
Биология. 11 класс
§ 16. Клеточный цикл. Репликация ДНК
Клеточный цикл . Все новые клетки образуются путем деления уже существующих. Этот принцип, как уже отмечалось, сформулировал Р. Вирхов еще в середине XIX в. Деление клеток обеспечивает непрерывность существования жизни на нашей планете. Именно благодаря ему осуществляются различные способы бесполого и полового размножения организмов. В основе процессов роста, регенерации и индивидуального развития многоклеточных организмов также лежит деление клеток.
Период существования клетки от момента ее образования из материнской клетки до собственного деления (включая это деление) или гибели называется клеточным циклом.
Продолжительность клеточного цикла у разных организмов и различных клеток в составе одного организма варьирует. Так, у бактерий в благоприятных условиях он длится примерно 20 мин. Короткие клеточные циклы (30—60 мин) характерны для бластомеров рыб и земноводных на этапе дробления, в то время как у млекопитающих промежуток времени между делениями бластомеров может достигать 10 ч и более. У взрослых мышей клетки кишечного эпителия делятся каждые 11—22 ч, а роговицы глаза — приблизительно один раз в трое суток. Для регулярно делящихся клеток многоклеточных организмов длительность клеточного цикла обычно составляет 12—36 ч.
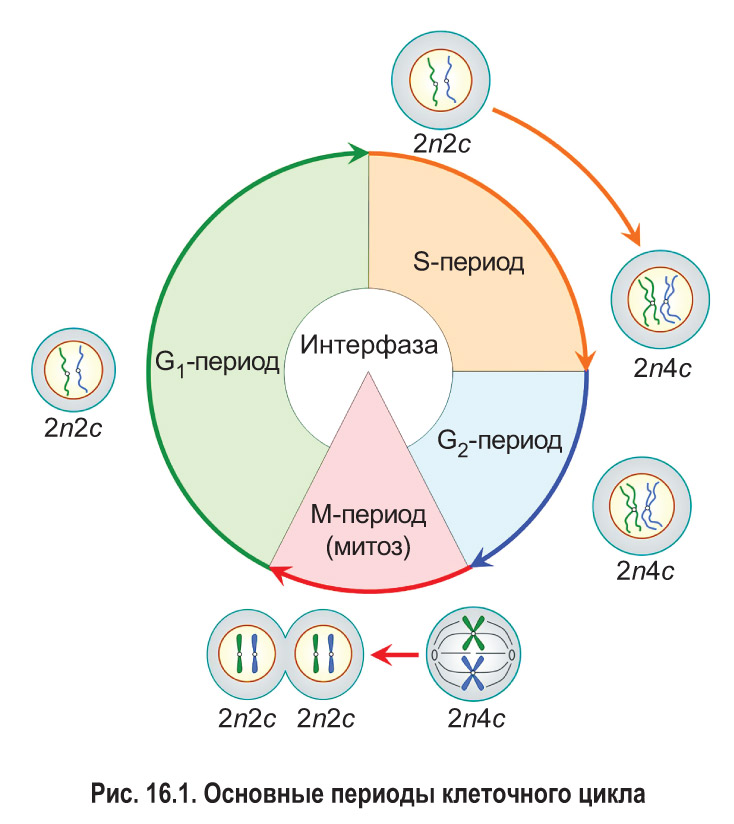 Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Пресинтетический, или G1-пе риод (от англ. gap — промежуток), начинается с момента образования новой клетки в результате деления материнской. Обычно это самый длительный период интерфазы и клеточного цикла в целом. В течение G1-периода молодая клетка интенсивно растет, в ней увеличивается количество органоидов и синтезируются различные соединения, необходимые для протекания процессов жизнедеятельности. В том числе образуются вещества, которые будут нужны для последующего удвоения молекул ДНК.
Вы уже знаете, что набор хромосом обозначают как n: например, 1n — гаплоидный набор, 2n — диплоидный. Набор молекул ДНК в клетках принято записывать с помощью буквы с. Из § 14 вам известно, что каждая хроматида содержит одну молекулу ДНК, т. е. количество молекул ДНК и хроматид в составе хромосом всегда совпадает. Таким образом, записи типа 1с, 2с, 4с отражают содержание в клетках не только молекул ДНК, но и соответствующих хроматид.
