
Параграф 31. обмен гликогена
Автор текста – Анисимова Елена Сергеевна.
Авторские права защищены. Продавать текст нельзя.
Курсив не зубрить.
Замечания можно присылать по почте: exam_bch@mail.ru
https://vk.com/bch_5
ПАРАГРАФ № 31. См. п. 28-30.
Обмен гликогена.»
Знать формулы глюкозы, глюкозо-6-фосфат и глюкозо-1-фосфата, уметь соединить остатки глюкозы 1,4 и 1,6 связями (фрагмент молекулы гликогена).
31. 1. Структура молекулы гликогена.
Определение. – Гликоген – это полимер, состоящий из остатков глюкозы, соединенных ;-1,4 гликозидными связями в линейных участках и ;-1,6 гликозидными связями в точках ветвления. Гликоген находится в мышцах и печени. При употреблении в пищу мышц и печени гликоген переваривается в ЖКТ до глюкозы – см. № 30.
Структура молекулы гликогена – самый первый остаток глюкозы присоединен к особому небольшому белку, который называется гликогенином и выполняет функцию «затравки» при синтезе молекулы гликогена (в том смысле, что с присоединения глюкозы к гликогенину начинается синтез гликогена).
К первому остатку глюкозы присоединяются еще несколько остатков ;-1,4-связями, образуя первую «ветку» гликогена.
К некоторым остаткам глюкозы первой ветки ;-1,6-гликозидными связями присоединяются остатки глюкозы, которые дают начало новым веткам молекулы гликогена.
В молекуле гликогена различают около 12 концентрических слоёв.
Внешние глюкозные остатки могут отщепляться от молекулы гликогена, превращаясь в глюкозу.
31. 2. Р а с щ е п л е н и е г л и к о г е н а
в печени и мышцах называется лизисом гликогена или ГЛИКОГЕНО/ЛИЗОМ (не путать с гликолизом – расщеплением глюкозы).
При гликогенолизе отщепляются самые внешние остатки глюкозы, «с концов веточек» (поэтому чем больше веточек и 1,6 связей – тем быстрее можно расщепить гликоген).
В клетках мышц глюкозные остатки отщепляются для использования в клетках самих мышц,
а в печени – для выхода глюкозы в кровь при ее дефиците, то есть при гипогликемии, которая возникает при голоде, стрессе, повышенном расходе глюкозы.
Но запасов гликогена печени организму хватает только на 12 часов – после этого глюкозу должен давать глюконеогенез, сырьём для которого служат белки мышц – п.33.
31. 2. 2. Регуляция расщепления гликогена (путём фосфоролиза – см. далее).
Расщепление гликогена (как и глюконеогенез) нужно и происходит при голоде под действием гормона голода глюкагона
и при стрессе под действием гормонов стресса ГКС и катехоламинов адреналина и норадреналина.
При сытости и покое расщепление гликогена не нужно и не происходит, поскольку оно тормозится гормоном покоя и сытости инсулином. При дефиците инсулина или его действия при сахарном диабете расщепление не тормозится инсулином, что приводит к ускорению расщепления гликогена и способствует гипергликемии.
Регуляция расщепления гликогена осуществляется через изменение активности и/или концентрации его ключевых ферментов: гликоген/фосфорилазы и гексозо-6-фосфатазы (см. далее):
инсулин препятствует работе ферментов расщепления гликогена, а глюкагон и ГКС с КА способствуют (ГКС индуцируют глюкозо-6-фосфатазу, а глюкагон и катехоламины активируют гликоген/фосфорилазу, с помощью вторых посредников – цАМФ и ионов кальция).
31. 2. 3. Способы гликогенолиза.
Есть два способа гликогенолиза –
1 – (в печени) если при расщеплении присоединяются молекулы глюкозы, то расщепление называется гидролизом (гликолитическим) и катализируется ферментом ;-амилазой, которая отщепляет по одной молекуле глюкозы;
2 – (в печени и в мышцах) если при расщеплении присоединяются молекулы фосфорной кислоты (Н3РО4), то расщепление называется фосфоролизом или фосфоролитическим и катализируется ферментом, который называется фосфорилазой гликогена.
31. 2. 4. Фосфоролиз гликогена (описание)
Фосфорилаза отщепляет один остаток глюкозы, присоединяя к нему фосфат (в первом положении),
в результате чего продуктами фосфорилазы становятся глюкозо-1-фосфат
и укороченная на один глюкозный остаток молекула гликогена (n-1).
