
- Биология. 10 класс
- *§ 29—1. Понятие размножения. Типы размножения
- Понятие размножения
- Особенности деления прокариотических клеток: способы и механизмы клеточного деления
- Особенности деления прокариотических клеток
- Способы деления прокариотических клеток
- Механизмы клеточного деления
- Деление прокариотических клеток
- Способы деления прокариотических клеток
- Механизмы осуществления клеточного деления
- Готовые работы на аналогичную тему
- § 17. Простое бинарное деление. Митоз. Амитоз
- Оглавление
Биология. 10 класс
*§ 29—1. Понятие размножения. Типы размножения
Понятие размножения
Одним из основных отличительных свойств живой материи является размножение — способность живых организмов воспроизводить себе подобных. Благодаря этому свойству обеспечивается непрерывность жизни на Земле.
Размножение организмов определенного вида предоставляет возможность воспроизведения признаков не только родительских особей, но и вида в целом. А увеличение численности особей в результате размножения способствует расселению вида и расширению его ареала. Следовательно, благодаря размножению обеспечивается непрерывность и преемственность в передаче наследственной информации от родителей к потомству в ряду поколений и поддерживается длительное существование вида.
В основе размножения лежит деление клеток. Как вы уже знаете, наследственная информация в клетках хранится в хромосомах. При размножении организмов передача наследственной информации в дочерние клетки происходит с помощью хромосом, которые перед делением клеток удваиваются. В соматических клетках (клетках тела) хромосомы парные. В каждой паре одна хромосома отцовская, другая — материнская, следовательно, в соматических клетках находится двойной набор хромосом. Соматические клетки содержат наследственную информацию обоих родителей. В половых клетках (гаметах) хромосомы непарные, так как при образовании гамет парные хромосомы расходятся и попадают в разные клетки. Следовательно, гаметы содержат одинарный набор хромосом и несут наследственную информацию только одного из родителей.
Источник
Особенности деления прокариотических клеток: способы и механизмы клеточного деления
Особенности деления прокариотических клеток
Способы деления прокариотических клеток
Деление прокариотических клеток представляет собой процесс образования дочерних клеток прокариот на основе материнской клетки.
Есть 2 наиболее важных события, происходящих на протяжении жизненного цикла клеток прокариот. Это:
Практически всегда прокариотические клетки делятся так, что в результате образуются две одинаковые по размеру дочерние клетки. В некоторых случаях такой процесс называют еще бинарным делением или делением пополам.
В большинстве случаев прокариотические клетки характеризуются наличием клеточных стенок. Поэтому в результате бинарного деления образуется септа.
Септа — это определенная система перегородок, находящихся между дочерними клетками, имеющая свойство расслаиваться посередине.
В ходе деления различных бактериальных клеток наблюдаются определенные различия. Оригинальная система деления — система деления грамотрицательный бактерий. Этот механизм был открыт в результате исследований бактерий E. coli. У этих бактерий механизм деления нарушен. Внутри этих клеток можно наблюдать мутации, затрагивающие гены, которые и формируют механизм клеточного деления.
Происходит формирование определенных фенотипов:
- филаменты или длинные клетки. Они формируются, когда септа по определенным причинам не может сформироваться. Филаменты могут распределяться внутри клетки равномерно, иметь один нуклеоид или несколько;
- миниклетки или клетки минимального размера. Эти клетки лишены ДНК. Их образование происходит в случае, если в ходе клеточного деления формируется не одна, а несколько септ;
- безъядерные клети или клетки нормального размера. Они также лишены ДНК.
Механизмы клеточного деления
Один из механизмов деления, характерных для прокариотических клеток — молекулярный. В этом механизме септальное кольцо или кольцевая органелла играют ключевую роль. Кольцевая органелла находится посередине клетки и способна сокращаться — так образуются перетяжки между двумя дочерними клетками.
Зрелое септальное кольцо — это сложный белковый комплекс. В него входит большое количество разнообразных белков.
Входящие в состав септального кольца белки бывают нескольких разновидностей:
- модулирующие сборку филаментов;
- те, что связывают кольцо с мембраной;
- белки, которые координируют процесс образования септы с сегрегацией ДНК;
- те, что синтезируют пептидогликан;
- белки, гидролизующие пептидогликан.
У перечисленных белков есть своя неповторимая роль в процессе перераспределения генетической информации. Также все они выступают в качестве связующих звеньев в ходе деления клеток.
