
- Репродукция вирусов
- Способ репродукции свойственный вирусам
- Паповавирусы и аденовирусы. Репродукция паповавирусов. Репродукция аденовирусов.
- Репродукция вируса
- Содержание:
- Общие закономерности репродукции вируса
- Этапы репродукции вируса
- Адсорбция на мембране клетки
- Проникновение в клетку
- Депротеинизация
- Синтез компонентов вирусов
- Формирование дочерних вирионов
- Выход вирионов
Репродукция вирусов




![]()
![]()
Для вирусов характерен дизъюнктивный (от disjuncus — разобщенный) способ репродукции-размножения. Потомство вируса возникает в результате сборки нуклеиновых кислот и белковых субъединиц, которые синтезируются раздельно клеткой хозяина.
Проникновение вируса в клетку и воспроизведение себе подобных проходит в несколько фаз:
1.проникновение в клетку хозяина,
2.синтез ферментов, необходимых для репликации вирусных нуклеиновых кислот,
3.синтез вирусных частей,
4.сборка и композиция зрелых вирионов,
5.выход зрелых вирионов из клетки.
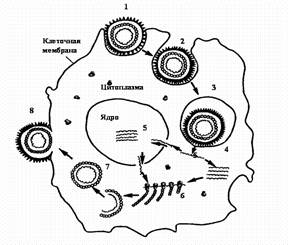
Стадии репродукции вирусов.
1 — адсорбция вириона на клетке; 2 — проникновение вириона в клетку путем виропексиса;
3 — вирус внутри вакуоли клетки; 4 — `раздевание вириона вируса; 5 — репликация вирусной нуклеиновой кислоты; 6 — синтез вирусных белков на рибосомах клетки; 7 — формирование вириона; 8 — выход вириона из клетки путем почкования.
Фаза I — адсорбция вириона на поверхности клетки.
Протекает в две стадии: первая — неспецифическая, когда вирус удерживается на поверхности клетки при помощи электростатических сил, т. е. благодаря возникновению противоположных зарядов между отдельными участками мембраны клеток и вируса. Эта фаза взаимодействия вируса с клеткой обратима, на нее оказывают влияние такие факторы, как рН и солевой состав среды.
Вторая стадия — специфическая, когда взаимодействуют специфические рецепторы вируса и рецепторы клетки, комплементарные друг другу. По химической природе рецепторы клетки могут быть мукопротеидами (или мукополисахаридами) и липопротеидами. Разные вирусы фиксируются на разных рецепторах: вирусы гриппа, парагриппа, аденовирусы — на мукопротеидах, а вирусы клещевого энцефалита, полиомиелита — на липопротеидах.
Фаза II — проникновение вируса в клетку. Электроноскопические наблюдения за процессом проникновения вирусов в чувствительные к ним клетки показали, что оно осуществляется посредством механизма, напоминающего пиноцитоз, или, как чаще называют, виропексис. В месте адсорбции вируса клеточная стенка втягивается внутрь клетки, образуется вакуоль, в которой оказывается вирион. Параллельно клеточные ферменты (липазы и протеазы) вызывают депротеинизацию вириона — растворение белковой оболочки и освобождение нуклеиновой кислоты.
Фаза III — скрытый период (период эклипса — исчезновения). В этот период в клетке невозможно определить наличие инфекционного вируса ни химическими, ни электронно-микроскопическими, ни серологическими методами. О сущности этого явления и его механизмов пока известно мало. Предполагается, что в скрытой фазе нуклеиновая кислота вируса проникает в хромосомы клетки и вступает с ними в сложные генетические взаимоотношения.
Фаза IV — синтез компонентов вириона. В этой фазе вирус и клетка представляют единое целое, вирусная нуклеиновая кислота выполняет генетическую функцию, индуцирует образование ранних белков и изменяет функцию рибосом. Ранние белки подразделяются на:
а) белки-ингибиторы (репрессоры), подавляющие метаболизм клеток
б) белки-ферменты (полимеразы), обеспечивающие синтез вирусных нуклеиновых кислот.