После этого от молекулы гликогена фосфорилазой отщепляются по одному следующие глюкозные остатки, пока не встретится 1,6-связь.
1,6 связь расщепляется так называемым противоветвящим ферментом, после чего 1,4-связи продолжают расщепляться фосфорилазой.
31. 2. 5. Р е а к ц и и ф о с ф о р о л и з а (три):
1-я реакция фосфоролиза:
гликоген(n) + фосфорная кислота (Н3РО4) = гликоген(n-1) и глюкозо-1-фосфат.
Один глюкозный остаток отщепился, к нему присоединился фосфата (без затраты АТФ!),
а в молекуле гликогена стало на один глюкозный остаток меньше (n-1).
2-я реакция фосфоролиза:
перенос фосфата из 1-го положения глюкозо-1-фосфата в 6-е положение, в результате чего глюкозо-1-фофат превращается в глюкозо-6-фосфат. Реакция обратима (обратная протекает при синтезе гликогена), фермент называется фосфоглюкомутазой. Остальные реакции в обмене гликогена необратимы.
Схема реакции: Глюкозо-1-фосфат ; глюкозо-6-фосфат.
3-я реакция фосфоролиза:
фосфат отщепляется от 6-го положения (путем гидролиза), в результате чего образуются фосфорная кислота и глюкоза, способная выйти в кровь для питания мозга и эритроцитов, повысить концентрацию глюкозы в крови.
В этом главное значение гликогенолиза в печени – являться одним их источников глюкозы для организма.
Схема реакции: глюкозо-6-фосфат + Н2О = глюкоза + фосфорная кислота.
Чтобы назвать фермент этой реакции, нужно добавить «аза» к глюкозо-6-фосфату: глюкозо-6-фосфатаза.
Ферменты, которые катализируют отщепление фосфатов (путем гидролиза, дефосфорилирование), называются фосфатазами.
В мышцах нет фермента глюкозо-6-фосфатаза, поэтому в них глюкозо-6-фосфат не превращается в глюкозу,
поэтому гликоген мышц не является резервом глюкозы для других тканей.
Образованный в мышцах глюкозо-6-фосфат вступает в реакции гликолиза, превращаясь в лактат (в анаэробных условиях напряженно работающей мышцы) – п.32.
Фосфорилаза и глюкозо-6-фосфатаза – ключевые фермента фосфоролиза.
31. 3. С и н т е з г л и к о г е н а .
31. 3. 1. Значение. –
Нужен для того, чтобы при голоде или стрессе в организме был резерв глюкозы для мозга и эритроцитов, который предотвратит «голодный обморок» и поддержит работоспособность.
31. 3. 2. Регуляция синтеза гликогена.
Поэтому при стрессе и голоде синтез гликогена не происходит (гормоны голода и стресса снижают синтез гликогена), а в покое и сытости синтез гликогена происходит под влиянием инсулина.
Регуляция синтеза гликогена осуществляется через изменение активности и/или концентрации его ключевых ферментов: гексокиназы и гликоген/синтазы (см. далее):
Инсулин способствует работе ферментов синтеза гликогена, а глюкагон и ГКС с КА препятствуют (ГКС репрессируют гексокиназу, а глюкагон и катехоламины инактивируют гликоген/синтазу с помощью вторых посредников – цАМФ и ионов кальция).
Синтез гликогена – один из процессов, использующих глюкозу, поэтому его протекание способствует снижению концентрации глюкозы в крови.
31. 3. 3. Реакции синтеза гликогена (четыре):
1-я реакция синтеза гликогена:
такая же, как в гликолизе и ПФП (п.32 и 35): присоединение к глюкозе фосфата (фосфорилирование), которое превращает её в глюкозо-6-фосфат. Источником АТФ является фосфат, катализирующие реакции такого типа (перенос фосфата от АТФ на субстрат) ферменты называются киназами; киназа, катализирующая фосфорилирование глюкозы и других гексоз на 6-му положению, называется гексокиназой.
Схема: глюкоза + АТФ ; глюкозо-6-фосфата + АДФ.
