
Способ получения энергии хемотрофы
Преобладающую часть бактерий составляют бактерии хемотрофы, получающие энергию в результате окислительно-восстановительных реакций расщепления химических веществ, которые в ряде случаев служат для них также источником питания. Разные бактерии получают энергию либо в процессе брожения либо в процессе дыхания.
При брожении АТФ образуется исключительни путём субстратного фосфорилирования, а в процессе дыхания преимущественно путём окислительного фосфорилирования за исключением начальных этапов превращения гексоз в триозы (гликолиз, см. ниже). Получение энергии субстратным фосфорилированием
Субстратное фосфорилирование может происходить при различных реакциях промежуточного метаболизма. При дегидрировании некоторых определённых субстратов часть энергии, освободившейся при окислении, сохраняется в форме высокоэнергетического фосфата. Богатая энергией фосфатная группа затем переносится на АДФ с образованием АТФ. Такой процесс называют фосфорилированием на уровне субстрата (субстратное фосфорилирование).
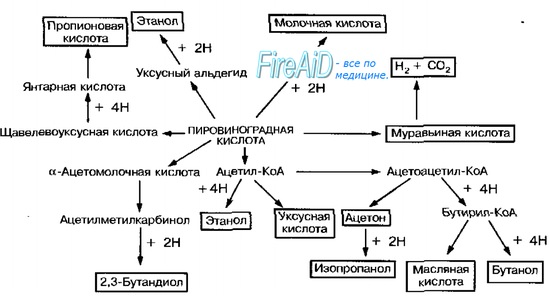 Рис. 4-7. Пути восстановления пирувата в процессе брожения.
Рис. 4-7. Пути восстановления пирувата в процессе брожения.
В обмене углеводов важнейшие реакции, приводящие к регенерации АТФ, катализируют фосфоглицераткиназа (1,3-бифосфоглицерат + АДФ — 3-фосфоглицерат + АТФ), пируваткиназа (фосфоэнолпируват + АДФ = пируват + АТФ) и ацетаткиназа (ацетилфосфат или бутирилфосфат + АДФ = ацетат или бутират + АТФ). Бактерии и дрожжи, сбраживающие сахара, располагают лишь тем АТФ, который получается с помощью этих ферментов.
Брожение. При брожении происходит анаэробное разложение углеводов и образование АТФ посредством субстратного фосфорилирования. Брожение характерно для факультативных и облигатных анаэробов. При брожении продукты расщепления органического субстрата могут служить одновременно и донорами и акцепторами водорода. Отдельные этапы окисления представляют собой дегидрирование, при котором водород переносится на НАД. Акцепторами водорода в составе НАДН+ служат промежуточные продукты расщепления субстрата. Кислород подавляет брожение, и оно у факультативных анаэробов сменяется дыханием.
По выходу энергии брожение уступает дыхательному метаболизму: при сбраживании микроорганизмами 1 моля глюкозы образуется от 1 до 4 молей АТФ. Для сравнения; у дрожжей на 1 моль сброженной глюкозы образуется 2 моля АТФ, а при дыхании — 38. При ферментации субстратов в среде накапливаются конечные продукты (лактат, бутират, ацетон, 2-пропанол, этанол, ацетоин и др.). Определение природы этих продуктов— важный момент для идентификация анаэробов. Например, образование ацетоина определяют в реакции Фогеса-Проскауэра, кислотообразование — в тесте с метиловым красным, индолообразование — по методу Грациана. Применяют и другие тесты, разработанные для идентификации бактерий.
Любое брожение проходит две стадии: первая (окисление) включает превращение глюкозы в пировиноградную кислоту, вторая (восстановление) — присоединение атомов водорода для восстановления пировиноградной кислоты (рис. 4-7). Сам процесс образования пировиноградной кислоты включает серию реакций, общих для брожения и аэробного дыхания. У микроорганизмов выделяют три пути образования триоз (пировиноградной кислоты и глицеральдегидфосфата) из углеводов: гликолитичес-кий (фруктозо-1,6-бифосфатный), пентозофосфатный и 2-кето-З-дезокси-б-фосфоглюконатный (КДФГ-путь, путь Энтнера-Дудорова-Парнаса). Согласно образующимся продуктам, брожение разделяют на следующие типы.