Синтез нуклеиновых кислот и белков протекает неодновременно и в разных структурных частях клетки. У вирусов, содержащих ДНК или РНК, эти процессы имеют некоторые различия и особенности.
Фаза V — формирование зрелых вирионов. Процесс «сборки» вируса осуществляется в результате соединения компонентов вирусной частицы. У сложных вирусов в этом процессе принимают участие клеточные структуры и происходит включение в вирусную частицу липидпых, углеводных, белковых компонентов клетки хозяина.
Процесс формирования вирионов начинается спустя определенное время после того, как начал осуществляться синтез составляющих их компонентов. Продолжительность этого периода довольно вариабельна и предопределяется природой вируса — для РНК-содержащих обычно короче, чем для ДНК-вирусов. Например, продукция полных вирусных частиц осповакцины начинается приблизительно спустя 5—6 ч после инфицирования клеток и продолжается в течение последующих 7—8 ч, т. е. после того как синтез вирусной ДНК уже завершен.

Между нуклеиновой кислотой и соответствующим белковыми субъединицами образуются очень прочные связи, о чем свидетельствуют трудности отделения белка от вирусной нуклеиновой кислоты. Большую прочность вирусной частице придают входящие в ее состав углеводы и особенно липиды.
Формирование вирионов, так же как и синтез компонентов вируса, происходит в разных местах клетки, при участии различных клеточных структур. После завершения процесса формирования образуется зрелая дочерняя вирусная частица, обладающая всеми свойствами родительского вириона. Но иногда наблюдается образование так называемых неполных вирусов, которые состоят или только из нуклеиновой кислоты, или из белка, или из вирусных частиц, формирование которых остановилось в какой-то промежуточной стадии.
Фаза VI — выход зрелых вирионов из клетки. Существуют два основных механизма выхода зрелых вирионов из клетки:
1) выход вириона с помощью почкования. В этом случае наружная оболочка вириона происходит из клеточной мембраны, она содержит как материал клетки хозяина, так и вирусный материал;
2) выход зрелых вирионов из клетки через бреши в мембране. Эти вирусы не имеют наружной оболочки. При таком механизме выхода вирусов клетка, как правило, погибает и в среде появляется большое количество вирусных частиц.
Причиной гибели зараженной клетки могут быть три механизма:
1.работа вируса, «истощающая» клетку;
2.защитная реакция клетки, запускающая генетическую программу ее гибели (апоптоз);
3. иммунная система организма, уничтожающая зараженную клетку.
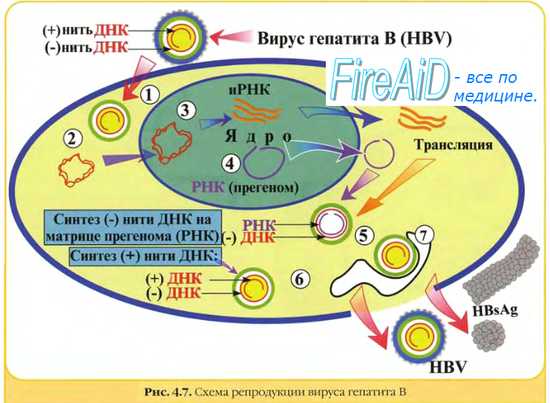
Кроме продуктивного типа взаимодействия вируса и клетки возможно интегративное сосуществование или вирогения. Вирогения характеризуется интеграцией (встраиванием) нуклеиновой кислоты вируса в геном клетки, а также репликацией и функционированием вирусного генома как составной части генома клетки. Для интеграции с клеточным геномом необходимо возникновение кольцевой формы двунитевой ДНК вируса. Встроенная в состав хромосомы клетки вирусная ДНК называется провирусом. Провирус реплицируется в составе хромосомы и переходит в геном дочерних клеток, т.е. состояние вирогении наследуется. Под влиянием некоторых физических или химических факторов провирус может переходить в автономное состояние с развитием продуктивного типа взаимодействия с клеткой. Дополнительная генетическая информация провируса при вирогении сообщает клетке новые свойства, что может быть причиной развития опухолей, аутоиммунных и хронических заболеваний. На способности вирусов к интеграции с геномом клетки основаны персистенция (от лат. persisto — постоянно пребывать, оставаться) вирусов в организме и развитие персистентных вирусных инфекций. Например, вирус гепатита В способен вызывать персистирующие поражения с развитием хронического гепатита и часто опухолей печени.