2-я реакция синтеза гликогена:
перенос фосфата из 6-го положения в первое, в результате чего глюкозо-6-фосфат превращается в глюкозо-1-фосфат. Эта реакция обратима, в обратную сторону она протекает при расщеплении гликогена (см. выше). Фермент – фосфоглюкомутаза. Остальные реакции синтеза гликогена необратимы.
Глюкозо-6-фосфат ; глюкозо-1-фосфат.
3-я реакция синтеза гликогена:
Образование УДФ-глюкозы из глюкозо-1-фосфата в результате присоединения к фосфату УМФ (п.70). Источником УМФ является УТФ поэтому УТФ называют макроэргом углеводного обмена. Затраты УТФ приравниваются к затратам АТФ. Расщепление УТФ до УМФ равнозначно трате двух АТФ. Таким образом, при синтезе гликогена на присоединение каждой молекулы глюкозы тратится 3 молекулы АТФ (третья – в первой реакции).
Глюкозо-1-фосфат + УТФ ;глюкозо-1-фосфат-УМФ (=УДФ-глюкоза) + ФФн
4-я реакция синтеза гликогена:
Глюкоза отщепляется от УДФ и переносится на растущую цепочку молекулы гликогена, присоединяясь к нему 1,4-гликозидной связью.
УДФ-глюкоза + гликоген с n-количеством глюкозных остатков ;
; УДФ + гликоген с (n+1) количеством глюкозных остатков.
31. 4. Гликогенозы и агликогенозы.
Встречаются люди с низкой активностью ферментов, участвующих в расщеплении гликогена
(гликоген/фосфорилазы и глюкозо-6-фосфатазы; второй ещё в ГНГ работает п.33) – из-за этого у них гликоген не расщепляется (путем фосфоролиза), накапливается в печени – это накопление называется ГЛИКОГЕНОЗОМ.
При гликогенозе не может образовываться глюкоза за счет расщепления гликогена, поэтому у людей с гликогенозом снижена способность переносить обычные перерывы в приемах пищи, поэтому им необходимо есть чаще, чем обычным людям (есть углеводную) пищу. Больший перерыв в приеме пищи может привести у таких людей к снижению концентрации глюкозы в крови (гипогликемии), появлению слабости, обмороку. Накопление гликогена приводит также к увеличению печени.
Гликогеноз – это пример метаболического блока: низкой скорости реакции из-за низкой активности фермент (из-за мутации гена). Пример первичной энзимопатии.
Дефицит глюкозо-6-фосфатазы тяжелее, так как в этом случае глюкоза не образуется и при ГНГ. Вся надежда на регулярное питание.
Встречаются люди со сниженной активностью фермента синтеза гликогена гликоген/синтазы из-за мутации кодирующего его гена. У них гликоген не синтезируется (или мало), поэтому и расщепляться при голоде не может.
Это отсутствие гликогена называется А-ГЛИКОГЕНОЗОМ (приставка «а-» означает отсутствие).
При агликогенозе образ жизни такой же, как при гликогенозе – нужно регулярно питаться, так как резерва глюкозы (гликоген) на случай голода отсутствует. Возможно, помогает ГНГ.
Источник
Р ЕГУЛЯЦИЯ ОБМЕНА ГЛИКОГЕНА
Метаболизм гликогена в печени регулируется несколькими гормонами, одни из которых активируют ферменты синтеза гликогена, а другие – ферменты распада гликогена.
Основные ферменты метаболизма гликогена активны либо в фосфорилированной, либо в дефосфорилированной форме. Присоединение фосфатов к ферменту производят протеинкиназы , источником фосфора является АТФ:
o фосфорилаза гликогена активируется после присоединения фосфатной группы, o синтаза гликогена после присоединения фосфата инактивируется .
Скорость фосфорилирования указанных ферментов повышается после воздействия на клетку адреналина, глюкагона и некоторых других гормонов. В результате адреналин и глюкагон вызывают гликогенолиз, активируя фосфорилазу гликогена.
Дефосфорилирование ферментов осуществляют протеинфосфатазы . Активатором протеинфосфатаз выступает инсулин . Вместе с этим, инсулин и глюкокортикоиды ускоряют синтез гликогена, увеличивая количество молекул гликогенсинтазы .
