
- Взаимодействие генов
- Способ взаимодействия неаллельных генов
- Комплементарность
- Эпистаз
- ДОМИНАНТНЫЙ ЭПИСТАЗ
- РЕЦЕССИВНЫЙ ЭПИСТАЗ
- Полимерия
- РЕШЕНИЕ
- ОТЛИЧИЕ КУЛЬТУРНЫХ ФОРМ ОТ ДИКИХ
- основные методы селекции
- ПОДБОР РОДИТЕЛЬСКИХ ПАР
- РАБОТЫ И. В. МИЧУРИНА
- ОТБОР
- ГИБРИДИЗАЦИЯ
- ИСКУССТВЕННЫЙ МУТАГЕНЕЗ
- ПОЯВЛЕНИЕ ПОЛИПЛОИДИИ
- ШЕЛКОПРЯДЫ АСТАУРОВА
- биотехнология
- ГЕННАЯ И КЛЕТОЧНАЯ ИНЖЕНЕРИЯ
- работы Н. И. Вавилова
- УЧЕНИЕ ОБ ИММУНИТЕТЕ РАСТЕНИЙ
- ЦЕНТРЫ ПРОИСХОЖДЕНИЯ КУЛЬТУРНЫХ РАСТЕНИЙ
Взаимодействие генов
Вы уже знаете о том, что гены могут взаимодействовать друг с другом по типу полного и неполного доминирования. Однако, в генетике встречается масса других примеров взаимодействия генов. В этой статье мы затронем те, которые ранее не обсуждались.
Кодоминирование
Кодоминирование — взаимодействия аллельных генов, при котором в гетерозиготном состоянии могут оказаться два доминантных гена одновременно, при этом каждый ген отвечает за свой признак.
Наиболее распространенным примером кодоминирования является наследование групп крови у человека.

Решим пару задач, которые укрепят понимание темы.
Пример решения задачи №1
«Родители имеют II и III группы крови, гетерозиготны. Какие группы крови можно ожидать у их детей?»
Гетерозиготный генотип матери — I A i 0 и генотип отца — I B i 0 . Составим схему решения для такого случая.

Итак, в результате такого брака может получиться ребенок с любой группой крови, в чем мы убедились.
Пример решения задачи №2
«Дигетерозиготная по B (III) группе и положительному резус-фактору вступила в брак с таким же мужчиной. Какое расщепление по фенотипу можно ожидать у детей?»
Сходу понятно, что гетерозиготы по III (B) группе крови будут записаны I B i 0 . Резус-фактор для нас новое понятие — это белок, находящийся на поверхности эритроцита (тогда говорят, что резус-фактор положителен), или отсутствующий (тогда у человека резус-фактор считается отрицательным). Генотипы записываются так:
- Резус-фактор положителен: Rh + Rh + , Rh + rh —
- Резус-фактор отрицателен: rh — rh —
В данной задаче сказано, что «дигетерозиготна по . и положительному резус-фактору» — значит, резус фактор будет записывать в генотипе — Rh + rh — .