В пресинтетическом периоде каждая хромосома состоит из одной хроматиды. Следовательно, в G1-периоде число хромосом (n) и хроматид (с) в клетке одинаковое. Набор хромосом и хроматид диплоидной клетки в G1-периоде клеточного цикла можно выразить записью 2n2c.
В синтетическом, или S-периоде (от англ. synthesis — синтез), происходит удвоение молекул ДНК — репликация, а также удвоение центриолей клеточного центра (в тех клетках, где он имеется). После завершения репликации каждая хромосома состоит уже из двух идентичных сестринских хроматид, которые соединены друг с другом в области центромеры. Количество хроматид в каждой паре гомологичных хромосом становится равным четырем. Таким образом, набор хромосом и хроматид диплоидной клетки в конце S-периода (т. е. после репликации) выражается записью 2n4c.
Постсинтетический, или G2-период, продолжается от окончания синтеза ДНК (репликации) до начала деления клетки. В это время клетка активно запасает энергию и синтезирует белки для предстоящего деления (например, белок тубулин для построения микротрубочек, образующих впоследствии веретено деления). В течение всего G2-периода набор хромосом и хроматид в клетке остается неизменным — 2n4c.
Итак, интерфаза обычно включает три периода: пресинтетический (G1), синтетический (S) и постсинтетический (G2). На протяжении всей интерфазы хромосомы не спирализованы. Они располагаются в ядре клетки в виде хроматина .
После завершения интерфазы начинается деление клетки. Основным способом деления клеток эукариот является митоз, поэтому данный этап клеточного цикла обозначают как М-период. При митозе происходит спирализация хроматина . Это приводит к формированию компактных двухроматидных хромосом. После этого сестринские хроматиды каждой хромосомы отделяются друг от друга и затем попадают в разные дочерние клетки. Значит, дочерние клетки, образовавшиеся в результате митоза и вступающие в новый клеточный цикл, имеют набор 2n2c.
Обобщенная информация об основных периодах клеточного цикла представлена в таблице 16.1.
Таблица 16.1. Основные периоды клеточного цикла
Период
Содержание наследственного материала в диплоидной клетке
Источник
Способ репликации генетического материала
Деление клеток происходит посредством митоза, при этом, во избежание потери генетической информации, вначале удваивается весь ядерный геном в S-фазе клеточного цикла. Продолжительность S-фазы составляет 8 ч. ДНК центромер хромосом реплицируется во время средней стадии митоза, что предшествует процессу сегрегации хромосом.
Репликация митохондриальной и ядерной ДНК происходит в разные фазы клеточного цикла. Несмотря на то что общая последовательность стадий при репликации ядерной ДНК у высших существ (эукариот) и у бактерий (прокариот) одинакова, сам процесс имеет незначительные отличия. Так, у эукариот во время репликации ДНК (ядерная) остаётся в нуклеосомной конфигурации.
Фрагменты ДНК, богатые парами оснований Г—Ц (R-полосы эухроматина в уплотнённом хроматине), экспрессируют гены «домашнего хозяйства», которые функционируют во всех клетках организма. Данные фрагменты реплицируются на ранней стадии S-фазы. Участки гетерохроматина, богатые парами оснований А—Т (G-полосы), экспрессируют небольшое количество генов и реплицируются на поздней стадии S-фазы.
Гены с большим содержанием пар А—Т, кодирующие различные свойства и функционирующие лишь в определённых клетках, входят в состав факультативного гетерохроматина. Их репликация происходит на ранней стадии S-фазы только в тех клетках, в которых они экспрессируются, и на поздних стадиях — в клетках, где экспрессии не происходит.
Область спирали ДНК, которая в начале репликации раскручивается в первую очередь, называют участком начала репликации (репликоном). В этом месте двойную нить расплетает фермент хеликаза, раскрывающий последовательность оснований. Процесс репликации происходит вдоль одной цепи со скоростью примерно 40-50 нуклеотидов в секунду одновременно в обоих направлениях. У высших существ имеется множество репликонов, расположенных на расстоянии 50 000—300 000 п.н. В месте разделения нити ДНК возникают репликационные вздутия, на каждом конце которого формируется репликационная вилка.
