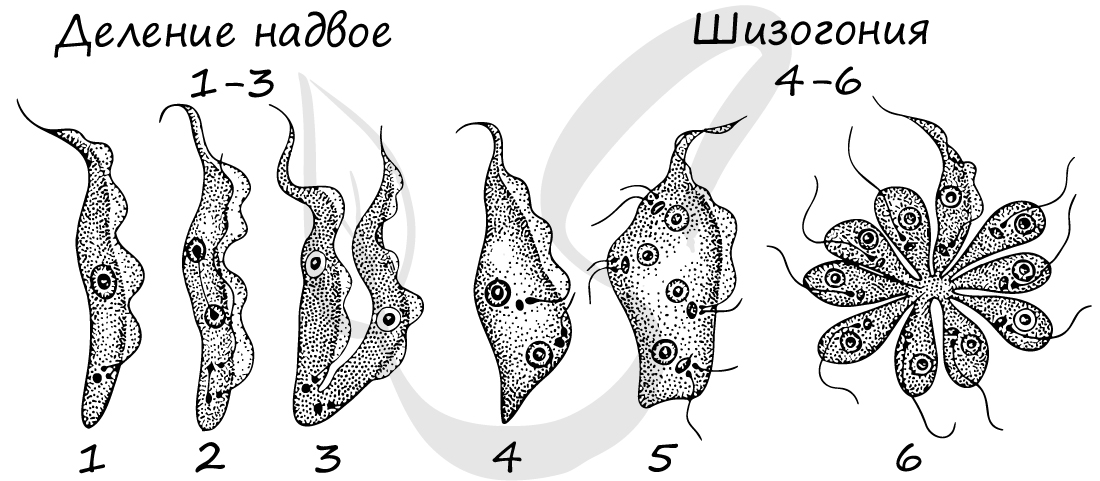
У простейших возможно бесполое и половое размножение. Бесполое осуществляется с помощью деления (митоз), шизогонией, спорообразованием (мейоз). Половое — с помощью копуляции и конъюгации.
Шизогония (от греч. schizo — разделяю) — множественное бесполое размножение, при котором, вследствие деления без разрыва цитоплазматической мембраны, клетка становится многоядерной, а затем распадается на множество дочерних клеток (соответственно количеству ядер).
Копуляция (от лат. copulatio — совокупление) — слияние как плазмы, так и ядер обеих копулирующих гаплоидных (n) особей.
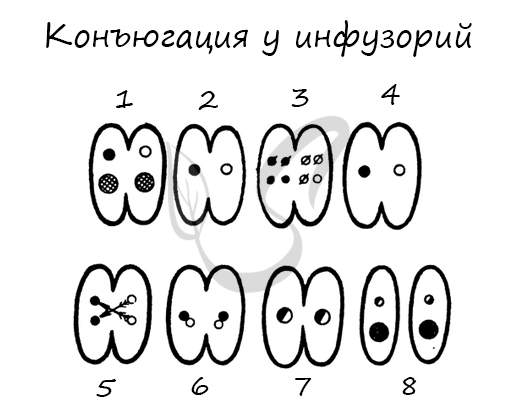
Конъюгация (от лат. conjugatio — соединение) — временное соединение двух особей, которые при этом обмениваются частями своего ядерного аппарата и цитоплазмой. В ходе конъюгации инфузорий объединяются их пронуклеусы, образовавшиеся в результате деления малого ядра (микронуклеуса) мейозом. После конъюгации происходит энергичное деление особей.
Значение простейших
Простейшие являются звеном в цепи питания. Фитопланктон (продуценты) — создатели органических веществ, служащие пищей для многих организмов. Зоопланктон (консументы) — питаются фитопланктоном и сами служат пищей для других организмов. Часть простейших являются причинами многих паразитарных заболеваний человека, растений и животных.