Функция септального кольца для многих белков все еще остается неопределенной.
Процесс формирования зрелой формы септального кольца характеризуется определенными особенностями.
После того как происходит деление, с помощью белка формируется спираль, прилегающая ко внутренней мембране. Эта спираль закручивается вдоль клеточной оси, постоянно меняет свое расположение и относительно быстро перемещается от одного полюса клетки к другому. Почти одновременно с этим завершается процесс репликации ДНК. Происходит захлопывание спирали и формирование Z-кольца посередине клетки.
Ученые считают, что Z-кольцо представлено короткой спиралью.
Следующий этап — созревание септального кольца. Этот процесс достаточно быстрый — продолжается от 14 до 21 минут. По истечении этого времени все ключевые белки присоединяются к Z-кольцу. Всего за 1-3 минут эти белки включаются в состав септального кольца.
До момента сборки септального кольца Z-кольцо отвечает за стимулирование синтезпептидогликана в центре клетки — в результате такого стимулирования клетка удлиняется.
Молекулярные основы описанного процесса все еще в полной мере не выяснены.
Белки, отвечающие за синтез полярных пептидогликанов включаются в септальное кольцо одними из последних. Одновременно с ними также включаются белки, обеспечивающие частичный гидролиз пептидогликана на границе раздела между клетками.
Завершает процесс клеточного деления формирование перетяжки и окончательное разделение двух дочерних клеток. Образование перетяжек имеет отношение ко всем компонентам клеточной оболочки: в частности, внутренней мембране, внешней мембране и слою пептидогликана.
Есть предположения, что Z-кольцо отвечает за инвагинацию внутренней части мембраны. Однако все еще остается непонятным механизм передачи напряжения на эту мембраны.
Одновременно с инвагинацией происходит синтез и преобразование пептидогликана септы с помощью ферментов септального кольца. Как только септа сформирована, в процесс деления клетки включаются ферменты пептидогликангидролазы, благодаря которым происходит отделение будущих дочерних клеток одна от другой.
Завершает процесс деления прокариотических клеток инвагинация и обособление внешних мембран дочерних клеток.
Из описанного выше ясно, что процесс деления клеток прокариот отличается собственными оригинальными чертами. Этот процесс называют прямым делением клетки или амитозом. Однако внутри такого деления происходят сложные преобразования белковых структур и реализация ДНК клетки.
Как вариант бинарного деления выступает почкование — многие рассматривают его как неравномерное бинарное деление. В результате почкования на одном полюсе материнской клетки формируется вырост или почка — в процессе роста она увеличивается. Как только эта почка достигает размеров материнской клетки, через некоторое время она от нее отделяется.
Прямым бинарным делением делятся клетки, дочерние клетки которых потом могут иметь довольно много морфологических и физиологических различий. Дочерняя и материнская клетки могут различаться, в том числе, своими размерами, однако генетический набор информации остается одинаковым и реализуется в последствии похожими принципами.
Такое деление дает возможность отслеживать процесс старения прокариотических клеток. Образованные в результате этого варианта деления дочерние клетки более жизнеспособные и лучше приспосабливаются к внешней среде.
Источник
Деление прокариотических клеток
Вы будете перенаправлены на Автор24
Деление прокариотических клеток – это процесс образования дочерних клеток прокариот из материнской клетки.
Способы деления прокариотических клеток
Ключевыми событиями в жизненном цикле клеток прокариот являются:
В большинстве случаев прокариотические клетки делятся с образованием двух идентичных по размеру дочерних клеток. Такой процесс иногда называют бинарным делением или делением пополам.
Чаще всего прокариотические клетки имеют клеточные стенки, поэтому бинарное деление сопровождается образованием септы.
Септа – это система перегородок между дочерними клетками, которая имеет свойство расслаиваться посередине.
Есть некоторые различия в процессе деления различных бактериальных клеток. Оригинальной является система деления грамотрицательных бактерий. Раскрытию данного механизма способствовало исследование бактерий E. сoli. Эти бактерии отличаются нарушенным механизмом деления. Внутри таких клеток происходят мутации, которые затрагивают гены, формирующие механизм клеточного деления. При этом формируется следующие фенотипы:
- филаменты или длинные клетки, сформированные в том случае, когда септа не образуется по тем или иным причинам. Филаменты могут быть равномерно распределенными внутри клетки, содержать единственный нуклеоид, содержать несколько нуклеоидов;
- миниклетки или клетки минимального размера, лишенные ДНК. Миниклетки образуются в том случае, если при клеточном делении формируется больше, чем одна септа;
- безъядерные клетки или клетки нормального размера, которые лишены ДНК.
