
- Как регулируется активность ферментов?
- Способы регуляции активности ферментов
- 1.Доступность субстрата или кофермента
- 2. Компартментализация
- 3. Генетическая регуляция
- 4. Ограниченный (частичный) протеолиз проферментов
- 5. Аллостерическая регуляция
- 6. Белок-белковое взаимодействие
- 7. Ковалентная (химическая) модификация
- Активаторы и ингибиторы ферментов
Как регулируется активность ферментов?
Активность ферментов в клетке непостоянна во времени. Ферменты чутко реагируют на ситуацию, в которой оказывается клетка, на факторы, воздействующие на нее как снаружи, так и изнутри. Главная цель такой чувствительности ферментов – отреагировать на изменение окружающей среды, приспособить клетку к новым условиям, дать должный ответ на гормональные и иные стимулы, а в некоторых ситуациях – предоставить клетке шанс выжить.
Способы регуляции активности ферментов
В клетке имеется несколько способов регуляции активности ферментов – одни способы подходят для любых ферментов, другие более специфичны.
1.Доступность субстрата или кофермента
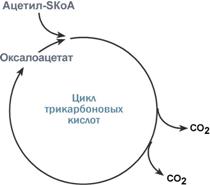
Роль оксалоацетата для работы ЦТК
Здесь работает закон действия масс – фундаментальный закон химической кинетики: при постоянной температуре скорость химической реакции пропорциональна произведению концентрации реагирующих веществ. Или упрощенно – скорость, с которой вещества реагируют друг с другом, зависит от их концентрации. Таким образом, изменение количества хотя бы одного из субстратов прекращает или начинает реакцию.
Например, для цикла трикарбоновых кислот (ЦТК) таким субстратом является оксалоацетат (щавелевоуксусная кислота). Наличие оксалоацетата «подталкивает» реакции цикла, что позволяет вовлекать в окисление молекулы ацетил-SКоА.
2. Компартментализация
Компартментализация – это сосредоточение ферментов и их субстратов в одном компартменте (одной органелле) – в эндоплазматическом ретикулуме, митохондриях, лизосомах, ядре, плазматической мембране и т.п.
3. Генетическая регуляция
Генетическая регуляция (изменение количества фермента) может происходить в результате увеличения или снижения его синтеза. С этой точки зрения ферменты можно подразделить на три группы:
- Конституитивные – такие ферменты, которые образуются в клетке постоянно, независимо от наличия субстрата (нейрональная NO-синтаза, ферменты гликолиза, β-окисления жирных кислот, репарации ДНК).
- Индуцируемые – синтез этих ферментов возрастает при наличии соответствующих стимулов — индукторов.
- Репрессируемые – образование таких ферментов в клетке при необходимости подавляется.
Изменение скорости синтеза фермента (индукция или репрессия) обычно зависит от количества определенных гормонов или метаболитов процесса.
Примеры индуцируемых ферментов:
- гормоны глюкокортикоиды стимулируют синтез ферментов глюконеогенеза, что обеспечивает стабильность концентрации глюкозы в крови при длительном голоданиии и устойчивость ЦНС к стрессу,
- исчезновение пищеварительных ферментов при длительном голодании и индукция их синтеза в восстановительный период в результате возобновления секреции гормонов ЖКТ,
- при беременности и после родов в молочной железе индуцируется синтез фермента лактозосинтазы под воздействием лактотропного гормона,
- токсические субстраты (например, этанол и барбитураты) стимулируют в печени синтез «своего» изофермента цитохрома Р450, который окисляет и обезвреживает эти вещества,
- при активации цитокинами моноцитов и макрофагов в них начинается синтез индуцируемой NO-синтазы.
Примеры репрессируемых ферментов:
- в печени репрессия фермента синтеза холестеролаГМГ-SKoA-редуктазы под влиянием холестерина и желчных кислот,
- в печени репрессия синтеза ферментов глюконеогенеза под действием инсулина,
- подавление синтеза триптофана бактериями при деятельности триптофанового оперона.
