
- Глюкагон — функции, синтез, действие
- Содержание
- Глюкагон [ править | править код ]
- Историческая справка [ править | править код ]
- Химические свойства [ править | править код ]
- Регуляция секреции [ править | править код ]
- Метаболизм [ править | править код ]
- Механизм действия [ править | править код ]
- Применение [ править | править код ]
- Способ транспорта глюкагона по кровеносному руслу
- Видео физиология гормонов поджелудочной железы и регуляции уровня глюкозы — профессор, д.м.н. П.Е. Умрюхин
- Способ транспорта глюкагона по кровеносному руслу
Глюкагон — функции, синтез, действие
Содержание
Глюкагон [ править | править код ]
Историческая справка [ править | править код ]
Еще до открытия инсулина в островках поджелудочной железы были обнаружены разные группы клеток. Сам глюкагон был открыт Мерлином и Кимбаллом в 1923 г., менее чем через 2 года после инсулина. Однако если открытие инсулина вызвало ажиотаж, то глюкагоном мало кто заинтересовался. Только по прошествии более 40 лет стало ясно, какую важную физиологическую роль играет этот гормон в регуляции обмена глюкозы и кетоновых тел, но его роль как лекарственного средства и на сегодняшний день невелика. Глюкагон используют лишь для быстрого купирования гипогликемии, а также в лучевой диагностике в качестве препарата, подавляющего моторику кишечника.
Химические свойства [ править | править код ]
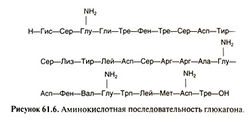
Глюкагон представляет собой одноцепочечный полипептид, состоящий из 29 аминокислотных остатков (рис. 61.6). Между глюкагоном и другими полипептидными гормонами, в том числе секретином, ВИП и гастроингибирующим пептидом, имеется значительная гомология. Аминокислотная последовательность глюкагона у млекопитающих высококонсервативна; она одинакова у человека, коровы, свиньи и крысы.
Глюкагон образуется из препроглюкагона — пептида-предшественника, состоящего из 180 аминокислот и пяти доменов которые подвергаются раздельному процессингу (Bell et al., 1983). За N-концевым сигнальным пептидом в молекуле препроглюкагона следует глицентиноподобный панкреатический пептид затем идут аминокислотные последовательности глюкагона и глюкагоноподобных пептидов типов 1 и 2. Процессинг препроглюкагона осуществляется в несколько этапов и зависит от ткани, в которой он происходит. В результате из одного и того же препрогормона в а-клетках островков поджелудочной железы и в нейроэндокринных клетках кишечника (L-клетках) образуются разные пептиды (Mojsov et al., 1986). Глицентин, важнейший промежуточный продукт процессинга, состоит из N-концевого глицентиноподобного панкреатического пептида и С-концево-го глюкагона, разделенных между собой двумя остатками аргинина. Оксинтомодулин состоит из глюкагона и С-концевого гексапептида, тоже разделенных между собой двумя остатками аргинина.
Физиологическая роль пептидов — предшественников глюкагона не ясна, однако сложная регуляция процессинга препроглюкагона предполагает, что всем им должны быть присущи особые функции. В секреторных гранулах а-клеток островков поджелудочной железы различимы центральное ядро из глюкагона и периферический ободок из глицентина. В L-клетках кишечника секреторные гранулы содержат только глицентин; по-видимому, эти клетки лишены фермента, который превращает глицентин в глюкагон. Оксинтомодулин связывается с рецепторами глюкагона на гепатоцитах и стимулирует аденилатциклазу; активность этого пептида составляет 10—20% активности глюкагона. Глюкагоноподобный пептид типа 1 — чрезвычайно сильный стимулятор секреции инсулина, но он не оказывает почти никакого действия на гепатоциты. Глицентин, оксинтомодулин и глюкагоноподобные пептиды обнаруживаются преимущественно в кишечнике. Их секреция продолжается и после панкреатэктомии.
Регуляция секреции [ править | править код ]
Секреция глюкагона регулируется глюкозой, поступающей с пищей, инсулином, аминокислотами и жирными кислотами. Глюкоза — мощный ингибитор секреции глюкагона. При приеме внутрь она оказывает гораздо более сильное действие на секрецию глюкагона, чем при в/в введении (как, впрочем, и на секрецию инсулина). Вероятно, эффект глюкозы опосредуется какими-то пищеварительными гормонами. Он утрачивается при нелеченном или декомпенсированном инсулинозависимом сахарном диабете, отсутствует и в культуре a-кпеток. Следовательно, действие глюкозы на а-клетки, по крайней мере отчасти, зависит от стимуляции ею секреции инсулина. Соматостатин, свободные жирные кислоты и кетоновые тела тоже тормозят секрецию глюкагона.