Обратите внимание, что ошибкой является записать рецессивный ген перед доминантным. За такое могут снять балл на экзамене: aA, bB, i 0 I A , rh — Rh + . Правильный вариант записи: Aa, Bb, I A i 0 , Rh + rh — .
Каждая особь образует 4 гаметы, поэтому потомков получается 16. Подсчитает расщепление по фенотипу:
- 9 потомков : положительный резус-фактор, III (B) группа крови
- 3 потомка : положительный резус-фактор, I (0) группа крови
- 3 потомка : отрицательный резус-фактор, III (B) группа крови
- 1 потомок : отрицательный резус-фактор, I (0) группа крови
Расщепление по фенотипу в данном случае получилось: 9:3:3:1. Здесь проявляется III закон Менделя — закон независимого наследования, так как гены, отвечающие за группу крови и резус-фактор, находятся в разных хромосомах.
Комплементарность
Комплементарность — тип взаимодействия неаллельных генов, при котором развитие признака определяется не одной, а двумя или более парами неаллельных генов, располагающихся в разных хромосомах.
Неаллельные гены — это гены, расположенные в разных локусах хромосом, которые отвечают за разные признаки. В генетике случается такое, что один неаллельный ген может влиять на другой (ген a подавляет действие гена B). В этом разделе статьи мы подробно разберемся с подобным взаимодействием и рассмотрим задачи, которые могут встретиться.
Таким образом, развитие признака определяется именно сочетанием генов друг с другом. Здесь логичнее подчеркнуть совместное действие генов, нежели чем сказать, что доминантный ген подавляет рецессивный — при комплементарности это не совсем так.
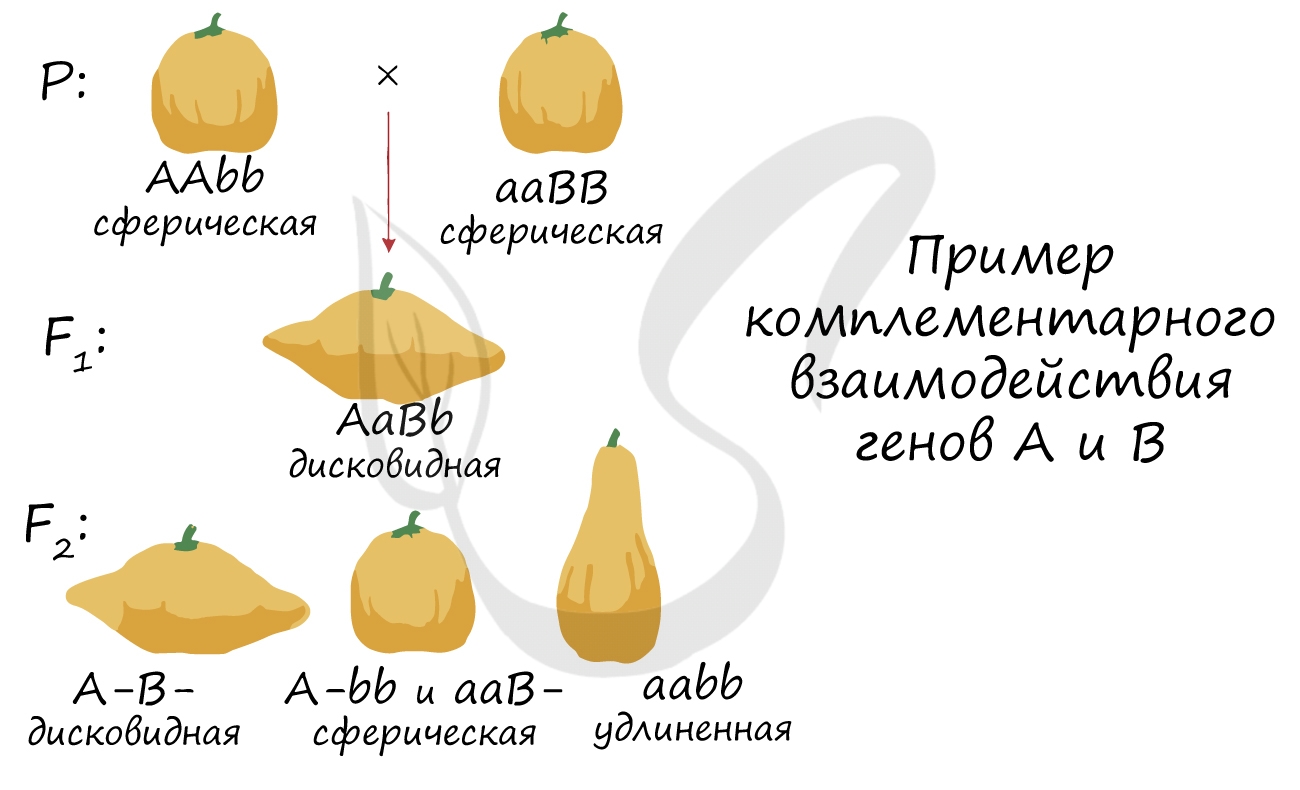
В каждой задаче свой случай комплементарного взаимодействия генов. Чтобы успешно их решать, надо помнить, что такое явление, как комплементарность, в принципе, возможно, и быть внимательным при написании генотипов особей и их гамет.
Пример решения задачи №3
Наследование слуха у человека определяется двумя доминантными генами из разных аллельных пар, один из которых детерминирует развитие слухового нерва, а другой – улитки. Определить вероятность рождения глухих детей, если оба родителя глухие, но по разным генетическим причинам (у одного отсутствует слуховой нерв, у другого улитка). По генотипу оба родителя являются дигомозиготными.