Источник
Способ репродукции свойственный вирусам
Вирусы, лишённые суперкапсида (например, аденовирусы) проникают в клетки путём виропексиса, а имеющие таковой (покс- и герпесвирусы) — за счёт слияния суперкапсида с клеточной мембраной. Репродуктивный цикл ДНК-содержащих вирусов включает раннюю и позднюю стадии (рис. 5-4). У крупных ДНК-вирусов имеется явное несоответствие между кодирующе ёмкостью генома и молекулярной массой вирусиндуцированных белков и белков, входящих состав вирионов. Например, у герпесвирусов лишь 15% ДНК кодирует все белки вирионов и их предшественников. Возможно, значительная часть генома содержит гены, кодирующие синтез ферментов и регуляторных белков. Папова-, адено- и герпесвирусы репродуцируются относительно однотипно, в то время как репродукция поксвирусов имеет некоторые особенности.
Ранняя стадия репродукции. Вирусная ДНК проникает в ядро клетки, где транскрибируется клеточной ДНК-зависимой РНК-полимеразой. При этом считываетеся, а затем транслируется часть вирусного генома («paнние гены»). В результате синтезируются «ранние белки» (регуляторные и матричные белки вирусные полимеразы).
Регуляторные белки выполняют различные функции. При заражении клетки они блокируют синтез клеточных РНК, ДНК и белка и одновременно способствуют экспрессии вирусного генома, изменяя специфичность реагирования клеточных полимераз и полирибосом. Они так же запускают репликацию клеточной ДНК, модифицированной встроенными геномами ДНК содержащих вирусов и ретровирусов, то есть репликацию вирусных геномов. Вирусспецифические полимеразы. В репликацию вирусных геномов также вовлечены вирусоспецифические ДНК-полимеразы, участвующие в образовании молекул ДНК дочерних популяций.
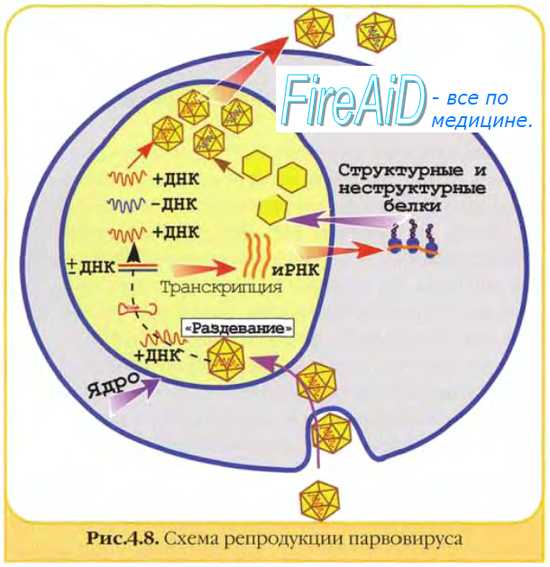
Матричные белки необходимы для репликации нуклеиновых кислот и сборки дочерних популяций. Они образуют электронно-плотные скопления в клетке, известные как тельца включений (например, тельца Гварнери при натуральной оспе).
Поздняя стадия репродукции. На этом этапе происходит синтез нуклеиновых кислот вируса. Не вся вновь синтезированная вирусная ДНК упаковывается в вирионы дочерней популяции. Часть ДНК («поздние гены») используется для синтеза «поздних белков», необходимых для сборки вирионов. Их образование катализируют вирусные и модифицированные клеточные полимеразы.