Большинство аминокислот стимулируют секрецию и глюкагона, и инсулина. Это объясняет, почему после приема чисто белковой пищи у человека не возникает опосредованная инсулином гипогликемия. Подобно глюкозе, аминокислоты более эффективны при приеме внутрь, чем при в/в введении. Следовательно, их эффект также может быть отчасти опосредован пищеварительными гормонами. Кроме того, секреция глюкагона находится под контролем вегетативной нервной системы. Раздражение симпатических нервных волокон, иннервирующих островки поджелудочной железы, а также введение адреностимуляторов и симпатомиметиков усиливают секрецию этого гормона. Ацетилхолин обладает аналогичным действием. Глюкагон при сахарном диабете. У больных с декомпенсированным сахарным диабетом концентрация глюкагона в плазме повышена. Благодаря своей способности усиливать глюконеогенез и гликогенолиз глюкагон усугубляет гипергликемию. Однако нарушения секреции глюкагона при сахарном диабете, по-ви-димому, носят вторичный характер и исчезают при нормализации уровня глюкозы в крови (Unger, 1985). Роль гиперглюкагонемии при сахарном диабете прояснили эксперименты с введением соматостатина (Gerich et al., 1975). Соматостатин, хотя и не нормализует полностью метаболизм глюкозы, существенно замедляет скорость развития гипергликемии и кетонемии у больных инсулинозависимым сахарным диабетом после внезапной отмены инсулина. У здоровых людей в ответ на гипогликемию секреция глюкагона увеличивается, а при инсулинозависимом сахарном диабете этот важный защитный механизм утрачивается еще в самом начале болезни.
Метаболизм [ править | править код ]
Глюкагон быстро разрушается в печени, почках и плазме, а также в тканях-мишенях (Peterson etal., 1982). EroT1/2 в плазме составляет всего 3—6 мин. Отщепление протеазами N-концевого остатка гистидина приводит к потере глюкагоном биологической активности.
Механизм действия [ править | править код ]
Глюкагон связывается с рецептором на мембране клеток-мишеней; этот рецептор представляет собой гликопротеид с молекулярной массой 60 ООО (Sheetz and Tager, 1988). Структура рецептора полностью не расшифрована, однако известно, что он сопряжен с Gj-белком, который активирует аденилатциклазу (гл. 2). Главный эффект глюкагона на гепатоциты опосредуется цАМФ. Модификация N-концевого участка молекулы глюкагона превращает его в частичный агонист: сродство к рецептору в той или иной степени сохраняется, а способность активировать аденилатциклазу в значительной степени утрачивается (Unson et al., 1989). Так ведут себя, в частности, [Фен’]-глюкагон и дез-Гис’-[Глу9]-глюкагонамид.
Посредством цАМФ-зависимого фосфорилирования глюкагон активирует фосфорилазу — фермент, который катализирует лимитирующую реакцию гликогенолиза. Одновременно происходит фосфорилирование гликогенсинтетазы, и ее активность снижается. В результате гликогенолиз усиливается, а гликогенез тормозится. цАМФ стимулирует также транскрипцию гена фосфоенолпируваткарбоксикиназы — фермента, который катализирует лимитирующую реакцию глюконеогенеза (Granner et al., 1986). В норме инсулин вызывает противоположные эффекты, и когда концентрации обоих гормонов максимальны, действие инсулина преобладает.
цАМФ опосредует фосфорилирование еще одного бифункционального фермента — 6-фосфофрукто-2-киназы/фрукто-зо-2,6-дифосфатазы (Pilkis et al., 1981; Foster, 1984). От этого фермента зависит внутриклеточная концентрация фруктозо-2,6-дифосфата, который, в свою очередь, регулирует глюконеогенез и гликогенолиз. Когда концентрация глюкагона высока, а инсулина низка, 6-фосфофрукто-2-киназа/фрукго-зо-2,6-дифосфатаза фосфорилируется и работает как фосфата-за, снижая содержание фруктозо-2,6-дифосфата в печени. Когда концентрация инсулина высока, а глюкагона низка, фермент дефосфорилируется и работает как киназа, увеличивая содержание фрукгозо-2,6-дифосфата. Фруктозо-2,6-дифосфат является аллостерическим активатором фосфофруктокиназы — фермента, который катализирует лимитирующую реакцию гликолиза. Таким образом, когда концентрация глюкагона высока, гликолиз тормозится, а глюконеогенез усиливается. Это приводит к увеличению уровня малонил-КоА, ускорению окисления жирных кислот и кетогенезу. Напротив, когда высока концентрация инсулина, гликолиз усиливается, а глюконеогенез и ке-тогенез подавляются (Foster, 1984).