Здесь проявляется I закон Менделя — закон единообразия гибридов первого поколения. Возможен только один вариант генотипа ребенка от такого брака. У ребенка будет развит и слуховой нерв, и улитка — ребенок не будет глухим, в отличие от родителей.
Эпистаз
Эпистаз (противоположное действие генов) — явление, при котором один ген аллельной пары (супрессор) в доминантном (доминантный эпистаз) или рецессивном (рецессивный эпистаз) состоянии может подавлять развитие признака, за развитие которого отвечает другая пара генов.
Широко известным примером рецессивного эпистаза является Бомбейский феномен, названный так в результате зафиксированного случая в индийском городе Бомбеи. Доктор Бхенде обнаружил, что у людей рецессивных по гену h (hh) на поверхности эритроцитов не синтезируются агглютиногены — в результате этого они могут быть универсальными донорами.
Говоря проще о Бомбейском феномене: у людей с генотипом hh всегда обнаруживается первая группа крови при любом генотипе — I A I A , I B I B , I A I B . Ген h подавляет гены I A и I B — на поверхности эритроцитов не образуются агглютиногены A и B.
Пример решения задачи №4
«Редкий рецессивный ген (h) в гомозиготном состоянии обладает эпистатическим действием по отношению к генам I A , I B и изменяет их действие до I группы крови (бомбейский феномен). Определите возможные группы крови у детей, если у мужа II гомозиготная, у жены IV и оба родителя гетерозиготны по эпистатическому гену»
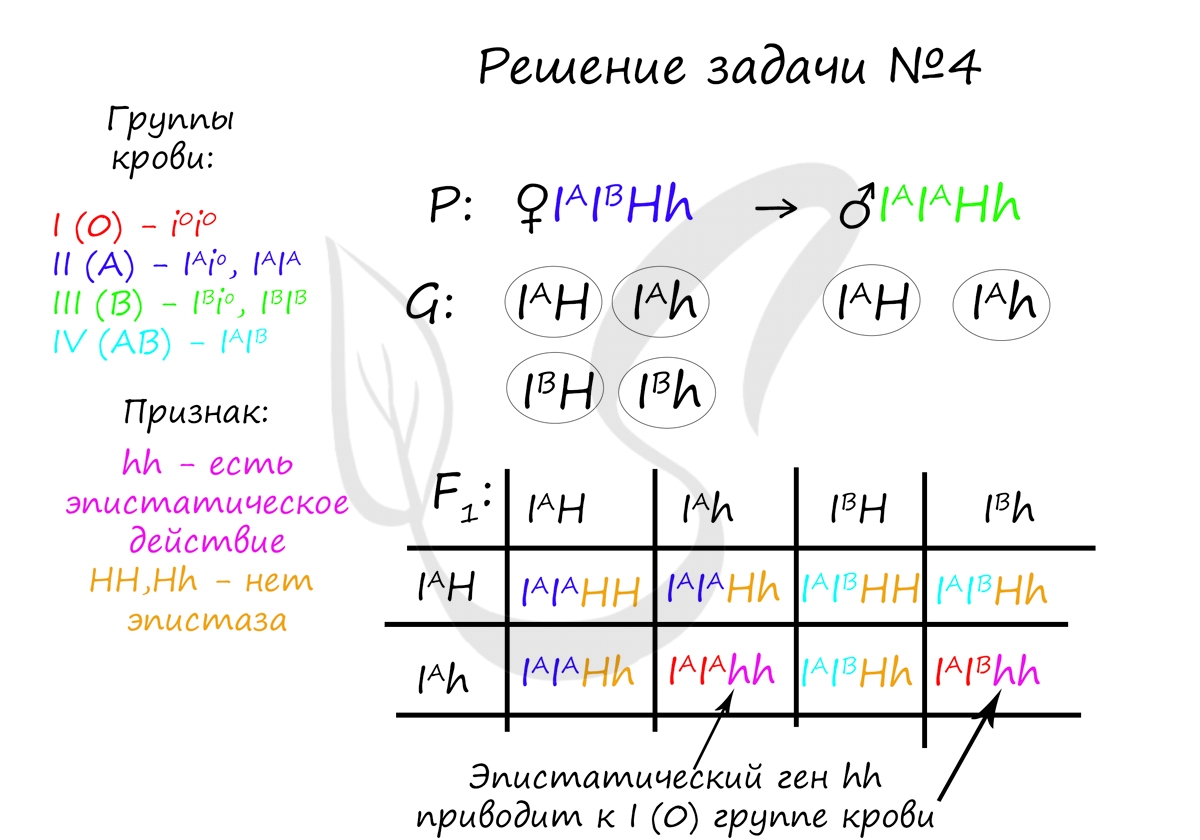
Вероятность рождения детей с i(0) группой крови в данном случае равна 2/8, или 1/4 (25%). Генотипами, у которых будет i(0) группа крови являются: I A I A hh и I A I B hh. Эпистатический рецессивный ген hh в гомозиготном состоянии всегда приводит к i(0) группе крови.
Полимерия
Полимерией называют зависимость определенного признака организма от нескольких пар аллельных генов, обладающих схожим действием. Такие гены называются полимерными. Часто выраженность признака зависит от соотношения доминантных и рецессивных аллелей — то есть чем больше доминантных генов, тем более выражен признак.