Новая ДНК синтезируется при участии ферментов, называемых ДНК-полимеразами, из дезоксирибонуклеотидтрифосфатов (АТФ, ГТФ и др.), которые превращаются в монофосфатные нуклеотиды (АМФ, ГМФ и др.). Отщепление и гидролиз пирофосфатов из трифосфатов обеспечивают процесс энергией и обусловливают его полную необратимость, делая молекулу ДНК достаточно устойчивой.
Все ДНК-полимеразы могут выстраивать новую ДНК только в направлении от 5′- к 3′-концу. Это означает, что ферменты должны двигаться вдоль матричной цепи от 3′- к 5′-концу. В связи с этим репликация может непрерывно происходить от репликона только по одной цепи, называемой опережающей. Из-за расположения Сахаров репликация по второй, отстающей цепи происходит только на коротких отрезках, известных как фрагменты Оказаки.
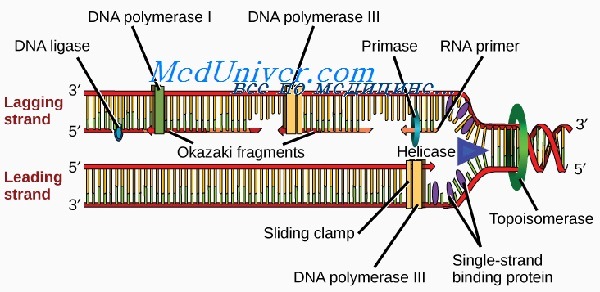
Длина новых фрагментов ДНК, образующихся вдоль отстающей цепи, в среднем составляет 100—200 пар нуклеотидов. Во время синтеза фрагменты Оказаки сшивает между собой фермент ДНК-лигаза. В ожидании репликации стабильность первичной одноцепочечной нуклеотидной последовательности отстающей цепи поддерживается белком, связывающим одноцепочечную ДНК (или спиральдестабилизирующим белком).
Для синтеза опережающей цепи необходим фермент ДНК-полимераза S, а для синтеза отстающей — ДНК-полимераза а. Последняя имеет субъединицу, называемую ДНКпраймазой, которая синтезирует короткую РНК-затравку, играющую роль праймера. Репликация мито-хондриальной ДНК происходит независимо от процессов в ядре. При этом используется ряд других ферментов, один из которых — ДНК-полимераза у.
В геноме присутствует большое количество копий пяти гистонных генов, благодаря чему происходит синтез множества гистонов (особенно во время S-фазы), которые сразу после репликации связываются с новой цепью ДНК.
Следует отметить, что процесс репликации носит название полуконсервативного, так как в состав дочерних молекул ДНК входит одна первичная цепь и одна синтезированная.
Репликация теломер ДНК
Основной проблемой синтеза ДНК на конце отстающей цепи служит то, что ДНК-полимеразе а необходимо прикрепиться выше конца последовательности, которая реплицирована, и работать проксимально в направлении от 5′- к 3′-концу. Для решения этой проблемы нужен ДНК-синтетический фермент теломераза, который продлевает отстающую цепь.
Теломераза — рибонуклеопротеин, содержащий матричную РНК с последовательностью 3′-ААУЦЦЦААУ-5′, которая комплементарна полутора повторам шестиосновной теломерной ДНК (5′-ГГГТТА-3′). Фрагмент последовательности 3′-ААУ РНК-теломеразы связывается с терминальным концом ТТА-5′ матричной отстающей цепи, при этом остальная часть РНК остаётся свободной. Затем к этой матричной РНК присоединяются дезоксирибонуклеотиды, тем самым продлевая повторяющуюся последовательность в ДНК на один сегмент.
После этого теломераза отщепляется и направляется к другому терминальному концу с последовательностью ТТА-5′, и процесс повторяется. Как только возникает достаточно длинный терминальный повтор, ДНК-полимераза а прикрепляется к полученному одноцепочечному фрагменту и достраивает вторую цепь по методу комплементарности в проксимальном 5’—3′-направлении, двигаясь к уже существующему двухцепочечному участку, последующее слияние с которым происходит благодаря действию ДНК-лигазы.
