
- Способы деления клетки делеция инверсия
- Небольшие делеции и инсерции
- Большие делеции и инсерции
- Эффекты рекомбинации
- Способы деления клетки делеция инверсия
- Учебное видео: митоз клетки и его стадии
- Способы деления клетки делеция инверсия
- Несбалансированные перестройки хромосом
- Делеции хромосом
- Дупликации хромосом
Способы деления клетки делеция инверсия
Мутации могут также вызываться инсерцией, инверсией, слиянием или делецией последовательности ДНК. Некоторые делеции и инсерции захватывают только несколько нуклеотидов и обычно легко обнаруживаются при секвенировании. В других случаях утрачивается, инвертируется, дублируется или транслоцируется значимый сегмент или целый ген, создавая новое размещение генной последовательности.
Такие мутации обычно обнаруживают на уровне блот-гибридизации ДНК пациента по Саузерну или ПЦР, выявляющими новые последовательности, созданные транслоцированным сегментом. В редких случаях делеции достаточно велики, чтобы быть видимыми на цитогенетическом уровне. Чтобы обнаруживаться даже с помощью высокоразрешающих методов окраски на стадии прометафазы, эти мутации обычно должны захватывать от 2 до 4 миллионов пар оснований ДНК.
Во многих примерах такие делеции захватывают более одного гена и вызывают синдромы генных последовательностей. Межхромосомные транслокации легче всего обнаружить методом SKY.
Небольшие делеции и инсерции
Некоторые делеции и инсерции влияют на незначительное число пар оснований. Когда число пар не кратно трем (т.е. захватывает не целое количество кодонов) и мутация находится в кодирующей последовательности, изменяется рамка считывания, начиная с участка инсерции или делеции. Такие мутации называются мутациями сдвига рамки.
В точке инсерции или делеции генерируется другая последовательность кодонов, кодирующая несколько аномальных аминокислот и завершающаяся стоп-кодоном в смещенной рамке. Если число оснований кратно трем, сдвига рамки не происходит, и в продукте гена обнаруживается инсерция или делеция соответствующих аминокислот.
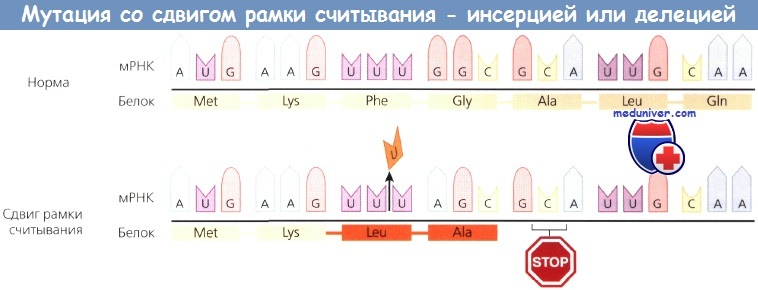 Мутация со сдвигом рамки считывания, при которой вставка (инсерция) или выпадение (делеция) пары оснований ведут к образованию стоп-кодона и, как правило, синтезу укороченного белка
Мутация со сдвигом рамки считывания, при которой вставка (инсерция) или выпадение (делеция) пары оснований ведут к образованию стоп-кодона и, как правило, синтезу укороченного белка
Большие делеции и инсерции
Изменения структурных генов, достаточно большие, чтобы выявляться блот-гибридизацией по Саузерну, встречаются сравнительно редко, но описаны при многих наследственных болезнях. Частота таких мутаций заметно отличается среди разных генетических заболеваний; некоторые характеризуются высокой частотой обнаруживаемых делеции, при других делеции бывают редкой причиной мутации.
Например, делеции большого гена дистрофина в Х-хромосоме при мышечной дистрофии Дюшенна или большого гена нейрофибромина при нейрофиброматозе I типа присутствуют более чем в 60% случаев. Большинство случаев а-талассемии — следствие делеции одного из двух генов b-глобина в хромосоме 16, тогда как b-талассемия только изредка вызывается делениями в гене b-глобина.
В некоторых случаях основа делеции гена хорошо известна и связана с нарушением рекомбинации между многочисленными копиями аналогичных или идентичных последовательностей ДНК. В других случаях причины делеции неизвестны.
Инсерция больших участков ДНК — причина мутаций, встречающихся значительно реже делеции. Тем не менее новый механизм мутации, инсерция последовательности LINE, описан в нескольких не родственных спорадических случаях у больных с гемофилией.