Источник
Энергия в клетке. Использование и хранение

Всем привет! Эту статью я хотел посвятить клеточному ядру и ДНК. Но перед этим нужно затронуть то, как клетка хранит и использует энергию (спасибо spidgorny). Мы будем касаться вопросов связанных с энергией почти везде. Давайте заранее в них разберемся.
Из чего можно получать энергию? Да из всего! Растения используют световую энергию. Некоторые бактерии тоже. То есть органические вещества синтезируются из неорганических за счет световой энергии. + Есть хемотрофы. Они синтезируют органические вещества из неорганических за счет энергии окисления аммиака, сероводорода и др. веществ. А есть мы с вами. Мы — гетеротрофы. Кто это такие? Это те, кто не умеет синтезировать органические вещества из неорганических. То есть хемосинтез и фотосинтез, это не для нас. Мы берем готовую органику (съедаем). Разбираем ее на кусочки и либо используем, как строительный материал, либо разрушаем для получения энергии.
Что конкретно мы можем разбирать на энергию? Белки (сначала разбирая их на аминокислоты), жиры, углеводы и этиловый спирт (но это по желанию). То есть все эти вещества могут быть использованы, как источники энергии. Но для ее хранения мы используем жиры и углеводы. Обожаю углеводы! В нашем теле основным запасающим углеводом является гликоген.

Он состоит из остатков глюкозы. То есть это длинная, разветвленная цепочка, состоящая из одинаковых звеньев (глюкозы). При необходимости в энергии мы отщепляем по одному кусочку с конца цепи и окисляя его получаем энергию. Такой способ получения энергии характерен для всех клеток тела, но особенно много гликогена в клетках печени и мышечной ткани.
Теперь поговорим о жире. Он хранится в специальных клетках соединительной ткани. Имя им — адипоциты. По сути это клетки с огромной жировой каплей внутри.
При необходимости, организм достает жир из этих клеток, частично расщепляет и транспортирует. По месту доставки происходит окончательное расщепление с выделением и преобразованием энергии.
Довольно популярный вопрос: «Почему нельзя хранить всю энергию в виде жира, или гликогена?»
У этих источников энергии разное назначение. Из гликогена энергию можно получить довольно быстро. Его расщепление начинается почти сразу после начала мышечной работы, достигая пика к 1-2 минуте. Расщепление жиров протекает на несколько порядков медленней. То есть если вы спите, или медленно куда-то идете — у вас постоянный расход энергии, и его можно обеспечить расщепляя жиры. Но как только вы решите ускориться (упали сервера, побежали поднимать), резко потребуются много энергии и быстро ее получить расщепляя жиры не получится. Тут нам и нужен гликоген.
Есть еще одно важное различие. Гликоген связывает много воды. Примерно 3 г воды на 1 г гликогена. То есть, для 1 кг гликогена это уже 3 кг воды. Не оптимально… С жиром проще. Молекулы липидов (жиры=липиды), в которых запасается энергия не заряжены, в отличие от молекул воды и гликогена. Такие молекулы называется гидрофобными (дословно, боящимися воды). Молекулы воды же поляризованы. Примерно так это выглядит.
По сути, положительно заряженные атомы водорода взаимодействуют с отрицательно заряженными атомами кислорода. Получается стабильное и энергетически выгодное состояние.
Теперь представим молекулы липидов. Они не заряжены и не могут нормально взаимодействовать с поляризованными молекулами воды. Поэтому смесь липидов с водой энергетически невыгодна. Молекулы липидов не способны адсорбировать воду, как это делает гликоген. Они «кучкуются» в так называемые липидные капли, окружаются мембраной из фосфолипидов (одна их сторона заряжена и обращена к воде снаружи, вторая — не заряжена и смотрит на липиды капли). В итоге, у нас есть стабильная система, эффективно хранящая липиды и ничего лишнего.
Окей, мы разобрались с тем, в каких формах хранится энергия. А что с ней происходит дальше? Вот отщепили мы молекулу глюкозы от гликогена. Превратили ее в энергию. Что это значит?