Механизмы осуществления клеточного деления
Также для прокариотических клеток характерен молекулярный механизм деления. В таком случае центральную роль играет септальное кольцо или кольцевая органелла, которая располагается посередине клетки и имеет способность сокращаться, образуя перетяжки между двумя дочерними клетками.
Готовые работы на аналогичную тему
Зрелое септальное кольцо представляет собой сложный белковый комплекс, который состоит из большого количества разных белков.
Все белки, которые входят в состав септального кольца делятся на следующие разновидности:
- модулирующие сборку филаментов;
- связывающие кольцо с мембраной;
- координирующие процесс образования спеты с сегрегацией ДНК;
- синтезирующие пептидогликан;
- гидролизующие пептидогликан.
Все представленные типы белков играют собственную оригинальную роль в ходе перераспределения генетической информации и выполняют роль связующих звеньев в ходе деления клеток.
Для многих белков функция септального кольца до сих пор остается не определенной.
Процесс формирования зрелой формы септального кольца также имеет ряд собственных особенностей. После его деления белок формирует прилегающую ко внутренней мембране спираль, которая закручивается вдоль клеточной оси. Такая спираль постоянно меняет собственное положение и достаточно быстро перемещается от одного полюса клетки к другому. Примерно в это же время завершается процесс репликации ДНК, спираль захлопывается и формируется Z — кольцо посередине клетки. Многие ученые предполагают, что Z – кольцо на самом деле также представлено короткой спиралью.
Далее происходит процесс созревания спетального кольца. Он длится приблизительно 14 – 21 минуту и после прохождения данного времени к Z – кольцу присоединяются все ключевые белки. Все белки включаются в состав септального кольца в течение 1 – 3 минут.
До сборки септального кольца Z-кольцо стимулирует синтезпептидогликана в центре клетки таким образом, что клетка начинает удлиняться. Молекулярные основы данного процесса до сих пор не установлены в полной мере.
Одним из последних в септальное кольцо включаются белки, которые отвечают за синтез полярных пептидогликанов. А также белки обеспечивают частичный гидролиз пептидогликана на границе раздела между клетками.
Завершающим этапом клеточного деления называют формирование перетяжки и конечное разделение двух дочерних клеток. Образование перетяжек затрагивает все компоненты клеточной оболочки, а именно внутреннюю мембрану, внешнюю мембрану, слой пептидогликана.
Также существует мнение о том, что за инвагинацию внутренней части мембраны отвечает Z-кольцо. Но механизм передачи напряжения на эту мембрану до сих пор полностью не известен.
Одновременно с данным процессом ферменты септального кольца синтезируют или преобразуют пептидогликан септы. После формирования спеты в работу по делению клетки вступают ферменты пептидогликангидролазы, который помогает отделять будущие дочерние клетки друг от друга.
Процесс деления прокариотических клеток завершается инвагинацией и обособлением внешних мембран дочерних клеток.
Таким образом, процесс деления клеток прокариот имеет собственные оригинальные черты, сам механизм обычно называют прямым делением клетки или амитозом, но внутри него лежат процессы сложных преобразований белковых структур и реализации ДНК клетки.
Иногда вариантом бинарного деления является почкование, которое рассматривается как неравномерное бинарное деление. При почковании на одном полюсе материнской клетки образуется вырост или почка, которая увеличивается в процессе роста. Постепенно почка достигает размеров материнкой клетки, отделяясь от нее через некоторое время.
Равновеликое бинарное деление дает начало таким дочерним клеткам, которые в последствии могут иметь достаточно большое количество морфологических и физиологических различий. Дочерняя и материнская клетки могут отличаться также и размерами, но при этом генетический набор информации идентичен и реализуется в дальнейшем по схожим принципам.
При подобном делении появляется возможность отслеживания процесса старения прокариотических клеток. Такие дочерние клетки гораздо жизнеспособнее и лучше приспосабливаются к условиям окружающей среды.