Паповавирусы и аденовирусы. Репродукция паповавирусов. Репродукция аденовирусов.
Адсорбция, проникновение и депротеинизация аналогичны таковым у РНК-содержащих вирусов, но у папова— и аденовирусов депротеинизация протекает в ядре, а у РНК-вирусов — в цитоплазме.
Ранняя фаза репродукции. Вирусная ДНК («ранние гены») транскрибируется в ядре клетки. На одной из нитей ДНК реализуется транскрипция вирусной «ранней» мРНК. Механизмы транскрипции вирусной ДНК аналогичны считыванию информации с клеточной ДНК. Специфическая мРНК транслируется, начинается синтез ферментов, необходимых для образования дочерних копий ДНК. Синтез клеточной ДНК может временно усиливаться, но затем обязательно подавляется регуляторными белками вируса.
Поздняя фаза репродукции. В течение поздней фазы дочерняя вирусная ДНК продолжает активно транскрибироваться клеточными РНК-полимеразами, в результате чего появляются продукты поздних вирусспецифических синтезов. «Поздняя» мРНК мигрирует в цитоплазму и транслируется на рибосомах. В результате синтезируются капсидные белки дочерней популяции, которые транспортируются в ядро и собираются вокруг молекул дочерней ДНК новых вирусных частиц. Выход полных дочерних популяций сопровождается гибелью клетки.
Источник
Репродукция вируса
Reproduction of the virus
Термин «репродукция» восходит к латинскому слову «production»– производство. Действительно, процесс размножения вирусов в корне отличается от бинарного деления клеток живых организмов. Он больше напоминает процесс, происходящий на заводском конвейере. Процесс репродукции вируса внутри инфицированной клетки называют жизненным циклом вируса. Цикл репродукции у РНК-вирусов продолжается 4–8 часов, а у ДНК-вирусов – 12–24 часа.
Содержание:
Общие закономерности репродукции вируса
К самостоятельному размножению вирусы не способны. Синтез вирусных белков и воспроизведение копий вирусного генома обеспечиваются биосинтетическими процессами клетки-хозяина. Для вирусов характерен дизъюнктивный (разобщенный) тип репродукции. Он осуществляется при взаимодействии вируса с инфицируемой клеткой. В этом случае белковые молекулы и нуклеиновые кислоты образуются отдельно друг от друга. После чего происходит сборка дочерних популяций.
Особенности репродукции вирусов зависят от типа вирусного генома. Однако отмечается существование целого ряда общих закономерностей репродукции вируса:
- Все вирусы, содержащие молекулу РНК, кроме вирусов гриппа и ретровирусов, репродуцируются в цитоплазме клетки. Геномы ретровирусов и вирусов гриппа при репродукции проникают в ядро клетки-хозяина.
- Все вирусы, содержащие молекулу ДНК, кроме вируса оспы репродуцируются в ядре и в цитоплазме клетки. В ядре происходит транскрипция и репликация вирусных нуклеиновых кислот, а в цитоплазме – трансляция вирусных белков и сборка дочерних вирионов. Вирус оспы размножается только в цитоплазме клетки.
- Процесс синтеза нуклеокапсидных белков происходит на свободных полирибосомах (не связанных с мембраной). Процесс синтеза суперкапсидных белков проходит на рибосомах, ассоциированных с мембранами.
- Белки отдельных вирусов после образования подвергаются протеолитическому процессингу (разрезанию или расщеплению).
Суперкапсидные белки оболочечных вирусов при транспортировке к клеточной мембране проходят гликозирование(присоединении к полипептиду углеводных остатков).
Этапы репродукции вируса
Репродукция или жизненный цикл вируса делят на несколько шесть последовательных этапов.
- адсорбция на мембране клетки;
- проникновение в клетку;
- депротеинизация;
- синтез компонентов вирусов;
- формирование дочерних вирионов;
- выход вирионов.