В клетке осуществляется реципрокная (взаимоисключающая) регуляция. При работе клетки и/или гормональных влияниях активируется протеинкиназа и, как следствие, активируется фосфорилаза гликогена и ингибируется гликогенсинтаза. Идут реакции катаболизма глюкозы, и образуется энергия. При отдыхе или в покое работают протеин-фосфатазы, которые освобождают ферменты от фосфорной кислоты: в результате гликоген-фосфорилаза дефосфорилируется и становится не активной, активируется гликогенсинтаза . Начинается запасание глюкозы в виде гликогена.
Для регуляции активности фосфорилазы и синтазы гликогена используется особый каскадный аденилатциклазный механизм.
А КТИВАЦИЯ ФОСФОРИЛАЗЫ ГЛИКОГЕНА
Скорость гликогенолиза лимитируется только скоростью работы фосфорилазы гликогена. Ее активность может изменяться тремя способами: ковалентная модификация, кальцийзависимая активация и аллостерическая активация с помощью АМФ.
Источник
Регуляция активности гликогенфосфорилазы и глигогенсинтазы
Действие этих гормонов в конечном счете сводится к изменению скорости реакций, катализируемых ключевыми ферментами метаболических путей обмена гликогена – гликогенсинтазой и гликогенфосфорилазой, активность которых регулируется аллостерически и фосфорилированием/ десфорилированием.
Гликогенфосфорилаза существует в 2 формах:
1) Фосфорилированная — активная (форма а); 2) дефосфорилированная — неактивная (форма в).
Фосфорилирование осуществляется путём переноса фосфатного остатка с АТФ на гидроксильную группу одного из сериновых остатков фермента. Следствие этого — конформационные изменения молекулы фермента и его активация.
Взаимопревращения 2 форм гликогенфосфорилазы обеспечиваются действием ферментов киназы фосфорилазы и фосфопротеинфосфатазы (фермент, структурно связанный с молекулами гликогена). В свою очередь, активность киназы фосфорилазы и фосфопротеинфосфатазы также регулируется путём фосфорилирования и дефосфорилирования.
Активация киназы фосфорилазы происходит под действием протеинкиназы А — ПКА (цАМФ-зависимой). цАМФ сначала активирует протеинкиназу А, которая фосфорилирует киназу фосфорилазы, переводя её в активное состояние, а та, в свою очередь, фосфорилирует гликогенфосфорилазу. Синтез цАМФ стимулируется адреналином и глюкагоном.
Активация фосфопротеинфосфатазы происходит в результате реакции фосфорилирования, катализируемой специфической протеинкиназой, которая, в свою очередь, активируется инсулином посредством каскада реакций с участием других белков и ферментов. Активируемая инсулином протеинкиназа фосфорилирует и тем самым активирует фосфопротеинфосфатазу. Активная фосфопротеинфосфатаза дефосфорилирует и, следовательно, инактивирует киназу фосфорилазы и гликогенфосфорилазу.

Влияние инсулина на активность гликогенсинтазы и киназы фосфорилазы. ФП-фосфатаза (ГР) — фосфопроте-инфосфатаза гранул гликогена. ПК (pp90S6) — протеинкиназа, активируемая инсулином.
Активность гликогенсинтазы также изменяется в результате фосфорилирования и дефосфорилирования. Однако есть существенные различия в регуляции гликогенфосфорилазы и гликогенсинтазы:
фосфорилирование гликогенсинтазы катализирует ПК А и вызывает её инактивацию;
дефосфорилирование гликогенсинтазы под действием фосфопротеинфосфатазы, наоборот, её активирует.
Регуляция метаболизма гликогена в печени
Повышение уровня глюкозы в крови стимулирует синтез и секрецию β-клетками поджелудочной железы гормона инсулина. Инсулин передает сигнал в клетку через мембранный каталитический рецептор — тирозиновую протеинкиназу. Взаимодействие рецептора с гормоном инициирует ряд последовательных реакций, приводящих к активации фосфопротеинфосфатазы гранул гликогена. Этот фермент дефосфорилирует гликогенсинтазу и гликогенфосфорилазу, в результате чего гликогенсинтаза активируется, а гликогенфосфорилаза становится неактивной.
Таким образом, в печени ускоряется синтез гликогена и тормозится его распад.