4. Ограниченный (частичный) протеолиз проферментов
Ограниченный (частичный) протеолиз проферментов подразумевает, что синтез некоторых ферментов осуществляется в виде более крупного предшественника и при поступлении в нужное место этот фермент активируется через отщепление от него одного или нескольких пептидных фрагментов. Подобный механизм защищает внутриклеточные структуры от повреждений.
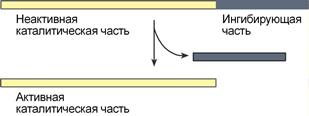
Схема активации фермента способом
«ограниченного протеолиза»
Примером служит активация протеолитических ферментов желудочно-кишечного тракта (трипсиноген, пепсиноген, прокарбоксипептидазы), факторов свертывающей системы крови, лизосомальных ферментов (катепсины).
Секреция ряда ферментов за пределы клетки в неактивном состоянии позволяет предохранить клетки от повреждения (пищеварительные ферменты) или сохранить белок в плазме крови до наступления определенного момента (факторы свертывания крови, белки системы комплемента, калликреин-кининовой и ренин-ангиотензиновой систем).
5. Аллостерическая регуляция
Аллостерические ферменты построены из двух и более субъединиц: одни субъединицы содержат каталитический центр, другие имеют аллостерический центр и являются регуляторными. Присоединение эффектора к аллостерической (регуляторной) субъединице изменяет конформацию белка и, соответственно, активность каталитической субъединицы.
Аллостерические ферменты обычно стоят в начале метаболических путей, и от их активности зависит течение многих последующих реакций. Поэтому они часто называются ключевыми ферментами.
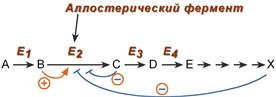
Общий принцип аллостерической регуляции
В качестве отрицательного регулятора может выступать конечный или промежуточный метаболит биохимического процесса или продукт данной реакции, т.е включается механизм обратной отрицательной связи . Если регуляторами являются начальный метаболит или субстрат реакции, то говорят о прямой регуляции , она может быть как положительной, так и отрицательной. Также регулятором могут быть метаболиты биохимических путей, каким то образом связанных с данной реакцией.
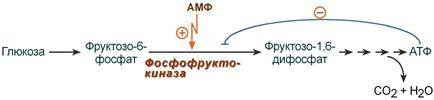
Регуляция фосфофруктокиназы конечным продуктом
Например, фермент энергетического окисления глюкозы, фосфофруктокиназа, регулируется промежуточными и конечными продуктами этого распада. При этом АТФ, лимонная кислота, фруктозо-1,6-дифосфат являются ингибиторами, а фруктозо-6-фосфат и АМФ – активаторами фермента.
- В большинстве клеток организма (кроме печени) при регуляции синтеза холестерола аллостерическим ингибитором ключевого фермента этого процесса ГМГ-КоА-редуктазы выступает сам холестерол, что быстро и точно регулирует его количество,
- Фермент ЦТК изоцитрат-дегидрогеназа активируется при помощи АМФ и АДФ и ингибируется влияниями АТФ и НАДН.
6. Белок-белковое взаимодействие
Термин белок-белковое взаимодействие обозначает ситуацию, когда в качестве регулятора выступают не метаболиты биохимических процессов, а специфичные белки. В целом ситуация схожа с аллостерическим механизмом: после влияния каких-либо факторов на специфичные белки изменяется активность этих белков, и они, в свою очередь, воздействуют на нужный фермент.
1. К примеру, мембранный фермент аденилатциклаза является чувствительным к воздействию мембранного G-белка, который сам активируется при действии на клетку некоторых гормонов (например, адреналина и глюкагона).
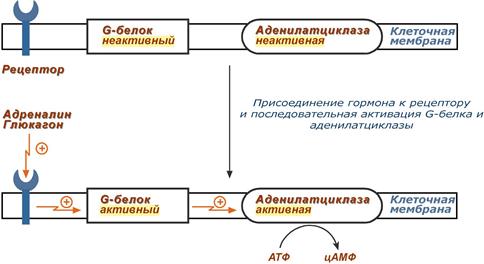
Упрощенная схема активации аденилатциклазы
Более подробно механизм активации G-белка можно посмотреть здесь.
2. Еще примером белок-белкового взаимодействия может быть регуляция активности протеинкиназы А через механизм ассоциации-диссоциации .