Адсорбция на мембране клетки
Адсорбция вириона на мембране клетки идет по пути взаимодействия вирусного белка (антирецептора) с клеточными рецепторами. Для каждого вируса на клеточной мембране существуют специфические рецепторы, с которым он и связывается. По химической природе рецепторы, фиксирующие вирус, могут являться мукопротеиновыми либо липопротеиновыми. Распознавание клеточных рецепторов осуществляют капсидные или суперкапсидные белки вириона.
Антирецепторы вирионов являются прикрепительными белками. Они могут иметь форму шипов, нитей, грибовидных структур.
В самом процессе адсорбции большую роль играют электрические заряды. Вирусы обычно отрицательно заражены, а участки клеточной стенки – положительно.
Процесс адсорбции занимает от пяти до девяноста минут. Количество специфических рецепторов на поверхности одной клетки 10 4 –10 5 .
Проникновение в клетку
Путь проникновения вируса в клетку зависит от наличия оболочки у вириона. Существует два пути:
- путь слиянии вирусной оболочки с клеточной мембраной.
- путь рецептор-опосредованного эндоцитоза.
Путем слияния суперкапсида с клеточной мембраной в клетку проникают оболочечные вирусы. Этот процесс обусловлен наличием специфических белков слияния. При этом наблюдается высвобождение нуклеокпсида в цитоплазму клетки.
Путем рецептор-опосредованного эндоцитоза в клетку проникают безоболочечные вирусы. Первоначально вирион связывается со специфическими рецепторами, расположенными на клеточной поверхности. Затем наблюдается инвагинация (впячивание) клеточной мембраны, образование эндосом (внутриклеточных вакуолей) и их слияние с лизосомами. В заключении вирусный геном в цитоплазме клетки освобождается из эндосомы.
Депротеинизация
Процесс депротеинизации (освобождения вирусной нуклеиновой кислоты – раздевание вируса) осуществляют протеолитические ферменты клетки (протеазы и липазы).
Смысл этого процесса состоит в удалении капсидов (вирусных оболочек). Конечные продукты раздевания вируса – сердцевины, нуклеокапсиды, нуклеиновые кислоты. Некоторые вирусы в качестве конечного продукта представлены нуклеиновыми кислотами, связанными с внутренним вирусным белком. После прохождения этапа депротеинизации выделить вирус из культуры клеток невозможно. Такое положение называют теневой фазой или фазой эклипса (затмения). В этот период вирус перестает существовать в качестве оформленного вириона.
Синтез компонентов вирусов
Синтез компонентов вирусов заключается в репликации вирусных нуклеиновых кислот и синтезе вирусных белков. Под репликацией понимается процесс самовоспроизведения нуклеиновых кислот, генов и хромосом, в основе которого лежит ферментативный синтез ДНК или РНК, проходящий по матричному синтезу.
Место синтеза компонентов дочерних вирионов зависит от типа генома:
реализация генетической информации у ДНК-содержащих вирусов идет по пути:
- ДНК → транскрипция → иРНК → трансляция → белок;
- реализация генетической информации у +РНК-содержащих вирусов идет без этапа транскрипции: +-РНК → трансляция → белок;
- реализация генетической информации у РНК-содержащих вирусов с негативным геномом по схеме: минус-РНК → транскрипция → иРНК → трансляция → белок.
- РНК-содержащиеретровирусы – идут по следующему пути передачи информации:
- РНК → обратная транскрипция → ДНК → транскрипция → иРНК → трансляция → белок.
ДНК-содержащий вирус, проникший в цитоплазму, транспортирует нуклеокапсид к ядру клетки. Вирусная ДНК проникает в структуры клеточного ядра, где и совершается транскрипция или переписывание информации с ДНК на РНК при помощи клеточной полимеразы. Исключение – вирус оспы. Несмотря на то, что он относится к ДНК-содержащим, но его транскрипция протекает в цитоплазме при участии ДНК-полимеразы, проникающего в клетку в составе вириона.
Результатом транскрипции является и то, что на одной из нитей ДНК синтезируется иРНК. В последствии, она перемещается в цитоплазму клетки и запускает процесс трансляции – перевода генетической информации с иРНК на последовательность аминокислот в вирусных белках.