Репаративные механизмы ДНК
Иногда в растущую цепь случайно вклинивается неправильное основание, однако, к счастью, у здоровых клеток присутствуют пострепликационные репаративные ферменты и система коррекции ошибочного спаривания оснований, которые исправляют подобные ошибки. В основе механизма действия данных систем лежат удаление и замена ошибочно вставленных оснований в соответствии с последовательностью матричной цепи. Для их функционирования необходимы ДНК-полимеразы b и е.
Значение ДНК для медицины. Патология пострепликационных механизмов репарации иногда обусловливает предрасположенность пациентов к некоторым онкологическим заболеваниям. К ним относят синдром множественной ломкости хромосом (синдром Блума), наследственную предрасположенность к раку молочной железы, вызванную мутациями генов BRCA1 и BRCA2, и аутосомно-доминантную форму рака кишечника (наследственный неполипозный рак толстой кишки).
Существует теория, утверждающая, что после каждого клеточного цикла теломеры укорачиваются на один повтор, а следовательно, количество делений клетки ограничено числом повторов в теломерной цепи. Согласно этому бесконечный рост и деление опухолевых клеток происходят из-за присутствия активных мутантных теломераз, которые препятствуют разрушению теломер.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в содержание раздела «генетика» на нашем сайте
Источник
Репликация ДНК
Репликация — это механизм самокопирования и основное свойство наследственного материала, которым выступают молекулы ДНК.
Особенностью ДНК является то, что обычно ее молекулы состоит из двух комплементарных друг другу цепей, образующих двойную спираль. В процессе репликации цепи материнской молекулы ДНК расходятся, и на каждой строится новая комплементарная цепь. В результате из одной двойной спирали образуется две, идентичные исходной. Т. е. из одной молекулы ДНК образуются две, идентичные матричной и между собой.
Таким образом, репликация ДНК происходит полуконсервативным способом, когда каждая дочерняя молекула содержит одну материнскую цепь и одну вновь синтезированную.
У эукариот репликация происходит в S-фазе интерфазы клеточного цикла.
Описанный ниже механизм и основные ферменты характерны для подавляющего большинства организмов. Однако бывают исключения, в основном среди бактерий и вирусов.
Расхождение цепей исходной молекулы ДНК обеспечивает фермент геликаза, или хеликаза, который в определенных местах хромосом разрывает водородные связи между азотистыми основаниями ДНК. Хеликазы перемещаются по ДНК с затратой энергии АТФ.
Чтобы цепочки снова не соединились, они удерживаются на расстоянии друг от друга дестабилизирующими белками. Белки выстраиваются в ряд со стороны пентозо-фосфатного остова цепи. В результате образуются зоны репликации, называемые репликационными вилками.
Репликационные вилки образуются не в любых местах ДНК, а только в точках начала репликации, состоящих из определенной последовательности нуклеотидов (около 300 штук). Такие места распознаются специальными белками, после чего образуется так называемый репликационный глаз, в котором расходятся две цепи ДНК.
Из точки начала репликация может идти как в одном, так и в двух направлениях по длине хромосомы. В последнем случае цепи ДНК расходятся вперед и назад, и из одного репликационного глазка образуются две репликационные вилки.
Репликон — единица репликации ДНК, от точки ее начала и до точки ее окончания.
Поскольку в ДНК цепи спирально закручены относительно друг друга, то разделение их хеликазой вызывает появление дополнительных витков перед репликационной вилкой. Чтобы снять напряжение, молекула ДНК должна была бы проворачиваться вокруг своей оси один раз на каждые 10 пар разошедшихся нуклеодидов, именно столько образуют один виток спирали. В таком случае ДНК бы быстро вращалась с затратой энергии. Но этого не происходит, т. к. природа нашла более эффективный способ справится с возникающим при репликации напряжением спирали.
Фермент топоизомераза разрывает одну из цепей ДНК. Отсоединенный участок проворачивается на 360° вокруг второй целой цепи и снова соединяется со своей цепью. Этим снимается напряжение, т. е. устраняются супервитки.