Сделаем небольшое отступление.
В клетке происходит порядка 1.000.000.000 реакций каждую секунду. При протекании реакции одно вещество трансформируется в другое. Что при этом происходит с его внутренней энергией? Она может уменьшаться, увеличиваться или не меняться. Если она уменьшается -> происходит выделение энергии. Если увеличивается -> нужно взять энергию из вне. Организм обычно совмещает такие реакции. То есть энергия, выделившаяся при протекании одной реакции идет на проведение второй.
Так вот в организме есть специальные соединения, макроэрги, которые способны накапливать и передавать энергию в ходе реакции. В их составе есть одна, или несколько химических связей, в которых и накапливается эта энергия. Теперь можно вернуться к глюкозе. Энергия выделившаяся при ее распаде запасется в связях этих макроэргов.
Разберем на примере.
Самым распространенным макроэргом (энергетической валютой) клетки является АТФ (Аденозинтрифосфат).
Выглядит примерно так.
В его состав входит азотистое основание аденин (одно из 4, используемых для кодирования информации в ДНК), сахар рибоза и три остатка фосфорной кислоты (поэтому и АденозинТРИфосфат). Именно в связях между остатками фосфорной кислоты накапливается энергия. При отщеплении одного остатка фосфорной кислоты образуется АДФ (АденозинДИфосфат). АДФ может выделять энергию, отрывая еще один остаток и превращаясь в АМФ (АденозинМОНОфосфат). Но эффективность отщепленная второго остатка намного ниже. Поэтому, обычно, организм стремится из АДФ снова получить АТФ. Происходит это примерно так. При распаде глюкозы, выделяющаяся энергия тратится на образование связи между двумя остатками фосфорной кислоты и образование ATP. Процесс многостадийный и пока мы его опустим.
Получившийся АТФ является универсальным источником энергии. Он используется везде, начиная от синтеза белка (для соединения аминокислот нужна энергия), заканчивая мышечной работой. Моторные белки, осуществляющие мышечное сокращение используют энергию, запасенную в АТФ, для изменения своей конформации. Изменение конформации это переориентация одной части большой молекулы относительно другой. Выглядит примерно так.
То есть химическая энергия связи переходит в механическую энергию. Вот реальные примеры белков, использующих АТФ для осуществления работы.
Знакомьтесь, это миозин. Моторный белок. Он осуществляет перемещение крупных внутриклеточных образований и участвует в сокращении мышц. Обратите внимание, у него имеется две «ножки». Используя энергию запасенную в 1 молекуле АТФ он осуществляет одно конформационное изменение, по сути один шаг. Самый наглядный пример перехода химической энергии АТФ в механическую.
Второй пример — Na/K насос. На первом этапе он связывает три молекулы Na и одну АТФ. Используя энергию АТФ, он меняет конформацию, выбрасывая Na из клетки. Затем он связывает две молекулы калия и, возвращаясь к исходной конформации, переносит калий в клетку. Штука крайне важная, позволяет поддерживать уровень внутриклеточного Na в норме.
А если серьезно, то:
Пауза. Зачем нам АТФ? Почему мы не можем использовать запасенную в глюкозе энергию напрямую? Банально, если окислить глюкозу до CO2 за один раз, мгновенно выделится экстремально много энергии. И большая ее часть рассеется в виде тепла. Поэтому реакция разбивается на стадии. На каждой выделяется немного энергии, она запасается, и реакция продолжается пока вещество полностью не окислиться.
Подитожу. Запасается энергия в жирах и углеводах. Из углеводов ее можно извлечь быстрее, но в жирах можно запасти больше. Для проведения реакций клетка использует высокоэнергетические соединения, в которых запасается энергия распада жиров, углеводов и тд… АТФ — основное такое соединение в клетке. По сути, бери и используй. Однако не единственное. Но об этом позже.
Источник
Принципы получения энергии в живых объектах.