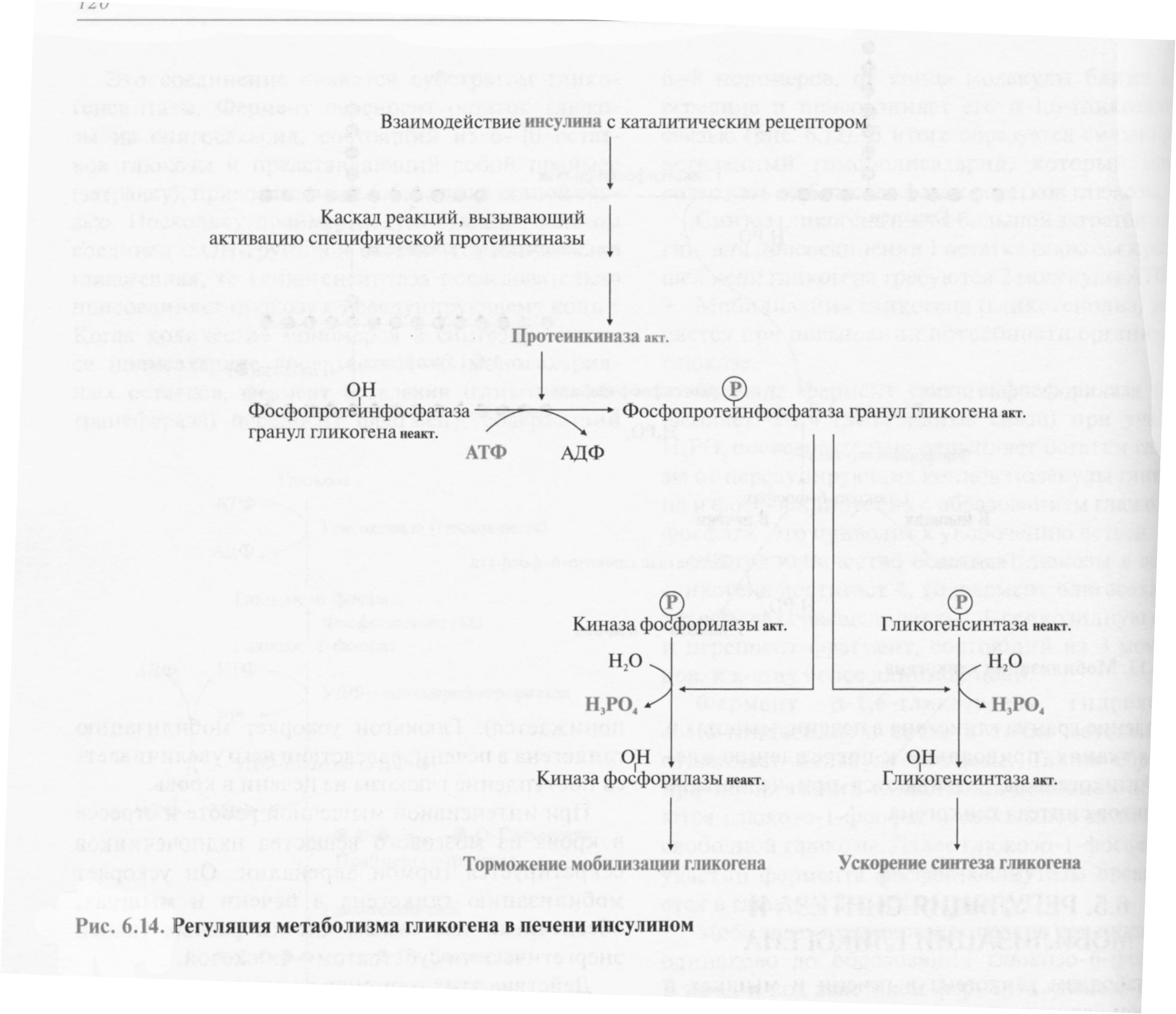
При голодании снижение уровня глюкозы в крови является сигналом для синтеза и секреции α-клетками поджелудочной железы глюкагона. Гормон передает сигнал в клетки через аденилатциклазную систему. Это приводит к активации проттеинкиназы А, которая фосфорилирует гликогенсинтазу и киназу фосфорилазы. В результате фосфорилирования гликогенсинтаза инактивируется и синтез гликогена тормозится, а киназа фосфорилазы становится активной и фосфорилирует гликогенфосфорилазу, которая становится активной. Активная гликогенфосфорилаза ускоряет мобилизацию гликогена в печени.