Протеинкиназа А является тетрамерным ферментом, состоящим из 2 каталитических (С) и 2 регуляторных (R) субъединиц. Активатором для протеинкиназы А является цАМФ. Присоединение цАМФ к регуляторным субъединицам фермента вызывает их отхождение от каталитических субъединиц. Каталитические субъединицы при этом активируются.
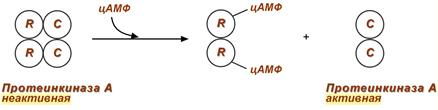
Активация протеинкиназы А при помощи цАМФ
7. Ковалентная (химическая) модификация
Ковалентная модификация заключается в обратимом присоединении или отщеплении определенной группы, благодаря чему изменяется активность фермента. Чаще всего такой группой является фосфорная кислота, реже метильные и ацетильные группы. Фосфорилирование фермента происходит по остаткам серина и тирозина. Присоединение фосфорной кислоты к белку осуществляют ферменты протеинкиназы, отщепление – протеинфосфатазы.
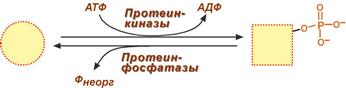
Изменение активности фермента
при фосфорилировании-дефосфорилировании
Ферменты могут быть активны как в фосфорилированном , так и в дефосфорилированном состоянии .
Например, в мышцах ферменты гликогенфосфорилаза и гликогенсинтаза
- при нагрузке фосфорилируются, при этом фосфорилаза гликогена становится активной и начинает расщепление гликогена и сжигание глюкозы, а гликогенсинтаза при этом неактивна.
- во время отдыха при синтезе гликогена оба фермента дефосфорилируются, синтаза при этом становится активной, фосфорилаза – неактивной.
Источник
Активаторы и ингибиторы ферментов
Активаторы и ингибиторы ферментов
Регуляция активности ферментов может осуществляться путём взаимодействия ферментов с различными биологическими компонентами или чужеродными соединениями, которые называются регуляторами ферментов. Они могут либо ускорять, либо замедлять ферментативную реакцию.
Активаторы – это вещества, увеличивающие скорость ферментативной реакции.
1. Вещества, влияющие на область активного центра. К ним относятся ионы металлов (Na + , K + , Fe 2+ , Co 2+ , Cu 2+ , Ca 2+ , Zn 2+ , Mg 2+ , Mn 2+ и др.). В ряде случаев ионы металлов выполняют функцию кофактора фермента. В других случаях они способствуют присоединению субстрата к активному центру фермента. Ионы металлов оказываются активаторами только в условиях дефицита их в организме.
2. Аллостерические эффекторы, которые связываются с аллостерическим (регуляторным) участком апофермента. Это связывание вызывает конформационные изменения в молекуле белка, приводящие к изменению структуры активного центра, что сказывается на связывании и превращении субстрата в активном центре. При этом активность фермента либо увеличивается (это аллостерические активаторы), либо уменьшается (это аллостерические ингибиторы). Аллостерическими эффекторами ферментов наиболее часто выступают различные метаболиты, а также гормоны, ионы металлов, нуклеозиды — АТФ, АДФ, АМФ.
3. Вещества, вызывающие модификации, не затрагивающие активный центр фермента. Возможно несколько вариантов таких модификаций:
— активация путём присоединения специфической модифицирующей группы к молекуле фермента. Пример: регуляция активности липазы.

 неактивная АТФ АДФ активная О
неактивная АТФ АДФ активная О


 ─СН2ОН ─СН2─О─Р─ОН
─СН2ОН ─СН2─О─Р─ОН
 фосфатаза │
фосфатаза │
В этом случае фосфатная группа присоединяется к гидроксильным группам аминокислот, находящихся в белковой части фермента. Отрицательно заряженные фосфатные группы могут разрывать слабые водородные и ионные связи в третичной структуре белка-фермента и влиять на конформационное состояние его активного центра. В зависимости от природы фермента фосфорилирование может его активировать или, наоборот, инактивировать. Реакции присоединения фосфатной группы катализируют ферменты протеинкиназы, а отщепления – фосфатазы. Активность этих ферментов в свою очередь находится под контролем гормональной системы.