Глюкагон, особенно в высоких концентрациях, действует не только на печень, но и на другие ткани. В жировой ткани он активирует аденилатциклазу и усиливает липолиз, в миокарде — увеличивает силу сердечных сокращений. Глюкагон расслабляет гладкие мышцы ЖКТ; такой же эффект оказывают аналоги гормона, не активирующие аденилатциклазу. В некоторых тканях (в том числе в печени) имеется еще один тип рецепторов глюкагона; связывание гормона с ними приводит к образованию ИФ3, ДАГ и увеличению внутриклеточной концентрации кальция (Murphy et al., 1987). Роль этого рецептора глюкагона в регуляции метаболизма остается неизвестной.
Применение [ править | править код ]
Глюкагон используют для лечения тяжелых эпизодов гипогликемии, обычно у больных сахарным диабетом, когда невозможно организовать в/в инфузию глюкозы. Кроме того, глюкагон применяют в лучевой диагностике как средство, подавляющее моторику ЖКТ.
Глюкагон, применяемый в лечебных целях, получают из бычьей и свиной поджелудочных желез. Аминокислотные последовательности человеческого, бычьего и свиного глюкагона идентичны. При гипогликемии 1 мг глюкагона вводят в/в, в/м или п/к. В экстренных ситуациях первые два пути введения предпочтительны. Улучшение наступает в течение 10 мин, что позволяет свести к минимуму риск поражения ЦНС. Гипергликемизирующий эффект глюкагона кратковременен и может вообще не проявиться, если запасы гликогена в печени истощены. После улучшения, наступившего под действием глюкагона, больному вводят глюкозу либо заставляют его что-нибудь съесть, чтобы предотвратить рецидив гипогликемии. Самые частые побочные эффекты глюкагона — тошнота и рвота.
Глюкагон назначают перед рентгеноконтрастными исследованиями верхних и нижних отделов ЖКТ, перед ретроградной идеографией (Monsein et al., 1986) и перед MPT (Goldberg and Thoeni, 1989) для того, чтобы расслабить гладкие мышцы желудка и кишечника. Его используют также для снятия спазма при остром дивертикулите, патологии желчных путей и сфинктера Одди, как вспомогательное средство при удалении желчных камней с помощью петли Дормиа, а также при обструкции пищевода и инвагинации кишечника (Friedland, 1983; Mortens-sonetal., 1984; Kadir and Gadacz, 1987). Введение глюкагона облегчает дифференциальную диагностику механической и паренхиматозной желтухи (Berstock et al., 1982).
Глюкагон стимулирует выброс катехоламинов клетками феохромоцитомы и используется как экспериментальное диагностическое средство при этой опухоли. Кроме того, глюкагоном пытались лечить шок, используя его инотропное действие на сердце. Препарат оказался полезным для тех больных, которые принимали β-адреноблокаторы, потому что β-адреностимуляторы у них неэффективны.
Источник
Способ транспорта глюкагона по кровеносному руслу
Глюкагон — гормон, секретируемый альфа-клетками островков Лангерганса при снижении уровня глюкозы в крови. Он выполняет функции, диаметрально противоположные функциям инсулина. Наиболее важная из них — увеличение уровня глюкозы в крови, т.е. действие, прямо противоположное влиянию инсулина.
Подобно инсулину глюкагон является крупным полипептидом. Его молекулярная масса — 3485, он состоит из цепочки из 29 аминокислот. Введение чистого глюкагона животным сопровождается выраженной гипергликемией. Инъекция 1 мкг/кг глюкагона способна увеличить концентрацию глюкозы в крови на 20 мг/дл крови (увеличение на 25%) в течение 20 мин. По этой причине глюкагон также называют гипергликемическим гормоном.
а) Влияние на метаболизм глюкозы. Главные влияния глюкагона на метаболизм глюкозы заключаются в:
(1) расщеплении гликогена печени (гликогенолиз);
(2) увеличении глюконеогенеза в печени.
Оба этих эффекта приводят к существенному увеличению глюкозы в других органах.
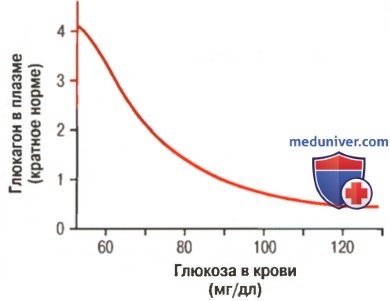 Приблизительная концентрация глюкагона при различных уровнях глюкозы в крови
Приблизительная концентрация глюкагона при различных уровнях глюкозы в крови
б) Глюкагон вызывает гликогенолиз и увеличивает концентрацию глюкозы в крови. Наиболее мощный эффект глюкагона опосредован его способностью вызывать гликогенолиз в печени, увеличивая таким образом концентрацию глюкозы в крови в течение нескольких минут.