Источник
§ 17. Простое бинарное деление. Митоз. Амитоз
| Сайт: | Профильное обучение |
| Курс: | Биология. 11 класс |
| Книга: | § 17. Простое бинарное деление. Митоз. Амитоз |
| Напечатано:: | Гость |
| Дата: | Четверг, 18 Ноябрь 2021, 01:20 |
Оглавление
Простое бинарное деление . Клетки прокариот в подавляющем большинстве случаев делятся с образованием двух одинаковых по размеру дочерних клеток. Этот процесс называется *простым бинарным делением* (простым делением надвое).
*Перед делением происходит репликация бактериальной хромосомы – кольцевой молекулы ДНК, которая в определенной точке прикреплена к плазмалемме. При этом образуются две идентичные дочерние хромосомы , также прикрепленные к цитоплазматической мембране (рис. 17.1). Затем специальные моторные белки перемещают дочерние хромосомы в противоположных направлениях, и расстояние между молекулами ДНК увеличивается.
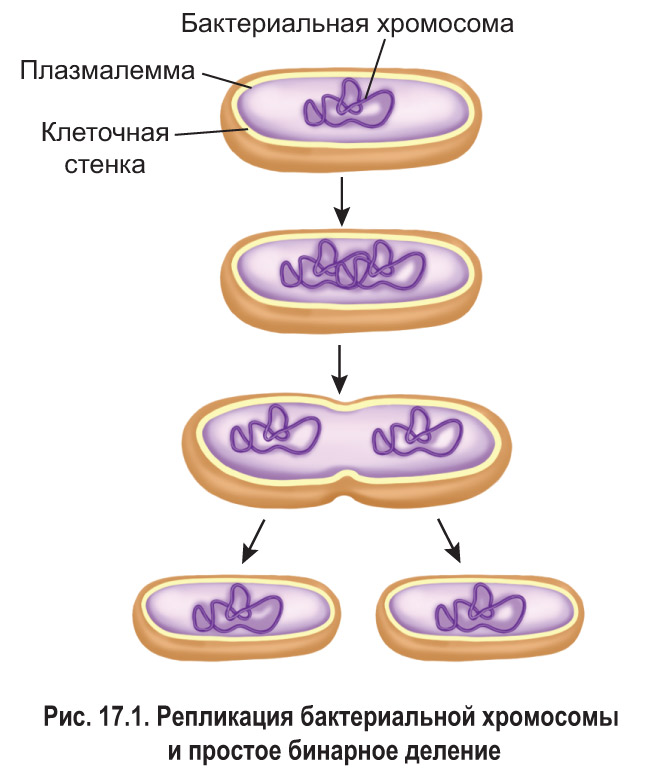
В процессе деления плазмалемма впячивается (врастает) внутрь клетки между двумя разошедшимися хромосомами. Одновременно в этой области происходит достраивание клеточных стенок будущих дочерних клеток. В конечном итоге бактериальная клетка разделяется надвое. При этом в каждой дочерней клетке оказывается по одной хромосоме. Простое бинарное деление свойственно только прокариотам.* Эукариотические клетки, в отличие от бактериальных, имеют более сложную организацию. Поэтому для них характерны другие способы деления: митоз, амитоз, мейоз.
Митоз — основной способ деления клеток эукариот, в результате которого из одной материнской клетки образуются две дочерние с таким же набором хромосом, как и в материнской клетке. Продолжительность митоза в среднем составляет 1—2 ч. *При этом клетки животных, как правило, делятся быстрее (30—60 мин), чем растительные (2—3 ч).* Митоз — непрерывный процесс, но для удобства его разделяют на несколько фаз. Митозом могут делиться клетки с различными наборами хромосом — гаплоидные (1n), диплоидные (2n), триплоидные (3n) и т. д. Рассмотрим процесс митотического деления на примере диплоидной клетки (табл. 17.1).