У человека полимерное действие генов заложено в наследовании количественных признаков (вес, рост, цвет кожи, давление).
Пример решения задачи №5
«Цвет кожи у мулатов наследуется по типу полимерии. При этом данный признак контролируется 2 аутосомными несцепленными генами. Сын белой женщины и негра женился на белой женщине. Может ли этот ребенок быть темнее своего отца?»
В данном случае полимерия проявляется в том, что чем больше доминантных генов в генотипе (A и B), тем более темный цвет кожи имеет человек. Это правило мы и применим для решения.
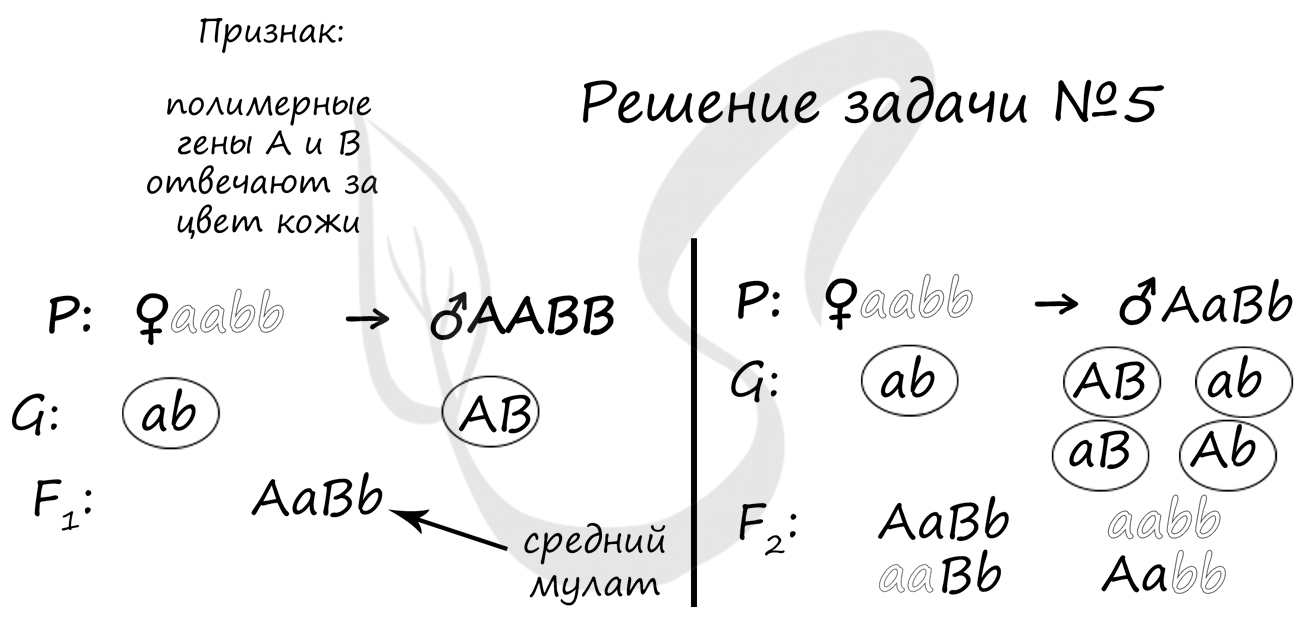
В результате первого брака (вспоминаем закон единообразия Менделя) получается AaBb — средний мулат. По условиям задачи он берет в жены белую женщину aabb. Очевидно, что в этой семье ребенок не может быть темнее своего отца: дети могут быть средними мулатами (AaBb), как отец, светлыми мулатами (aaBb, Aabb), либо белыми, как мать (aabb).
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Способ взаимодействия неаллельных генов
Очень редко развитие того или иного признака определяется одним геном. Часто один ген обусловливает развитие нескольких признаков. Такое явление называется плейотропным действием гена. Распространено и обратное — один признак контролируется несколькими генами.
Неаллельные гены — это гены, расположенные в различных участках (локусах) хромосом и кодирующие неодинаковые белки. Неаллельные гены могут взаимодействовать между собой, когда один признак проявляется под действием совокупности нескольких генов.
Выделяют три формы взаимодействия неаллельных генов:
Комплементарность
К комплементарным, или дополняющим друг друга, генам относятся такие неаллельные гены, которые при совместном проявлении обусловливают развитие нового признака. На примере наследования окраски цветков у душистого горошка можно понять сущность комплементарного действия генов. При скрещивании двух растений с белыми цветками у гибридов F1 цветки оказались пурпурными. При самоопылении растений из F1 в F2 наблюдалось расщепление растений по окраске цветков в отношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/16 растений, белые у 7/16.

Объяснение такого результата состоит в том, что каждый из доминантных генов не может вызвать появление окраски, определяемой пигментом антоцианом. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника пигмента — пропигмента. Ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Цветки душистого горошка с генотипом ааВВ и ААbb имеют белый цвет: в первом случае есть фермент, но нет пропигмента, во втором — есть пропигмент, но нет фермента, переводящего пропигмент в пигмент.

Проведем скрещивание двух растений душистого горошка с белыми цветками. В одной из линий, АAbb, есть доминантный аллель А, а у другой, ааBB, есть доминантный аллель В. У растений, имеющих доминантные гены А и В одновременно, есть и пропигмент (обеспечиваемый А), и фермент (обеспечиваемый В), необходимые для образования пурпурного пигмента. У гибридов F1 генотип AaBb, есть оба доминантных гена, поэтому они имеют пурпурные цветки. При самоопылении этих растений получаем F2. Результаты отражены в решётке Пеннета, розовым выделены генотипы тех растений, которые будут иметь пурпурные цветки.
| гаметы | AB | Ab | aB | ab |
| AB | AABB | AABb | AaBB | AaBb |
| Ab | AABb | AAbb | AaBb | Aabb |
| aB | AaBB | AaBb | aaBB | aaBb |
| ab | AaBb | Aabb | aaBb | aabb |
Комплементарное взаимодействие может давать не два, а три фенотипа. Это происходит в тех случаях, когда двойные рецессивные гомозиготы aabb по фенотипу отличаются от рецессивных гомозигот только по одному гену — aaBb, Aabb. В этом случае расщепление будет 9:6:1. Например, это наблюдается при наследовании формы плодов у тыквы:

Возможно и возникновение четырёх фенотипов. Например, при скрещивании кур с различной формой гребня.

aabb — простой (листовидный)
Эпистаз
Эпистаз — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I.
Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным.
ДОМИНАНТНЫЙ ЭПИСТАЗ
При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3. Последний вариант возникает, когда рецессивная гомозигота по гипостатичному гену фенотипически неотличима от фенотипа I (например, обе формы неокрашены).