Это достигается комплексом последовательных событий.
1. Глюкагон активирует аденилатциклазу в мембране гепатоцитов.
2. Это приводит к образованию циклического аденозинмонофосфата.
3. цАМФ активирует белок-регулятор протеинкиназы.
4. Белок-регулятор протеинкиназы активирует протеинкиназу.
5. Это, в свою очередь, приводит к активации киназы фосфорилазы В.
6. Киназа фосфорилазы В превращает фосфорилазу в фосфорилазу А.
7. Это обеспечивает расщепление гликогена и появление глюкозо-1-фосфата.
8. Глюкозо-1-фосфат дефосфорилируется, затем глюкоза выделяется из клеток печени.
Эта последовательность событий чрезвычайно важна по нескольким причинам: во-первых, это один из наиболее тщательно изученных процессов, где циклический аденозинмонофосфат функционирует как вторичный посредник; во-вторых, это демонстрация каскадной системы, в которой каждый последующий продукт производится в большем количестве, чем предшествующий, поэтому данный процесс дает представление о возможностях механизма биологического усиления. Такой тип механизма усиления используется повсеместно в организме для управления многими, если не большинством клеточных метаболических систем, т.к. часто усиливающий эффект этого механизма возрастает в миллион раз. Это объясняет способность всего нескольких микрограмм глюкагона буквально за несколько минут обеспечить увеличение концентрации глюкозы в крови в 2 раза по сравнению с исходным уровнем.
Инъекция глюкагона всего через 4 ч может вызвать столь интенсивный гликогенолиз, что это приведет к опустошению депо гликогена в печени.
Видео физиология гормонов поджелудочной железы и регуляции уровня глюкозы — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
Способ транспорта глюкагона по кровеносному руслу
Глюкагон является мощным контринсулярным гормоном и его эффекты реализуются в тканях через систему вторичного посредника аденилатциклаза—цАМФ. В отличие от инсулина, глюкагон повышает уровень сахара крови, в связи с чем его называют гипергликемическим гормоном.
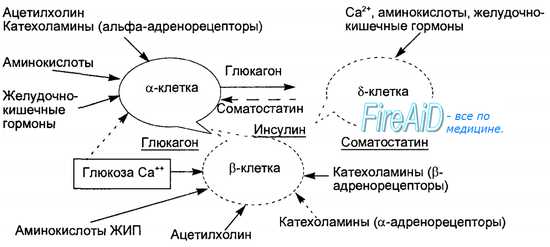 Рис. 6.21. Функциональная организация островков Лангерганса как «мини-органа». Сплошные стрелки — стимуляция, пунктирные — подавление гормональных секретов. Ведущий регулятор — глюкоза — при участии кальция стимулирует секрецию инсулина р-клетками и, напротив, тормозит секрецию глюкагона альфа-клетками. Всасывающиеся в желудке и кишечнике аминокислоты являются стимуляторами функции всех клеточных элементов «мини-органа». Ведущий «внутриорганный» ингибитор секреции инсулина и глюкагона — соматостатин, активация его секреции происходит под влиянием всасывающихся в кишечнике аминокислот и гастроинтестинальных гормонов при участии ионов Са2+. Глюкагон является стимулятором секреции как соматостатина, так и инсулина.
Рис. 6.21. Функциональная организация островков Лангерганса как «мини-органа». Сплошные стрелки — стимуляция, пунктирные — подавление гормональных секретов. Ведущий регулятор — глюкоза — при участии кальция стимулирует секрецию инсулина р-клетками и, напротив, тормозит секрецию глюкагона альфа-клетками. Всасывающиеся в желудке и кишечнике аминокислоты являются стимуляторами функции всех клеточных элементов «мини-органа». Ведущий «внутриорганный» ингибитор секреции инсулина и глюкагона — соматостатин, активация его секреции происходит под влиянием всасывающихся в кишечнике аминокислот и гастроинтестинальных гормонов при участии ионов Са2+. Глюкагон является стимулятором секреции как соматостатина, так и инсулина.
Основные эффекты глюкагона проявляются в следующих сдвигах метаболизма в организме:
• активация гликогенолиза в печени и мышцах;
• активация глюконеогенеза;
• активация липолиза и подавление синтеза жира в адипоцитах;
• повышение синтеза кетоновых тел в печени и угнетение их окисления;
• стимуляция катаболизма белков в клетках тканей, прежде всего печени, и увеличение синтеза в ней мочевины.
Образующиеся в островках Лангерганса гастрин и панкреатический полипептид основную роль играют в регуляции процессов пищеварения, их эффекты и физиологическая роль рассмотрены в соответствующей главе.
Источник