Таблица 17.1. Митоз
Схема
Фаза и процессы, происходящие в ней
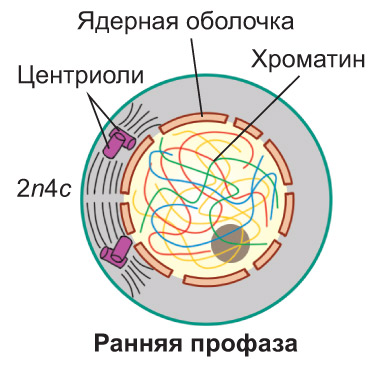
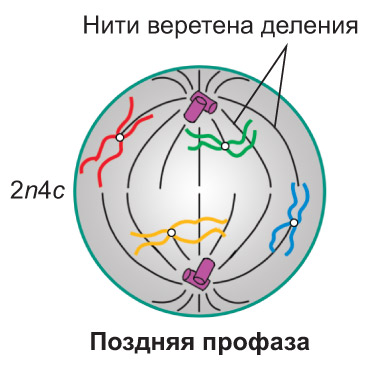
Профаза. В ядре клетки начинается спирализация хроматина , что приводит к формированию хромосом. *В начале профазы нити хроматина , спутанные в клубок, едва различимы в световой микроскоп. Постепенно они укорачиваются и одновременно утолщаются, так что к концу профазы уже можно четко видеть отдельные хромосомы .* Каждая из них состоит из двух сестринских хроматид, соединенных в области центромеры. *В этом участке хромосомы располагаются особые белковые структуры — кинетохоры, к которым впоследствии будут прикрепляться нити веретена деления. Как правило, каждая хроматида имеет по одному кинетохору.* По мере формирования хромосом исчезают ядрышки. Оболочка ядра распадается на мелкие фрагменты. Частично спирализованные хромосомы оказываются в гиалоплазме, располагаясь в ней беспорядочно (хаотически). Набор хромосом и хроматид в клетке можно выразить записью 2n4c.
Во время профазы два клеточных центра (удвоение этого органоида, как вы знаете, произошло в S-периоде интерфазы) инициируют образование микротрубочек. Из них начинает формироваться веретено деления. В процессе его образования центриоли попарно расходятся к разным полюсам клетки. *Часть нитей веретена деления направляется от одного полюса к противоположному. Другие нити прикрепляются к кинетохорам и способствуют перемещению хромосом в экваториальную плоскость клетки. Кроме того, формируются короткие микротрубочки, которые отходят от полюсов клетки радиально, образуя так называемую фигуру звезды.* В клетках, не имеющих клеточного центра (что характерно, например, для большинства растений), веретено деления формируется без участия центриолей. *В этом случае центрами организации микротрубочек являются бесструктурные аморфные зоны, расположенные на полюсах клетки. Их называют полярными шапочками*
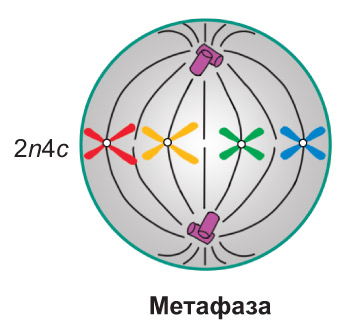
Метафаза. Завершается формирование веретена деления. Хромосомы располагаются в центральной части клетки, примерно на равном расстоянии от полюсов, *образуя так называемую метафазную пластинку*. При этом их центромеры находятся в экваториальной плоскости клетки, *а кинетохоры сестринских хроматид, связанные с нитями веретена деления, обращены к двум противоположным полюсам*. В метафазе спирализация хромосом достигает максимума. *Именно в этот период наиболее удобно изучать особенности строения хромосом и подсчитывать их общее количество в клетке*
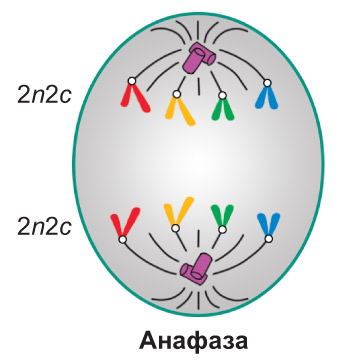
Анафаза. Центромера каждой хромосомы разделяется надвое, и сестринские хроматиды отделяются друг от друга. С этого момента их называют дочерними хромосомами. Нити веретена деления, прикрепленные к центромерам, укорачиваются, и дочерние хромосомы расходятся к противоположным полюсам клетки. *Как правило, при этом они принимают V-образную форму: центромеры обращены к полюсам, а плечи — к экваториальной плоскости клетки. Перемещение всех дочерних хромосом к полюсам происходит с одинаковой скоростью.* В конце анафазы у каждого полюса клетки оказывается идентичный набор дочерних хромосом (молекул ДНК) — 2n2c. *В клетках, не имеющих клеточной стенки (у животных и др.), анафаза сопровождается растяжением клетки в длину. Для растительных и других клеток, покрытых клеточной стенкой, такое явление не характерно*

Телофаза. Нити веретена деления постепенно разрушаются. Вблизи каждого полюса клетки происходит деспирализация (раскручивание) дочерних хромосом с образованием хроматина . Одновременно с этим вокруг деспирализующихся хромосом формируются оболочки двух новых ядер. Далее в образовавшихся ядрах возникают ядрышки. *На этом завершается деление ядра — кариокинез, и начинается деление цитоплазмы с образованием двух дочерних клеток — цитокинез. Перед цитокинезом органоиды клетки, как правило, более или менее равномерно распределяются по цитоплазме.*