Каждая отдельная цепь ДНК старой молекулы используется в качестве матрицы для синтеза новой комплементарной себе цепи. Добавление нуклеотидов к растущей дочерней цепи обеспечивает фермент ДНК-полимераза. Существует несколько разновидностей полимераз.
В репликационной вилке к освободившимся водородным связям цепей согласно принципу комплиментарности присоединяются свободные нуклеотиды, находящиеся в нуклеоплазме. Присоединяющиеся нуклеотиды представляют собой дезоксирибонуклеозидтрифосфаты (дНТФ), а конкретно дАТФ, дГТФ, дЦТФ, дТТФ.
После образования водородных связей фермент ДНК-полимераза связывает нуклеотид фосфоэфирной связью с последним нуклеотидом синтезируемой дочерней цепи. При этом отделяется пирофосфат, включающий два остатка фосфорной кислоты, который потом расщепляется на отдельные фосфаты. Реакция отщепления пирофосфата в результате гидролиза энергетически выгодна, так как связь между первым, который уходит в цепь, и вторым фосфатными остатками богата энергией. Эта энергия используется полимеразой.
Полимераза не только удлиняет растущую цепь, но и способна отсоединять ошибочные нуклеотиды, т. е. обладает корректирующей способностью. Если последний нуклеотид, который должен быть присоединен к новой цепи, не комплементарен матричному, то полимераза его удалит.
ДНК-полимераза может присоединять нуклеотид только к -OH группе, находящейся при 3-м атоме углерода дезоксирибозы. Таким образом цепь синтезируется только со стороны своего 3´-конца. То есть синтез новой цепи ДНК идет в направлении от 5´- к 3´-концу. Поскольку в двуцепочечной молекуле ДНК цепи антипараллельны, то процесс синтеза по материнской, или матричной, цепи идет в обратном направлении – от 3´- к 5´-концу.
Поскольку цепи ДНК антипараллельны, а синтез новой цепи возможен только в направлении 5´→3´, то в репликационной вилке дочерние цепи будут синтезироваться в разных направлениях.
На матрице 3´→5´ сборка новой полинуклеотидной последовательности происходит по большей части непрерывно, так как эта цепь синтезируется в направлении 5´→3´. Антипараллельная матрица характеризуется 5´→3´ направлением, поэтому синтез дочерней цепи по ходу движения вилки здесь не возможен. Здесь он был бы 3´→5´, но ДНК-полимера не может присоединять к 5´-концу.
Поэтому синтез на матрице 5´→3´ выполняется небольшими участками — фрагментами Оказаки (названы в честь открывшего их ученого). Каждый фрагмент синтезируется в обратном ходу образования вилки направлении, что обеспечивает соблюдение правила сборки от 5´- к 3´-концу.
Другим «недостатком» полимеразы является то, что она не может сама начать синтез участка дочерней цепи. Причина этого кроется в том, что ей необходим -OH-конец нуклеотида, уже соединенного с цепью. Поэтому необходима затравка, или праймер. Им выступает короткая молекула РНК, синтезируемые ферментом РНК-праймазой и спаренная с матричной цепью ДНК. Синтез каждого участка Оказаки начинается со своей РНК-затравки. Та цепь, которая синтезируется непрерывно, обычно имеет один праймер.
После удаления праймеров и застраивания брешей ДНК-полимеразой отдельные участки дочерней цепи ДНК сшиваются между собой ферментом ДНК-лигазой.
Непрерывная сборка идет быстрее, чем фрагментарная. Поэтому одна из дочерних цепей ДНК называется лидирующей, или ведущей, вторая — запаздывающей, или отстающей.
У прокариот репликация протекает быстрее: примерно 1000 нуклеотидов в секунду. В то время как у эукариот только около 100 нуклеотидов. Количество нуклеотидов в каждом фрагменте Оказаки у эукариот составляет примерно до 200, у прокариот — до 2000.
У прокариот кольцевые молекулы ДНК представляют собой один репликон. У эукариот каждая хромосома может содержать множество репликонов. Поэтому синтез начинается в нескольких точках, одновременно или нет.
Ферменты и другие белки репликации действуют совместно, образуя комплекс и двигаясь по ДНК. Всего в процессе участвует около 20 разных белков, здесь были перечислены лишь основные.
Источник