Живые организмы с точки зрения термодинамики – открытые системы. Между системой и окружающей средой возможен обмен энергии, который происходит в соответствии с законами термодинамики. Каждое органическое соединение, поступающее в организм, обладает определенным запасом энергии (Е). Часть этой энергии может быть использована для совершения полезной работы. Такую энергию называют свободной энергией (G). Направление химической реакции определяется значением DG. Если эта величина отрицательна, то реакция протекает самопроизвольно. Такие реакции называются экзергоническими. Если DG положительно, то реакция будет протекать только при поступлении свободной энергии извне – это эндергонические реакции.
По способу получения энергии все живые организмы делятся на автотрофов и гетеротрофов. Автотрофные организмынеобходимый им углерод получают из СО2 атмосферы, из которого строят все углеродсодержащие компоненты. Источником энергии служит солнечный свет, который улавливается в процессе фотосинтеза, поглощается специальными светочувствительными белками (хлорофилл зелёных растений). К автотрофным организмам относятся: зелёные растения, сине-зелёные водоросли, зелёные и пурпурные бактерии.
Автотрофные организмы также используют различные способы получения энергии. По используемому источнику энергии автотрофы делятся на фототрофов и хемотрофов. Фототрофы получают энергию для восстановления углекислого газа до органических соединений в виде света (растения и некоторые группы бактерий, прежде всего, цианобактерии). Эти организмы являются основным источником органического вещества в биосфере.
Другая группа автотрофов использует для своей жизнедеятельности энергию, выделяющуюся при окислении неорганических веществ. Их называют хемотрофами. Представителями этой группы являются только бактерии. Каждый вид таких бактерий способен использовать только один источник энергии. Используемые ими вещества должны в достаточных количествах присутствовать в окружающей среде.
Открытие бактериального хемосинтеза принадлежит С.Н. Виноградскому. Он впервые доказал существование в природе автотрофных незеленых организмов — серо-, железо- и нитрифицирующих бактерий и описал сущность процесса хемосинтеза.
Известны также бактерии-хемосинтетики, способные окислять метан, окись углерода, водород.
Гетеротрофные организмыполучают углерод в виде сложных соединений (белков, углеводов, липидов и др.), которые содержатся в пищевых продуктах. Энергию гетеротрофы получают в экзэргонических реакциях, которые происходят в митохондриях. К гетеротрофным организмам относятся животные организмы, в том числе человек. Среди гетеротрофов есть такие, которые нуждаются лишь в одном или нескольких простых органических веществах. Например, пекарские дрожжи могут жить, получая только глюкозу и минеральные соли, а кишечная палочка — только ацетат натрия (соль уксусной кислоты). Другие гетеротрофы не могут образовывать целый ряд необходимых им органических веществ. Например, млекопитающие нуждаются в 10 незаменимых аминокислотах (фен, мет, лей, тре, три, иле, вал, лиз, арг, гис), ряде ненасыщенных жирных кислот и витаминов. В настоящее время гетеротрофы подразделяются на:
— органоавтотрофов (употребляющие органические вещества как источники энергии),
— литогетеротрофов (использующие органические вещества как источники углерода),
— органогетеротрофов (органические вещества служат для них источниками энергии и источниками углерода).
Энергетический обмен также различен у разных групп организмов. Гетеротрофные организмы получают энергию за счет окисления органических веществ, поступающих извне. Хорошим окислителем является молекулярный кислород, при кислородном окислении выделяется много энергии. Однако кислород в силу своей реакционной способности повреждает важные вещества клетки (белки, липиды, ДНК) и поэтому токсичен. По отношению к молекулярному кислороду организмы подразделяются на:
1. Анаэробы. Они способны жить в отсутствие кислорода, осуществляя реакции внутримолекулярного окисления и восстановления или используя в качестве окислителей другие вещества среды, например нитраты (так называемое нитратное дыхание).
2. Аэробы — большинство ныне живущих гетеротрофных организмов, которые осуществляют энергетический обмен, окисляя органические вещества молекулярным кислородом. В клетках таких организмов обязательно существуют системы защиты от окисления клеточных структур кислородом. Это, во-первых, ферменты, разрушающие активные формы кислорода (супероксиддисмутаза, каталаза), а во-вторых, антиоксиданты — молекулы, которые сами окисляются, нейтрализуя свободные радикалы.