Семейство повторных последовательностей LINE представляет класс повторяющейся ДНК, которая может транскрибироваться в РНК, которая, в свою очередь, после обратной транскрипции генерирует последовательности ДНК, способные включать себя в различные места в геноме. У нескольких пациентов с гемофилией А в экзоне гена фактора VIII обнаружена прерывающая кодирующую последовательность и инактивирующая ген LINE последовательность длиной несколько килобаз.
Это обнаружение позволяет предполагать, что по крайней мере некоторые из 850 000 копий семейства LINE, имеющихся в геноме человека, способны вызвать болезнь за счет инсерции.
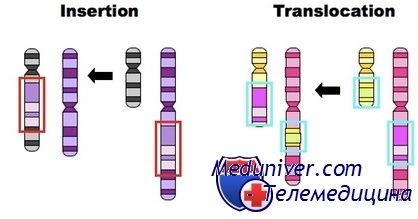
Эффекты рекомбинации
Важная причина мутаций при некоторых заболеваниях — делеции или дупликации, вызванные рекомбинацией весьма похожих или идентичных последовательностей ДНК. Например, как причина дупликации нескольких экзонов при семейной гиперхолестеринемии подтверждена рекомбинация между разными участками семейства диспергированных повторов класса Alu, располагающихся в интронах гена рецептора ЛПНП.
В других случаях ген может принадлежать семейству генов, представленных аналогичными копиями гена, располагающихся в хромосоме последовательно. Когда члены такого семейства генов располагаются тандемно в одном хромосомном регионе, они иногда неправильно спариваются или в мейозе (когда спариваются два гомолога), или после репликации в митозе (когда обмениваются ДНК две сестринские хроматиды).
Рекомбинация, происходящая между неправильно спаренными хромосомами или сестринскими хроматидами, может привести к делеции или дупликации гена. Считают, что неравный кроссинговер вызывает делецию одного из генов b-глобина при а-талассемии и изменения в числе копий гена зеленого пигмента в кластере генов красного и зеленого зрительного пигмента в Х-хромосоме как у людей с нормальным цветовым зрением, так и у мужчин со Х-сцепленными дефектами восприятия красного и зеленого цветов.
Возможны также аномальные спаривание и рекомбинация между двумя сходными последовательностями, повторяющимися в одной нити ДНК; в зависимости от ориентации последовательностей, такая рекомбинация может вести к делеции или инверсии. Например, почти половина всех случаев тяжелой гемофилии — следствие рекомбинации, инвертирующей множество экзонов и нарушающей структуру гена.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
Способы деления клетки делеция инверсия
Атипические митозы. Наряду с типичными картинами митоза нередко наблюдаются атипические митозы, возникновение которых обусловлено теми или иными отклонениями в деятельности указанных выше основных клеточных компонентов — участников митоза. Так, если по каким-либо причинам клеточный центр не разделится надвое, то образуется однополюсное веретено деления. Такой митоз называют однополюсным, униполярным (от лат. unus — один) или моноцентрическим (от греч. monos — один). При этом варианте митоза хромосомы не расходятся к полюсам веретена, и деление клетки не завершается плазмотомией.
Нередко встречаются многополюсные митозы, в процессе которых образуется несколько клеточных центров (до 5-6 и больше). Это связано с делением клеточного центра (центросомы) не на две, как обычно, а на большее число центриолей. По числу полюсов различают многополюсные, или мультиполярные (от лат. multum — много), или полицентрические (от греч. poly — много), а митозы называют три-, квадри-, гекса- и т. д. полярными. Чаще всего многополюсный митоз не завершается плазмотомией, вследствие чего образуются многоядерные гигантские клетки. Если же наступает цитотомия, то дочерние клетки содержат неравноценный генетический материал (анэуплоидия).
Ряд алкалоидов (колхицин) избирательно действует на веретено деления, в результате чего исключается возможность расхождения хромосом в анафазе. В этом случае течение митоза приостанавливается на стадии метафазы. Указанная разновидность атипического митоза в литературе известна под названием колхицинового митоза, или К-митоза. Колхицин используют в экспериментальной цитологии при подсчете количества митозов в тканях. Фибриллярные структуры веретена деления нарушаются также под действием бета-меркаптоэтанола, эфира, нагревания и повышенного давления.