1 — глюкагон и адреналин взаимодействуют со специфическими мембранными рецепторами. Комплекс гормон-рецептор влияет на конформацию G-белка, вызывая диссоциацию его на протомеры и замену в α-субъединице ГДФ на ГТФ;
2 — α-субъединица, связанная с ГТФ, активирует аденилатциклазу, катализирующую синтез цАМФ из АТФ;
3 — в присутствии цАМФ протеинкиназа А обратимо диссоциирует, освобождая обладающие каталитической активностью субъединицы С;
4 — протеинкиназа А фосфорилирует и активирует киназу фосфорилазы;
5 — киназа фосфорилазы фосфорилирует гликогенфосфорилазу, переводя её в активную форму;
6 — протеинкиназа А фосфорилирует также гликогенсинтазу, переводя её в неактивное состояние;
7 — в результате ингибирования гликогенсинтазы и активации гликогенфосфорилазы гликоген включается в процесс распада;
8 — фосфодиэстераза катализирует распад цАМФ и тем самым прерывает действие гормонального сигнала. Комплекс α-субъединица-ГТФ затем распадается.
При интенсивной физическеой работе и стрессе в крови повышается концентрация адреналина. В печени есть два типа мембранных рецепторов адреналина. Эффект адреналина в печени обусловлен фосфорилированием и активацией гликогенфосфорилазы. Адреналин имеет сходный с глюкагоном механизм действия. Но возможно включение и другой эффекторной системы передачи сигнала в клетку печени.

Регуляция синтеза и распада гликогена в печени адреналином и Са 2+ .
ФИФ2— фосфатидилинозитолбисфосфат; ИФ3— инозитол-1,4,5-трифосфат; ДАГ — диацилглицерол; ЭР — эндоплазматический ретикулум; ФС — фосфодитилсерин.
1 — взаимодействие адреналина с α1-рецептором трансформирует сигнал через активацию G-белка на фосфолипазу С, переводя её в активное состояние;
2 — фосфолипаза С гидролизует ФИФ2 на ИФ3 и ДАГ;
3 — ИФ3 активирует мобилизацию Са 2+ из ЭР;
4 — Са 2+ , ДАГ и фосфодитилсерин активируют протеинкиназу С. Протеинкиназа С фосфорилирует гликогенсинтазу, переводя её в неактивное состояние;
5 — комплекс 4Са 2+ — кальмодулин активирует киназу фосфорилазы и кальмодулин-зависимые протеинкиназы;
6 — киназа фосфорилазы фосфорилирует гликогенфосфорилазу и тем самым её активирует;
7 — активные формы трёх ферментов (кальмодулинзависимая протеинкиназа, киназа фосфорилазы и протеинкиназа С) фосфорилируют гликогенсинтазу в различных центрах, переводя её в неактивное состояние.
Какая система передачи сигнала в клетку будет использована, зависит от типа рецепторов, с которыми взаимодействует адреналин. Так, взаимодействие адреналина с β2-рецепторами клеток печени приводит в действие аденилатциклазную систему. Взаимодействие же адреналина с α1-рецепторами «включают» инозитолфосфатный механизм трансмембранной передачи гормонального сигнала. Результат действия обеих систем — фосфорилирование ключевых ферментов и переключение процессов с синтеза гликогена на его распад. Следует отметить, что тип рецепторов, который в наибольшей степени вовлекается в ответ клетки на адреналин, зависит от концентрации его в крови.
В период пищеварения преобладает влияние инсулина, так как инсулин-глюкагоновый индекс в этом случае повышается. В целом инсулин влияет на обмен гликогена противоположно глюкагону. Инсулин снижает концентрацию глюкозы в крови в период пищеварения, действуя на метаболизм печени следующим образом:
снижает уровень цАМФ в клетках и тем самым активируя протеинкиназу В. Протеинкиназа В, в свою очередь, фосфорилирует и активирует фосфодиэстеразу цАМФ — фермент, гидролизующий цАМФ с образованием АМФ;
активирует фосфопротеинфосфатазу гранул гликогена, которая дефосфорилирует гликогенсинтазу и таким образом её активирует. Кроме того, фосфопротеинфосфатаза дефосфорилирует и, следовательно, инактивирует киназу фосфорилазы и гликогенфосфорилазу;
индуцирует синтез глюкокиназы, тем самым ускоряя фосфорилирование глюкозы в клетке.
Источник