Наследование окраски шерсти у собак (пример доминантного эпистаза):
A — черная окраска, а — коричневая, I — подавляет окраску, i — не подавляет.
РЕЦЕССИВНЫЙ ЭПИСТАЗ
Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена в гомозиготном состоянии аллелей гипостатичного гена (ii > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7. Последний вариант возникает, когда рецессивная гомозигота по гипостатичному гену фенотипически неотличима от фенотипа ii (например, обе формы неокрашены).
Рецессивный эпистаз проявляется при наследовании окраски шерсти у домовых мышей.
А — окраска агути (рыжевато-серая)
а — черная окраска
В — способствует проявлению окраски
b — супрессор (подавляет действие А и а)
Мыши с генотипом А-bb и ааbb имеют одинаковый фенотип — все белые.

Полимерия
Полимерия — взаимодействие неаллельных генов, при котором степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного гена имеют одинаковый нижний индекс.
Например, у пшеницы А — тёмно-красный цвет зёрен, а — белый цвет зёрен. За цвет отвечают два гена — 1 и 2. Первый ген может быть представлен аллелями А1 и а1, второй — аллелями А2 и а2.
В зависимости от того, каких аллелей больше — А или а, оттенок зёрен будет изменяться.

Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от числа доминантных аллелей всех генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1 для двух генов, 64:1 для трёх генов и т. д. Например, цвет кожи у людей зависит от четырёх генов, поэтому от брака двух мулатов практически никогда не рождаются дети с белой кожей.
Разберём решение задач с неаллельным взаимодействием генов.
При скрещивании двух божьих коровок с красными надкрыльями все гибриды первого поколения имели оранжевые надкрылья. Их скрестили между собой и получили во втором поколении 115 жуков с оранжевыми надкрыльями, 79 с красными надкрыльями и 13 с жёлтыми надкрыльями. Как наследуется цвет надкрылий?
РЕШЕНИЕ
Так как в первом поколении все потомки одинаковы, родители гомозиготны. Появление во втором поколении третьей окраски и численные соотношения указывают на то, что это дигибридное скрещивание. Значит, в потомстве имеется 16 генотипических классов. Общее число потомков второго поколения = 115 + 79 + 13 = 207. Размер генотипического класса = 207 : 16 = 13. С оранжевой окраской 115 : 13 = 9 генотипических классов. С красной окраской 79 : 13 = 6 генотипических классов. С жёлтой окраской 13 : 13 + 1 генотипический класс. Таким образом, во втором поколении расщепление 9:6:1, что соответствует комплементарному взаимодействию генов. Родительские формы ААbbи ааВВ, гибриды первого поколения АаВb. Во втором поколении 9 А__В_ : 6 (ааВ_ + А_bb) :1 ааbb.
Селекция — наука, разрабатывающая пути создания новых и улучшения существующих сортов растений, пород животных и штаммов микроорганизмов.
Создание новых сортов и пород основывается на таких важнейших свойствах живого организма, как наследственность и изменчивость. Именно поэтому генетика — наука об изменчивости и наследственности организмов — является теоретической основой селекции.
Имея свои собственные задачи и методы, селекция твердо опирается на законы генетики, является важной областью практического использования закономерностей, установленных генетикой. Вместе с тем селекция опирается и на достижения других наук. На сегодняшний день генетика вышла на уровень целенаправленного конструирования организмов с нужными признаками и свойствами.
Сорт, порода и штамм — устойчивая группа организмов, искусственно созданная человеком и имеющая определенные наследственные особенности.
Все особи внутри породы, сорта и штамма имеют сходные, наследственно закрепленные морфологические, физиолого-биохимические и хозяйственные признаки и свойства, а также однотипную реакцию на факторы внешней среды.
Основные направления селекции:
- высокая урожайность сортов растений, плодовитость и продуктивность пород животных;
- улучшение качества продукции (например, вкус, внешний вид плодов и овощей, химический состав зерна — содержание белка, клейковины, незаменимых аминокислот и т. д.);
- физиологические свойства (скороспелость, засухоустойчивость, зимостойкость, устойчивость к болезням, вредителям и неблагоприятным климатическим условиям).
- выведение стрессоустойчивых пород (для разведения в условиях большой скученности — на птицефабриках, фермах и т. п.);
- пушное звероводство;
- рыбоводство — разведение рыбы в искусственных водоемах.
ОТЛИЧИЕ КУЛЬТУРНЫХ ФОРМ ОТ ДИКИХ
| Культурные формы | Дикие формы |
| развиты признаки, полезные для человека и часто вредные в естественных условиях | наличие признаков, неудобных для человека (агрессивность, колючесть и т. п.) |
| высокая продуктивность | низкая продуктивность (мелкие плоды; низкая масса, яйценоскость, удойность) |
| хуже адаптируются к меняющимся условиям среды | высокая адаптивность |
| не имеют средств защиты от хищников и вредителей (горьких или ядовитых веществ, шипов, колючек и т. п.) | наличие естественных защитных приспособлений , повышающих жизнестойкость, но неудобных для человека |
основные методы селекции
Основные методы селекции:
ПОДБОР РОДИТЕЛЬСКИХ ПАР
Данный метод применяется прежде всего в селекции животных, т. к. для животных характерно половое размножение и немногочисленное потомство.
Выведение новой породы — процесс длительный, требующий больших материальных затрат. Это может быть целенаправленное получение определенного экстерьера(совокупности фенотипических признаков), повышение молочности, жирности молока, качества мяса и т. д.
Разводимые животные оцениваются не только по внешним признакам, но и по происхождению и качеству потомства. Поэтому необходимо хорошо знать их родословную. В племенных хозяйствах при подборе производителей всегда ведется учёт родословных, в которых оцениваются экстерьерные особенности и продуктивность родительских форм в течение ряда поколений.
РАБОТЫ И. В. МИЧУРИНА
ОТБОР