В экваториальной плоскости клеток животных компоненты цитоскелета формируют кольцевую перетяжку. Она углубляется, пока не произойдет полное разделение двух дочерних клеток. Клетки растений в связи с наличием жесткой клеточной стенки делятся иначе. *В экваториальную плоскость растительной клетки направляются пузырьки комплекса Гольджи. Здесь они сливаются друг с другом. При этом содержимое пузырьков используется для построения так называемой срединной пластинки, а из мембран пузырьков по обе стороны от нее достраиваются цитоплазматические мембраны будущих дочерних клеток. Срединная пластинка служит основой для формирования клеточных стенок новых клеток. В отдельных ее участках имеются поры. Благодаря этому между образующимися дочерними клетками сохраняются тяжи цитоплазмы (цитоплазматические мостики), которые, как вы уже знаете, называются плазмодесмами. В конечном итоге срединная пластинка соединяется с клеточной стенкой материнской клетки, и материнская клетка оказывается разделенной на две дочерние*
Запомнить последовательность фаз митотического деления клетки поможет фраза «ПРОфессор МЕТнул АНАнас в ТЕЛефон» или слово «ПриМАТ».
Как уже отмечалось, при митозе образуются две дочерние клетки с одинаковым набором хромосом — таким же, как в исходной материнской клетке, вступавшей в митоз. Отличие заключается лишь в том, что каждая хромосома дочерней клетки, вступающей в интерфазу нового клеточного цикла, до репликации представлена не двумя идентичными хроматидами, а одной.
Биологическое значение митоза. В ходе митоза молекулы ДНК, которые содержались в ядре материнской клетки, точно и равномерно распределяются между дочерними. Следовательно, две новые клетки получают одинаковую наследственную информацию и оказываются генетически идентичными друг другу и материнской клетке. Таким образом, митотическое деление обеспечивает точную передачу генетической информации в ряду поколений клеток, обусловливает поддержание постоянного числа хромосом.
Благодаря митозу в многоклеточном организме происходит увеличение количества клеток. Это лежит в основе роста и развития всех многоклеточных организмов, а также обеспечивает процессы регенерации — восстановления поврежденных тканей и органов. Бесполое размножение многих организмов (деление одноклеточных протистов, почкование кишечнополостных, вегетативное размножение растений и т. д.) также обусловлено митотическим делением клеток.
*Нормальный ход митоза может быть нарушен различными внешними или внутриклеточными факторами. Например, действие ионизирующих излучений и определенных химических веществ может приводить к разрыву хромосом на отдельные фрагменты. При этом некоторые из них оказываются лишенными центромеры. Такие участки хромосом не могут связываться с нитями веретена деления и, следовательно, перемещаться под их действием. Если во время телофазы хромосомный фрагмент, лишенный центромеры, находится возле одного из полюсов, он может быть включен в состав одного из дочерних ядер. Если же подобный участок хромосомы располагается вблизи экваториальной плоскости клетки, вероятность его вхождения в ядро одной из формирующихся клеток практически равна нулю. Такие хромосомные фрагменты в дальнейшем подвергаются расщеплению.
Иногда из-за неправильного формирования нитей веретена деления сестринские хроматиды той или иной хромосомы , отделившиеся друг от друга, перемещаются к одному и тому же полюсу. В результате одна из дочерних клеток получает лишнюю хромосому (2n + 1), а в другой клетке, наоборот, одна из хромосом не будет иметь парной, гомологичной хромосомы (2n — 1). Действие на клетку некоторых химических веществ может приводить к изменению вязкости гиалоплазмы или нарушению процессов сборки и распада микротрубочек веретена деления. При этом в анафазе часто наблюдается несинхронное расхождение дочерних хромосом к полюсам. Одни из них движутся быстрее, а другие — медленнее. В итоге «опоздавшие» хромосомы не включаются в состав дочерних ядер, а образуют в цитоплазме так называемые микроядра, которые впоследствии разрушаются.