Носители энергии
Макроэргические соединения– это вещества, содержащие богатые энергией связи. Макроэргическая связь обозначается символом
(знак «тильда»). Понятие макроэргическая связь довольно условно и применяется для обозначения тех связей, которые гидролизуются в водной средес выделением значительной энергии. Так, гидролиз концевой фосфоангидридной связи АТФ (АТФ + Н2О → АДФ + Фн), ведет к освобождению 34,5 кДж/моль энергии). Однако, если реакция протекает в неводной среде (например, в липидном слое мембран), то образование и разрушение АТФ протекает без больших затрат энергии.
Вещества, гидролиз которых, приводит к высвобождению более 21 кДж/моль (>5ккал) энергии относят к высокоэнергетическим (макроэргам), а вещества освобождающие меньшие количества энергии — к низкоэнергетическим. К макроэргам относят: АТФ, другие трифосфаты нуклеозидов (ГТФ, ЦТФ, УТФ, ТТФ), аргининфосфат, креатинфосфат, ацетилфосфат, 1,3-дифосфоглицерат, фосфоенолпируват и др. К группе высокоэнергетических соединений относятся также тиоэфиры. В то время как высокоэнергетические фосфорные соединения являются, в основном, промежуточными продуктами углеводного обмена, то тиоэфиры — это промежуточные продукты обмена жирных кислот. В природе основным тиосодержащим соединением является кофермент А (HS-CoA).
К низкоэнергетическим — глюкозо-6-фосфат, глюкозо-1-фосфат, глицерофосфат и другие (см. таблицу). Уникальная роль АТФ состоит в том, что она имеет промежуточное значение энергии гидролиза и выполняет роль связующего звена (разменной монеты) между высоко- и низкоэнергетическими соединениями.

АТФ с точки зрения химического строения представляет собой адениловый нуклеозид, связанный с тремя остатками фосфорной кислоты. Первый из них (α-остаток) связан сложноэфирной связью с пятым углеродным атомом пентозы. Это обычная связь. Изменение свободной энергии реакции ΔG о ′ не превышает 12,5 кДж/моль, как и у большинства связей в органических соединениях. Второй фосфатный остаток (β) и третий (γ) связаны с α-остатком и друг с другом фосфоангидридными связями, у которых ΔG о ′ (при рН 7, температуре 37 о , концентрации реагентов 1М и избытке Mg 2+ ) составляет (-30,6) кДж/моль (макроэргические связи).
 Они легко гидролизуются с выделением энергии. В клетке никогда не происходит прямого переносафосфатных групп от высокоэнергетического к низкоэнергетическому соединению, а также от высокоэнергетического донора фосфатной группы к высокоэнергетическому акцептору или от низкоэнергетического к низкоэнергетическому. Почти все реакции переноса фосфатных групп идут с участием АТФ ↔ АДФ. Таким образом, функция системы АТФ↔АДФ состоит именно в том, что она служит посредником–переносчиком фосфатных групп от высокоэнергетических к низкоэнергетическим фосфатам по схеме:
Они легко гидролизуются с выделением энергии. В клетке никогда не происходит прямого переносафосфатных групп от высокоэнергетического к низкоэнергетическому соединению, а также от высокоэнергетического донора фосфатной группы к высокоэнергетическому акцептору или от низкоэнергетического к низкоэнергетическому. Почти все реакции переноса фосфатных групп идут с участием АТФ ↔ АДФ. Таким образом, функция системы АТФ↔АДФ состоит именно в том, что она служит посредником–переносчиком фосфатных групп от высокоэнергетических к низкоэнергетическим фосфатам по схеме:
Фосфоенолпируват +АДФ → АТФ + пируват (фермент — пируваткиназа)
Глюкоза + АТФ → АДФ + глюкозо-6-фосфат (фермент – гексокиназа или глюкокиназа).
Процесс синтеза АТФ из АДФ называется фосфорилированием. Выделяют субстратное и окислительное фосфорилирование.При субстратном фосфорилировании для синтеза АТФ используется энергия непосредственно от окисляемого субстрата (например, отдельные реакции гликолиза). Если на синтез АТФ используется энергия, выделяемая при транспорте е — в дыхательной (электрон-транспортной) цепи, то такое фосфорилирование называется окислительным.
Источник