Искусственный отбор — сохранение для дальнейшего размножения особей с интересующими селекционера признаками. Формы отбора: массовый и индивидуальный.
- Интуитивный (бессознательный) отбор — самая древняя форма отбора, используемая ещё древним человеком: отбор особей по фенотипу, т.е. с наиболее полезными сочетаниями признаков.
- Методический отбор — отбор для размножения особей с чётко определёнными признаками, согласно цели и с учетом их фенотипов и генотипов.
- Массовый отбор — устранение из размножения особей, не имеющих ценные признаки, либо имеющих нежелательные признаки (например, агрессивных).
Массовый отбор может быть эффективен в том случае, если отбираются качественные, просто наследуемые и легко определяемые признаки. Массовый отбор обычно проводят среди перекрестноопыляемых растений. При этом селекционеры отбирают растения по фенотипу с интересующими их признаками. Недостаток массового отбора заключается в том, что селекционер не всегда может определить лучший генотип по фенотипу.
- Индивидуальный отбор — выделение отдельных особей с интересующими человека признаками и получение от них потомства.
Индивидуальный отбор более эффективен при отборе особей по количественным, сложно наследуемым признакам. Этот вид отбора позволяет точно оценить генотип благодаря анализу наследования признаков у потомства. Индивидуальный отбор применяют по отношению к самоопыляемым растениям (сорта пшеницы, ячменя, гороха и др.).
ГИБРИДИЗАЦИЯ
В селекционной работе с животными применяют в основном два способа скрещивания: инбридинг и аутбридинг.
Инбридинг — скрещивание близкородственных форм: в качестве исходных форм используются братья и сестры или родители и потомство.
Результат: получение гомозиготных организмов → разложение исходной формы на ряд чистых линий.
Минусы: пониженная жизнеспособность (рецессивные гомозиготы зачастую несут наследственные заболевания).
Такое скрещивание в определённой степени аналогично самоопылению у растений, которое также приводит к повышению гомозиготности и, как следствие, к закреплению хозяйственно ценных признаков у потомков. При этом гомозиготизация по генам, контролирующим изучаемый признак, происходит тем быстрее, чем более близкородственное скрещивание используют при инбридинге. Однако гомозиготизация при инбридинге, как и в случае растений, ведет к ослаблению животных, снижает их устойчивость к воздействию среды, повышает заболеваемость.
В селекции инбридинг обычно является лишь одним из этапов улучшения породы. За ним следует скрещивание разных межлинейных гибридов, в результате которого нежелательные рецессивные аллели переводятся в гетерозиготное состояние и вредные последствия близкородственного скрещивания заметно снижаются.
Аутбридинг — неродственное скрещивание между особями одной породы или разных пород животных в пределах одного вида.
Результат: получение большого количества гетерозиготных организмов → поддержание полезных качеств и усиление их выраженности в ряду следующих поколений.
Отдалённая гибридизация — получение межвидовых и межродовых гибридов.
Отдалённая гибридизация в селекции животных применяется значительно реже, чем в селекции растений.
Межвидовые и межродовые гибриды животных и растений чаще всего бесплодны, так как нарушается мейоз и гаметогенез не происходит. При этом восстановление плодовитости у животных представляет более сложную задачу, поскольку получение полиплоидов на основе умножения числа хромосом у них невозможно.
Преодоление бесплодия межвидовых гибридов растений впервые удалось осуществить в начале 20-х годов ХХ века советскому генетику Г. Д. Карпеченко при скрещивании редьки и капусты. Это вновь созданное человеком растение не было похоже ни на редьку, ни на капусту. Стручки занимали как бы промежуточное положение и состояли из двух половинок, из которых одна напоминала стручок капусты, другая — редьки. Каждая из исходных форм имела в половых клетках по 9 хромосом. В этом случае клетки полученного от них гибрида имели 18 хромосом. Но некоторые яйцеклетки и пыльцевые зёрна содержали все 18 хромосом (диплоиды), а при их скрещивании создано растение с 36 хромосомами, которое оказалось плодовитым. Так была доказана возможность использования полиплоида для преодоления нескрещиваемости и бесплодия при отдалённой гибридизации.
Бывает, что бесплодны особи только одного пола. Например, у гибридов высокогорного быка яка и рогатого скота бесплодны (стерильны) самцы, а самки плодовиты (фертильны).
Но иногда гаметогенез у отдалённых гибридов протекает нормально, что позволило получить новые ценные породы животных. Примером являются архаромериносы, которые, как и архары (горные бараны), могут пастись высоко в горах, а как мериносы дают хорошую шерсть. Получены плодовитые гибриды от скрещивания местного (индийского) крупного рогатого скота с зебу. При скрещивании белуги и стерляди получен плодовитый гибрид — бестер, хорька и норки — хонорик, продуктивен гибрид между карпом и карасём.
В природе встречаются гибриды зебры и лошади (зеброид), бизона и зубра (зубробизон), тетерева и куропатки (межняк), зайца-русака и зайца-беляка (тумак), соболя и лисицы (кидус), а также тигра и льва (лигр).
В качестве примеров межродовых гибридов растений можно назвать гибрид пшеницы и ржи (тритикале), пшенично-пырейный гибрид, гибрид смородины и крыжовника (йошта), гибрид брюквы и кормовой капусты (куузика), гибриды озимой ржи и житняка, травянистого и древовидного томатов и др.
Гетерозис — явление повышенной жизнеспособности, урожайности, плодовитости гибридов первого поколения, превышающих по этим параметрам обоих родителей.
Уже со второго поколения гетерозисный эффект угасает. По-видимому, это происходит вследствие снижения числа гетерозиготных организмов и повышения доли гомозигот.
Классическими примерами проявления гетерозиса являются мул (гибрид кобылы и осла) и лошак (гибрид коня и ослицы) (рис. 1,2) . Это сильные, выносливые животные, которые могут использоваться в значительно более трудных условиях, чем родительские формы.