Отклонения от нормального течения митоза могут быть связаны с изменениями структуры хромосом. Эти изменения называются хромосомными аберрациями (от лат. aberratio — уклонение). Различают несколько разновидностей хромосомных аберраций: 1) слипание хромосом; 2) разрыв хромосомы на фрагменты; 3) выпадение участка хромосомы при ее разрыве (нехватка, или делеция); 4) транслокация — обмен обломками между двумя хромосомами; 5) инверсия — переворачивание на 180° фрагмента хромосомы, расположенного между двумя переломами; 6) дупликация — удвоение какого-либо участка хромосомы; 7) отставание хромосом во время анафазного их перемещения с образованием хромосомных мостиков между расходящимися к полосам комплексами хромосом.
Из обособившихся при этом хромосом могут возникать маленькие ядра — кариомеры. Хромосомные аберрации возникают при действии на клетки ионизирующего излучения.
Эндомитоз. Эндомитозом (от греч. endon — внутри) называют вариант митоза, когда репликация хромосом не сопровождается исчезновением ядерной оболочки и образованием веретена деления. При эндомитозе в одних случаях хромосомы выявляются, в других же — они не видны. Путем повторных эндомитозов количество хромосомных наборов в клеточном ядре может значительно увеличиваться. Ядро приобретает гигантские размеры. Эндомитоз лежит в основе полиплоидии.
Полиплоидией (от греч. poly — много и ploos — складывать) называют такое состояние клетки, когда в ней в результате предшествующих эндомитозов оказывается более двух гаплоидных наборов хромосом. Полиплоидизация, в отличие от митоза, осуществляется без снижения специфических клеточных функций и свойственна полифункциональным элементам (клеткам печени, сердца, слюнных желез и др.). В зависимости от числа хромосомных наборов в полиплоидных клетках их называют три-(при 3), тетра-(при 4), пента-(при 5) и т. д. плоидными. Полиплоидные клетки отличаются гигантскими размерами. Они довольно часто встречаются в опухолевых тканях, а также в тканях, подвергнутых действию проникающей радиации.
Среди модификаций митоза имеется еще один особый его вариант, называемый мейозом (от греч. meiosis — уменьшение). В результате мейоза происходит уменьшение числа хромосом вдвое (от диплоидного к гаплоидному). Этот способ клеточного деления характерен для половых клеток.
Амитоз — прямое деление клетки (ядра). При этом происходит перешнуровывание или фрагментация ядра без выявления хромосом и образования веретена деления. Одной из форм амитоза может быть сегрегация геномов — множественное перешнуровывание полиплоидного ядра с образованием мелких дочерних ядер. Как правило, амитоз встречается в полиплоидных, отживающих или патологически измененных клетках и ведет к образованию многоядерных клеток. В последние годы факт существования амитоза как способа нормальной репродукции клеток отрицается.
Учебное видео: митоз клетки и его стадии

— Вернуться в оглавление раздела «гистология»
Источник
Способы деления клетки делеция инверсия
Структурные перестройки происходят вследствие разрывов хромосом с последующим соединением в аномальной комбинации. Хотя перестройки могут происходить многими способами, все вместе они бывают реже, чем анеуплоидий; в целом структурные аномалии присутствуют примерно у 1 из 375 новорожденных.
Хромосомные перестройки происходят спонтанно с низкой частотой, а также могут быть вызываны повреждающими агентами (кластогенами), например ионизирующим излучением, некоторыми вирусными инфекциями, многими химическими веществами. Подобно аномалиям числа хромосом, структурные перестройки могут присутствовать или во всех клетках человека, или в мозаичной форме.
Структурные перестройки определяют как сбалансированные, если хромосомный набор имеет нормальный состав хромосомного материала, или несбалансированные, если отмечен дополнительный или отсутствующий материал. Некоторые перестройки стабильны и способны проходить через митотические и мейотические деления клетки неизменными, другие — неустойчивы. Чтобы быть стабильной, перестроенная хромосома должна иметь функциональную центромеру и две теломе-ры.
Несбалансированные перестройки хромосом
При несбалансированных перестройках фенотип бывает аномальным из-за делеции или дупликации, а в некоторых случаях из-за их сочетания. Дупликация части хромосомы приводит к частичной трисомии; делеция — к частичной моносомии. Любое изменение, нарушающее нормальный баланс функциональных генов, может заканчиваться аномальными проявлениями.