Синтез белков наблюдается в рибосомах клетки-хозяина. Одновременно в ядре клетки протекает репликация (образование) дочерних нуклеиновых кислот на матрице материнской ДНК.
Синтезированные дочерние молекулы ДНК в составе нуклеокапсида путем почкования перемещаются из ядра клетки в цитоплазму. При этом они захватывают фрагмент ядерной мембраны. В цитоплазме процесс репродукции завершается.
Репликация «плюс»-РНК-содержащих вирусов происходит в цитоплазме. В данном случае функцию иРНК выполняет вирусная нуклеиновая кислота. В итоге трансляции на рибосомах формируется белковая молекула, которая разрезается клеточными протазами на структурные или неструктурные вирусные белки. Одновременно образуется полимераза, способствующая образованию «минус»-РНК на матрице родительской «плюс»-РНК. На матрице «минус»-РНК происходит синтез молекул «плюс»-РНК, принимающих участие в биосинтезе белков дочерних вирионов.
Вирусы, содержащие «минус»-РНК отличаются тем, что их геном не способен выполнить функцию иРНК. Поэтому в цитоплазме клетки на матрице «минус»-РНК первоначально синтезируется «плюс»-РНК. Процесс катализируется полимеразой (транскрипазой), находящейся в составе проникшего в клетку вириона.
Синтез «плюс»-РНК приводит к образованию полных и коротких нитей. Короткие «плюс»-РНК–нити принимают участие в синтезе ферментов и бликов для дочерних популяций. Полные нити «плюс»-РНК служат матрицей для синтеза молекул «минус»-РНК дочерних вирионов. Последние транспортируются к клеточной мембране и высвобождаются, захватывая при этом ее фрагмент. Этот фрагмент служит для вирусной частицы суперкапсидом.
РНК-содержащие ретровирусыотличаются тем, что после проникновения в клетку генетическая информация с их РНК переписывается на ДНК, то есть с помощью фермента ревертаза происходит обратная транскрипция. Ревертаза так же попадает в клетку вместе с ретровирусом. Вновь образованная ДНК интегрирует с клеточным геномом и в его составе участвует в образовании иРНК, необходимой для синтеза вирусных белков. Транскрипцию интегрированной ДНК в составе клеточных геномов (переписывание информации с ДНК на РНК) осуществляет клеточная ДНК-зависимая РНК полимераза.
Формирование дочерних вирионов
Сборка дочерних вирионов возможна только при узнавании вирусных нуклеиновых кислот и белков и самопроизвольном их соединении друг с другом. На мембранах эндоплазматического ретикулума взаимодействуют нуклеиновая кислота и белки просто устроенных вирусов, что приводит к образованию упорядоченной структуры.
Сложно устроенные вирусы характеризуются многоступенчатой сборкой. Первоначально их нуклеиновые кислоты взаимодействуют с внутренними белками, образуя нуклеокапсиды. Затем нуклеокапсиды выстраиваются с внутренней стороны клеточной мембраны под участками, модифицированными оболочечными вирусными белками. В результате происходит самосброска вирионов. Количество зрелых вирионов, сформировавшихся в клетке, варьирует от 10 до 10000 и более.
Выход вирионов
Высвобождение дочерних вирионов из клетки может быть осуществлено двумя способами:
- взрывной – путем лизиса клетки;
- путем почкования.
Путь лизиса клетки тесно связан с деструкцией клетки. Он характерен для безоболочечных вирусов, у которых нет суперкапсидной оболочки (суперкапсида).
Выход путем почкования характерен для оболочечных вирусов. При этом клетка-хозяин некоторое время сохраняет жизнеспособность. Содержащие суперкасид вирусы высвобождаются в течении 2–6 часов. Вначале суперкапсидные белки устанавливаются на наружной поверхности мембраны в виде своеобразных шипов, вытесняя клеточные белки. Затем через модифицированную клеточную мембрану проходит нуклеокапсид с образованием суперкапсида.
Источник