Рис. 1. Мул Рис. 2. Лошак
Продолжительность их жизни значительно выше, чем у родительских видов.
Лошак меньше мула ростом и строптив, поэтому менее удобен для использования в хозяйственной деятельности человека.
Гетерозис широко применяют в промышленном птицеводстве, например — бройлерные цыплята, отличающиеся очень быстрым ростом. Цыплёнок-бройлер — финальный гибрид, полученный в результате скрещивания нескольких линий разных пород кур (мясных родительских форм), проверенных на сочетаемость. Первоначально для такого скрещивания использовали породы корниш (в качестве отцовской формы) и белый плимутрок (в качестве материнской формы).
ИСКУССТВЕННЫЙ МУТАГЕНЕЗ
Искусственный мутагенез чаще всего используется как метод селекции растений. Он основан на применении физических и химических мутагенов для получения форм растений с выраженными мутациями. Такие формы в дальнейшем используются для гибридизации или отбора.
В селекции растений широко используется полиплоидия.
Полиплоидия — увеличение числа наборов хромосом в клетках организма, кратное гаплоидному (одинарному) числу хромосом; тип геномной мутации.
Половые клетки большинства организмов гаплоидны (содержат один набор хромосом — n), соматические — диплоидны (2n). Организмы, клетки которых содержат более двух наборов хромосом, называются полиплоидами, три набора — триплоидами (3n), четыре — тетраплоидами (4n) и т. д. Наиболее часто встречаются организмы с числом хромосомных наборов, кратным двум, — тетраплоиды, гексаплоиды (6n) и т. д.
Полиплоиды с нечётным числом наборов хромосом (триплоиды, пентаплоиды и т. д.) обычно не дают потомства (стерильны), т. к. образуемые ими половые клетки содержат неполный набор хромосом — не кратный гаплоидному.
ПОЯВЛЕНИЕ ПОЛИПЛОИДИИ
Благодаря полиплоидии выведены высокоурожайные полиплоидные сорта сахарной свеклы, хлопчатника, гречихи и др. Полиплоидные растения часто более жизнеспособны и плодовиты, чем нормальные диплоиды. О их большей устойчивости к холоду свидетельствует увеличение числа видов-полиплоидов в высоких широтах и в высокогорьях.
Поскольку полиплоидные формы часто обладают ценными хозяйственными признаками, искусственную полиплоидизацию применяют в растениеводстве для получения исходного селекционного материала.
Получение полиплоидов в эксперименте тесно связано с искусственным мутагенезом. С этой целью используют специальные мутагены (например, алкалоид колхицин), нарушающие расхождение хромосом в митозе и мейозе.
Получены урожайные полиплоиды ржи, гречихи, сахарной свёклы и других культурных растений; стерильные триплоиды арбуза, винограда, банана популярны благодаря бессемянным плодам.
Применение отдалённой гибридизации в сочетании с искусственной полиплоидизацией позволило отечественным учёным получить плодовитые полиплоидные гибриды растений (Г. Д. Карпеченко, гибрид-тетраплоид редьки и капусты) и животных (Б. Л. Астауров, гибрид-тетраплоид тутового шелкопряда).
ШЕЛКОПРЯДЫ АСТАУРОВА
биотехнология
Биотехнология — наука, изучающая возможность модификации биологических организмов для обеспечения потребностей человека.
Применение биотехнологии (рис. 3):
- производство лекарств, удобрений, средств биологической защиты растений;
- биологическая очистка сточных вод;
- восстановление ценных металлов из морской воды;
- коррекция и исправление генетических патологий.

Рис. 3. Возможности биотехнологии
Например, включение в геном кишечной палочки гена, ответственного за образование у человека инсулина, позволило наладить промышленное получение этого гормона (рис. 4).