Большие делеции или дупликации, включающие несколько миллионов пар оснований, могут быть обнаружены при обычном хромосомном исследовании, в том числе при кариотипировании с высоким разрешением. Для обнаружения малых делеции или дупликаций обычно показаны более сложные анализы, включая FISH или микрочиповый анализ.
Важный класс несбалансированных перестроек — субмикроскопические изменения области теломеры у пациентов с идиопатической умственной отсталостью. У нескольких процентов таких пациентов обнаружены небольшие делеции, дупликации и транслокации. Прицельный цитогенетический или геномный анализ теломерных и субтеломерных участков методами FISH или CGH-матриц может помочь в выяснении причин необъяснимой умственной задержки с целью генетического консультирования.
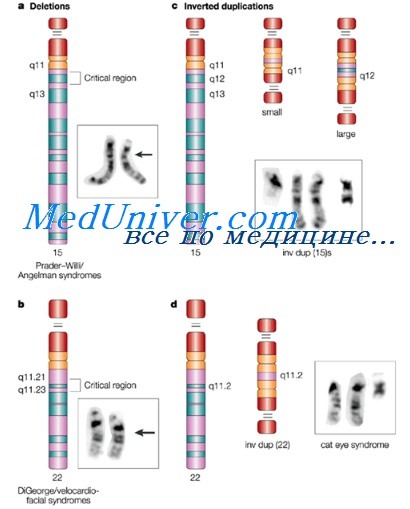
Делеции хромосом
Делеция — потеря сегмента хромосомы, приводящая к хромосомному дисбалансу. Носитель хромосомной делеции (с одним нормальным гомологом и одним делетированным) моногамен по генетической информации в соответствующем сегменте нормального гомолога.
Клинические последствия обычно отражают гаплонедостаточность (буквально — невозможность единственной копии генетического материала нормально выполнять функции, выполняемые двумя копиями) и зависят от размера удаленного сегмента, количества и функций содержащихся в нем генов. Цитогенетически видимые аутосомные делеции наблюдают с частотой приблизительно 1 на 7000 живых новорожденных. Малые, субмикроскопические делеции, обнаруживаемые микроматричным (биочиповым) анализом, более часты, но, как упоминалось раньше, клиническое значение большинства таких вариантов пока еще не полностью определено.
Делеция может находиться в конце хромосомы (терминальная) или внутри хромосомного плеча (интерстициальная). Делеции могут быть следствием простого разрыва хромосомы с последующей утратой ацентрического сегмента. Кроме того, в некоторых случаях к делециям может приводить неравный кроссинговер между гомологичными хромосомами или хроматидами.
Делеции также могут порождаться аномальным расхождением сбалансированной транслокации или инверсии, как показано ниже. При обследовании пациентов с дисморфиями и при пренатальной диагностике выявлено множество делеций. Знание функций генов, входящих в делетированные сегменты, и их влияние на фенотипические последствия существенно выросли в ходе реализации проекта «Геном человека».
Методы окрашивания высокого разрешения и FISH могут выявлять малые делеции, невидимые на обычных метафазных пластинках. Если цитогенетическая окраска высокого разрешения позволяет идентифицировать делецию размерами не менее нескольких миллионов пар оснований, то делеций, не обнаруживаемые кариотипическим методом, и неопределенные делеций с фенотипическими последствиями стандартно обнаруживают методами FISH и биочиповым анализом с использованием зондов, специфичных для интересующей области.
Дупликации хромосом
Дупликации, подобно делециям, могут происходить из-за неравного кроссинговера или ошибок расхождения хромосом в мейозе у носителей транслокаций или инверсий. В общих чертах дупликации оказываются менее вредными, чем делеций. Тем не менее, поскольку дупликация в гамете приводит к хромосомному дисбалансу (т.е. к частичной трисомии) и поскольку хромосомные разрывы могут нарушать гены, дупликации часто приводят к различным фенотипическим аномалиям.
Хотя описано много дупликаций, достаточно изучены только некоторые из них. Тем не менее с дупликациями конкретных регионов хромосом связаны определенные фенотипы. Например, дупликация всего или части короткого плеча хромосомы 12р ведет к развитию синдрома Паллистера-Киллиана, при котором у пациентов отмечают характерные краниофациальные черты, умственную задержку и целый спектр других врожденных дефектов, вероятно, обусловленный трисомией или тетрасомиеи конкретного гена, расположенного в дуплицированной области.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник