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Курс лекций по протозоологии издательско-полиграфический центр «Международный Соломонов университет»
| Назва | Курс лекций по протозоологии издательско-полиграфический центр «Международный Соломонов университет» |
| Сторінка | 12/18 |
| Дата | 11.09.2012 |
| Розмір | 2.16 Mb. |
| Тип | Курс лекций |
7. РАЗМНОЖЕНИЕ ПРОСТЕЙШИХДля размножения простейших характерны два типа процессов. Один из них — агамное или бесполое размножение — сводится к увеличению числа особей данного вида и может происходить различными способами. Особи, образованные в результате такого размножения также могут иметь разную судьбу. Они могут непосредственно вырастать в организм, подобный материнскому, проходить через метаморфоз (как бродяжки сукторий или перитрих), целиком превращаться в гамету, или гаметоцит (как спорозоиты грегарин). Кроме того, в жизненный цикл многих протист включается половой процесс. Он связан с образованием специальных стадий — гамет, которые могут формироваться разными способами. Половой процесс может вовсе не сопровождаться размножением (конъюгация, автогамия, партеногенез). Наиболее существенным моментом полового процесса является перестройка ядра и других компонентов клетки, протекающая через ряд фаз и приводящая к образованию нового поколения, отличающегося по своей жизнеспособности и наследственным свойствам от предыдущих поколений. Как уже отмечалось, не все типы полового процесса приводят к увеличению числа особей, поэтому в отношении простейших обычно говорят о наличии у них бесполого размножения с одной стороны и полового процесса с другой. БЕСПОЛОЕ РАЗМНОЖЕНИЕПростое деление надвое или монотомия. Этот тип размножения наиболее распространен у простейших и клеток тканей многоклеточных. В типичном случае оно представляет собой правильное равномерное разделение тела простейшего на две одинаковые дочерние особи, этому предшествует период питания и роста. Классический пример — размножение амеб. У амеб деление не имеет определенной ориентации. Тело амеб вытягивается, на экваторе намечается поперечная перетяжка, она постепенно углубляется и, наконец, клетка разделяется на две половины, которые немедленно начинают самостоятельно двигаться и питаться. Деление цитоплазмы сопровождается митотическим делением ядра. Остальные клеточные органоиды либо сами делятся, либо просто распределяются между дочерними особями, а недостающие образуются заново. Такое же неориентированное деление наблюдается у части солнечников и радиолярий. Наличие скелета, естественно, несколько усложняет этот процесс. У раковинных амеб — тестацей (Arcella) и громиид (Euglypha) перед делением в специальных вакуолях закладываются зачатки кремнеземных пластинок раковины. Затем начинается деление, плоскость которого расположена перпендикулярно продольной оси раковины. Ядро делится и примерно половина цитоплазмы вместе с дочерним ядром и кремнеземными пластинками выходит из устья наружу и окружается новой раковиной. У радиолярий, имеющих скелет из двух створок, положение плоскости деления определяется положением створок. При делении створки расходятся вместе с половиной цитоплазмы, и достраивается недостающая створка. У жгутиконосцев и цилиат положение плоскости деления более постоянно и стабильно. У жгутиконосцев оно, как правило, происходит в продольном направлении, у инфузорий — в поперечном. У жгутиконосцев деление начинается с переднего конца тела и продолжается к заднему концу. При этом бывает, что отделяющиеся половины клетки отгибаются в противоположные стороны, что создает впечатление поперечного деления. У динофлагеллят плоскость деления отклонена от продольной — деление происходит в косом направлении. Также в косом направлении проходит плоскость деления у опалин. У сложно организованных мастигофор деление связано с явлениями реорганизации. При этом некоторые органеллы резорбируются и формируются заново у дочерних особей. Подобные явления могут происходить с базальными зернами жгутика, аксостилем и жгутиками полимастигин. У инфузорий процесс деления приводит к еще более сложной реорганизации, чем у жгутиковых. Так у гипотрих у передней дочерней особи старые адоральные мембранеллы постепенно уменьшаются в размерах, и, в то же время, у их основания появляются зачатки новых мембранелл, которые растут по мере резорбции старых. Точно так же заменяются брюшные цирри, так что обе дочерние особи получают полностью новый набор локомоторных органелл. Особый случай реорганизации инфузорий — стоматогенез. Стоматогенез — морфогенетический процесс, который можно наблюдать при делении, регенерации инфузорий, а также при трансформации микростомных форм в макростомные. Первый этап стоматогенеза — образование новых кинетосом, составляющих зачаток околоротовой цилиатуры. Этот этап у разных групп инфузорий протекает по-разному, что служит основанием для филогенетических и систематических построений. Исходной формой стоматогенеза считается телокинетальный — образование зачатков цилиатуры, при котором кинетосомы происходят от передних отрезков соматических кинет или же соседних с ними фрагментов кинет. Он характерен для представителей класса Kinetofragminophorea. Многие гименостоматы, спиротрихи и перитрихи имеют буккокинетальный стоматогенез — новые кинетосомы возникают рядом с кинетосомами старого ротового аппарата. При этом базальные тела специализированных отделов перистома дают начало разным компонентам нового рта. Разделение двух новообразованных комплексов ресничек происходит путем серии сложных перемещений, типа сдвигов и