Рис. 4. Биотехнология получения инсулина
В биотехнологии успешно применяются методы генной и клеточной инженерии.
ГЕННАЯ И КЛЕТОЧНАЯ ИНЖЕНЕРИЯ
Генная инженерия — искусственное, целенаправленное изменение генотипа микроорганизмов с целью получения культур с заранее заданными свойствами.
Исследования в области генной инженерии распространяются не только на микроорганизмы, но и на человека. Они особенно актуальны при лечении болезней, связанных с нарушениями в иммунной системе, в системе свертывания крови, в онкологии.
Основной метод генной инженерии: выделение необходимых генов, их клонирование и введение в новую генетическую среду. Например, введение определённых генов с помощью плазмиды в организм бактерии для синтеза ею определённого белка (рис. 5).

Рис. 5. Применение генной инженерии
Основные этапы решения генно-инженерной задачи следующие:
- Получение изолированного гена.
- Введение гена в вектор (плазмиду) для переноса в организм.
- Перенос вектора с геном (рекомбинантной плазмиды) в модифицируемый организм.
- Преобразование клеток организма.
- Отбор генетически модифицированных организмов и устранение тех, которые не были успешно модифицированы.
Клеточная инженерия — это направление в науке и селекционной практике, которое изучает методы гибридизации соматических клеток, принадлежащих разным видам, возможности клонирования тканей или целых организмов из отдельных клеток.
Включает культивирование и клонирование клеток на специально подобранных средах, гибридизацию клеток, пересадку клеточных ядер и другие микрохирургические операции по «разборке» и «сборке» (реконструкции) жизнеспособных клеток из отдельных фрагментов.
На данный момент удалось получить гибриды между клетками животных, далёких по систематическому положению, например мыши и курицы. Соматические гибриды нашли широкое применение как в научных исследованиях, так и в биотехнологии.
Гибридные клетки, полученные от клеток человека и мыши и человека и китайского хомячка, участвовали в расшифровке генома человека.
Гибриды между опухолевыми клетками и лимфоцитами обладают свойствами обеих родительских клеточных линий: они неограниченно делятся и могут вырабатывать определённые антитела. Такие антитела применяют в лечебных и диагностических целях в медицине.
В эмбриологии для изучения процессов дифференцировки клеток и тканей в ходе онтогенеза используют организмы — химеры , состоящие из клеток с разными генотипами . Их создают путём соединения клеток разных зародышей на ранних этапах их развития.
Клонирование животных — ещё один метод клеточной инженерии: ядро соматической клетки пересаживают в лишённую ядра яйцеклетку с последующим выращиванием зародыша во взрослый организм.
Преимущество клеточной инженерии в том, что она позволяет экспериментировать с клетками, а не с целыми организмами.
Методы клеточной инженерии часто применяют в сочетании с генной инженерией.
работы Н. И. Вавилова
Николай Иванович Вавилов — российский генетик, растениевод, географ.
- Н. И. Вавилов организовал 180 экспедиций (20−30 гг. ХХ века) по самым труднодоступным и зачастую опасным районам земного шара с целью изучения многообразия и географического распространения культурных растений.
- Им была собрана уникальная, самая крупная в мире коллекция культурных растений (к 1940 г. коллекция включала 300 000 образцов), которые ежегодно размножаются в коллекциях Всероссийского института растениеводства имени Н. И. Вавилова (ВИР) и широко используются селекционерами как исходный материал для создания новых сортов зерновых, плодовых, овощных, технических, лекарственных и других культур.
- Создал учение об иммунитете растений.
УЧЕНИЕ ОБ ИММУНИТЕТЕ РАСТЕНИЙ
ЦЕНТРЫ ПРОИСХОЖДЕНИЯ КУЛЬТУРНЫХ РАСТЕНИЙ
Центры происхождения культурных растений — географические области, являющиеся родиной дикорастущих предков культурных растений.
Центры происхождения важнейших культурных растений связаны с древними очагами цивилизации и местом первичного возделывания и селекции растений. Подобные очаги одомашнивания (центры доместикации) выявлены и у домашних животных.
Было выделено восемь центров происхождения культурных растений (рис. 6):
1. Средиземноморский (спаржа, маслины, капуста, лук, клевер, мак, свекла, морковь).
2. Переднеазитский (инжир, миндаль, виноград, гранат, люцерна, рожь, дыня, роза).
3. Среднеазиатский (нут, абрикос, горох, груша, чечевица, лен, чеснок, мягкая пшеница).
4. Индо-Малайский (цитрусовые, хлебное дерево, огурец, манго, черный перец, кокосовая пальма, банан, баклажан).
5. Китайский (просо, редька, вишня, яблоко, гречиха, слива, соя, хурма).
6. Центральноамериканский (тыква, фасоль, какао, авокадо, махорка, кукуруза, батат, хлопчатник).
7. Южноамериканский (табак, ананас, томат, картофель).
8. Абиссинский центр (банан, кофе, сорго, твердая пшеница).
В поздних работах Н. И. Вавилова Переднеазиатский и Среднеазиатский центры объединяются в Юго-западноазиатский центр.

Рис. 6. Центры происхождения культурных растений
В настоящее время выделяют 12 первичных центров происхождения культурных растений.
Источник