Известны химические соединения, которые разрушают нити веретена деления, но не влияют на процесс разделения сестринских хроматид. Одним из таких веществ является алкалоид колхицин. Его воздействие на клетку, находящуюся в метафазе митоза, приводит к распаду микротрубочек веретена деления. Далее сестринские хроматиды разделяются, но без участия веретена деления они не могут разойтись к полюсам и остаются в центральной части клетки. Через некоторое время вокруг этих хроматид (теперь уже дочерних хромосом) образуется общая ядерная оболочка, и они входят в состав одного ядра. Так возникает полиплоидная клетка, содержащая набор хромосом, удвоенный по сравнению с исходным.
Встречаются также случаи, когда в телофазе после образования двух ядер не происходит цитокинез. Вследствие этого клетка оказывается двуядерной.
Таким образом, нарушение правильного протекания митоза может приводить к неравномерному распределению генетического материала между дочерними клетками, возникновению полиплоидных или двуядерных клеток и т. д.*
Амитоз . При амитозе в клетке не формируется веретено деления, в ее ядре не происходит спирализация хроматина , сохраняются ядрышки и ядерная оболочка. Оставаясь в состоянии, характерном для интерфазы, ядро делится надвое перетяжкой. При этом молекулы ДНК ( хромосомы ) распределяются между дочерними ядрами неравномерно, случайным образом. В ряде случаев амитотическое деление ядра не сопровождается последующим *цитокинезом*. Это приводит к появлению двуядерных и даже многоядерных клеток. Если же дочерние клетки все-таки образуются, то клеточные компоненты, как и ДНК, распределяются между ними случайно и неравномерно.
Амитоз — сравнительно редкое явление. Этим способом делится, например, большое ядро инфузорий. У многоклеточных организмов амитоз наблюдается при различных патологических процессах (рост опухолей, воспаление и др.), а также в стареющих, обреченных на гибель клетках. Как правило, клетки, возникшие в результате амитоза, теряют способность вступать в нормальный клеточный цикл и в дальнейшем делиться митозом.

Для прокариотических клеток характерно простое бинарное деление, для эукариотических — митоз, амитоз и мейоз. Митоз — основной способ деления клеток эукариот, в результате которого из одной материнской клетки образуются две дочерние с таким же набором хромосом. В ходе митоза происходит спирализация хроматина с образованием компактных хромосом и формирование веретена деления. Затем сестринские хроматиды каждой хромосомы отделяются друг от друга и с помощью веретена деления равномерно распределяются между образующимися дочерними клетками. У многоклеточных организмов митотическое деление клеток обеспечивает процессы роста, развития и регенерации. В основе бесполого размножения многих организмов также лежит митоз.
При амитозе ядро клетки делится перетяжкой без спирализации хроматина и образования веретена деления. Это приводит к неравномерному распределению молекул ДНК между дочерними ядрами. Во многих случаях амитотическое деление ядра не сопровождается последующим разделением клетки на две дочерние.
 | 1. Какие способы деления характерны для клеток прокариот? Для эукариотических клеток? Амитоз , митоз, мейоз, простое бинарное деление. 2. Что такое митоз? Охарактеризуйте фазы митоза. 3. В связи с чем дочерние клетки, образовавшиеся в результате митоза, получают одинаковую наследственную информацию? В чем заключается биологическое значение митоза? 4. Установите соответствие между соматическими клетками человека, находящимися в различных периодах интерфазы и митоза, и количеством хромосом и хроматид в этих клетках. 5) У каждого полюса клетки в конце анафазы 6) В каждой дочерней клетке в конце телофазы а) 23 хромосомы , 23 хроматиды б) 23 хромосомы , 46 хроматид в) 46 хромосом, 46 хроматид г) 46 хромосом, 92 хроматиды 5*. В чем заключаются различия между митозом и амитозом? Как вы думаете, почему митоз называют непрямым делением клетки, а амитоз — прямым? 6*. В ядре неделящейся клетки наследственный материал (ДНК) находится в виде аморфного рассредоточенного вещества — хроматина . Перед делением хроматин спирализуется и образует компактные структуры — хромосомы , а после деления возвращается в исходное состояние. Для чего клетки совершают такие сложные видоизменения своего наследственного материала? Источник |