— активация путём перехода неактивного предшественника — профермента в активный фермент за счёт частичного протеолиза.
Некоторые ферменты синтезируются в клетке первоначально неактивными и после секреции из клетки переходят в активную форму. Неактивные предшественники называются проферменты (зимогены). Под действием активатора происходит частичный гидролиз профермента с отщеплением от него неактивного пептида, в результате чего открывается активный центр. Так происходит активация ферментов желудочно-кишечного тракта, переваривающих белки пищи. Например, фермент пепсиноген, синтезированный в клетках желудка, затем в просвете желудка под действием соляной кислоты превращается в активный пепсин путём удаления неактивного участка полипептидной цепи:
неактивный HCl активный
п епсиноген пепсин + пептид
епсиноген пепсин + пептид
— активатор вызывает диссоциацию субъединиц фермента, имеющего четвертичную структуру (отщепление одной из субъединиц фермента).
Ингибиторами называют вещества, вызывающие снижение активности фермента. Следует различать инактивацию и ингибирование фермента. Сам по себе факт торможения ферментативной реакции в присутствии какого-либо вещества ещё не говорит о том, что это вещество – ингибитор. Любые денатурирующие агенты вызывают инактивацию фермента и торможение ферментативной реакции. Ингибиторы, в отличие от денатурирующих агентов, действуют в малых концентрациях и вызывают специфическое снижение ферментативной активности.
По прочности связывания с ферментом ингибиторы делятся на обратимые и необратимые. Необратимые ингибиторы прочно связываются с ферментом, тогда как комплекс фермент – обратимый ингибитор непрочен. Если сильно разбавить раствор фермента с обратимым ингибитором, то их комплекс распадается и активность фермента восстанавливается.
По механизму действия ингибиторы делятся на конкурентные и неконкурентные. Конкурентные ингибиторы имеют структурное сходство с молекулой субстрата, что позволяет им занять место субстрата в активном центре фермента:
Встраиваясь вместо субстрата в активный центр, такой ингибитор не даёт ферментативной реакции осуществиться. То есть, субстрат конкурирует с ингибитором за активный центр. С активным центром связывается то соединение, молекул которого больше. Снять конкурентное ингибирование можно, увеличив концентрацию субстрата.

На принципе конкурентного ингибирования основано действие многих фармакологических препаратов (например, сульфаниламидных), инсектицидов, фосфорорганических боевых отравляющих веществ (зарин, зоман).
Неконкурентные ингибиторы не имеют структурного сходства с субстратами. Они или связываются с каталитическими группами активного центра фермента, или, связываясь с ферментом вне активного центра, изменяют конформацию активного центра таким образом, что это препятствует превращению субстрата. Поскольку неконкурентный ингибитор не влияет на связывание субстрата, то в отличие от конкурентного ингибирования наблюдается образование тройного комплекса:
К неконкурентным ингибиторам относятся ионы тяжёлых металлов: ртути, свинца, кадмия, мышьяка. Они блокируют SH-группы, входящие в каталитический участок фермента. Снять действие неконкурентного ингибитора избытком субстрата, как при конкурентном ингибировании, нельзя, а можно лишь веществами, связывающими ингибитор (реактиваторами). Тяжелые металлы лишь в небольших концентрациях играют роль ингибиторов, в больших концентрациях они действуют как денатурирующие агенты.

Наиболее важными неконкурентными ингибиторами являются образующиеся в живой клетке промежуточные продукты метаболизма, способные обратимо связываться с аллостерическими участками фермента – аллостерические ингибиторы. Они занимают ключевое положение в метаболизме, поскольку тонко реагируют на изменения в обмене веществ и регулируют прохождение веществ по целой системе ферментов. Например, аллостерическая регуляция проявляется в виде ингибирования конечным продуктом первого фермента цепи. Эта регуляция сходна с регуляцией по механизму обратной связи и позволяет контролировать выход конечного продукта, в случае накопления которого прекращается работа первого фермента цепи:

 А → В → С → D
А → В → С → D

Е1, Е2, Е3 – ферменты; А, В, С, D — метаболиты
Источник