поворотов. Паракинетальный стоматогенез встречается у некоторых гименостомат и спиротрих. При нем старые ротовые структуры рассасываются, новые возникают за счет (или вблизи) соматических ресничных рядов на вентральной стороне организма. Если не удается установить никакой связи между новыми кинетосомами и прежними ротовыми структурами, говорят об апокинетном стоматогенезе. Непосредственно после образования базальных телец на них уже обнаруживаются различные системы микротрубочек, как, например, нематодесмы, постцилиарные и трансверсальные микротрубочки. Затем кинетосомы мигрируют к плазмалемме, где начинается рост ресничек. На следующем этапе стоматогенеза кинетосомы выстраиваются в пределах зачатка в определенном порядке. Этот процесс происходит в определенном направлении и в строго определенной последовательности. В конце стоматогенеза развивается аппарат захват пищи — зона, от которой отшнуровываются пищевые вакуоли. Палинтомия. Палинтомией называется деление надвое, повторяющееся многократно, без промежуточной стадии питания и роста. Характерными чертами палинтомии являются: 1. Предшествующий гипертрофический рост материнской особи, что дает ей возможность делиться многократно. 2. Палинтомия очень часто наступает в периоды жизненного цикла, которым предшествует усиленное питание организма. 3. Палинтомия обычно характеризуется временной дедифференцировкой продуктов деления, так как продукты деления не успевают пройти процесс реорганизации. Поэтому палинтомия часто проходит на стадии цисты. Палинтомию подразделяют на линейную, когда дочерние клетки располагаются в виде цепочки (динофлагелляты, инфузории-апостоматы) и табличную, которая возникает в результате чередования продольных делений в двух взаимно перпендикулярных плоскостях, вследствие чего продукты деления располагаются параллельно друг другу в плоскую табличку. Такая палинтомия характерна для фитомонад, некоторых динофлагеллят и эвгленид. Часто палинтомия наблюдается при образовании гаметоцитов и гамет. Палиноспорогенез (видоизмененная палинтомия). Палиноспорогенез характерен для динофлагеллят, паразитирующих на сальпах, аппендикуляриях, морских копеподах и полихетах. В его основе лежит сочетание повторного ложнопоперечного деления с дифферецировкой на две неравноценные дочерние особи — трофоцит, который остается связанным с хозяином и гоноцит, продолжающий делиться путем линейной палинтомии. Трофоцит при этом питается и растет, отделяя от себя новые гоноциты. Таким путем создаются сложные комплексы клеток, состоящие из трофоцита и целой группы гоноцитов, находящихся на разных стадиях палинтомии. При этом достигается высокая продуктивность, что характерно для паразитов. Почкование. Почкование выводят из неравномерного деления. При этом морфологически различают материнскую особь (трофонт) и дочернюю (бродяжку, телотрох, почку, томит). Различают монотомическое почкование, когда образуется одна бродяжка, и множественное почкование. Кроме того, у разных групп инфузорий имеется внутреннее почкование, наружное и некоторые переходные формы почкования. Почкование характерно для сидячих инфузорий — сукторий, перитрих, хонотрих, ринходид, встречается у солнечников. Характерной особенностью почкования является отсутствие реорганизаций у материнских особей — они, как правило, сохраняют неизменными все, в том числе локомоторные или ротовые органеллы. Среди мастигофор почкование встречается крайне редко, в частности известно простое наружное почкование у Haplozoon и некоторых хризомонад. У солнечников известно множественное наружное почкование. У некоторых миксоспоридий отмечается обособление и отделение участков тела плазмодиальных стадий. ^ Синтомия или множественное деление . При этом типе размножения животное на определенном этапе жизненного цикла (обычно после периода питания и роста) испытывает серию повторных делений ядра и становится временно многоядерным, а затем сразу распадается на соответствующее количество клеток. В зависимости от того, образуются ли в результате процесса вегетативные особи или половые клетки, размножающаяся посредством синтомии стадия называется шизонтом или гамонтом, а когда множественное деление следует непосредственно за половым процессом — споронтом. Продукты шизогонии, соответственно, называются мерозоитами, гамогонии — гаметами, а спорогонии — спорозоитами для споровиков и зооспорами для других групп. Синтомия широко распространена среди споровиков, реже встречается у мастигофор (динофлагелляты — Noctiluca) и отсутствует у инфузорий. Плазматомия. Плазматомией называют тип бесполого размножения, при котором тело многоядерных простейших делится на две или большее число многоядерных же дочерних особей. При этом деление клетки происходит несогласованно с ядерными делениями и независимо от них. К плазматомии относят деление плазмодиев слизневиков, миксоспоридий, амеб — пеломиксин и др. Некоторые авторы ставят под сомнение выделение этого способа размножения в особый тип. ^ Размножение колоний. Еще один тип бесполого размножения. Выделяют три способа размножения колоний. Первый заключается в делении целой колонии надвое. Таким способом размножаются колониальные радиолярии и хризомонады. Второй способ сводится к отделению от колонии особей (бродяжек), которые закладывают новые колонии. Этот способ встречается у сидячих жгутиконосцев и перитрих (Zoothamnium arbuscula). Третий способ сходен со вторым, но здесь бродяжки не образуются, а особи-основательницы остаются в составе старой колонии и дают начало дочерней колонии. Он характерен для колониальных вольвоцин. Источник |



