
Научная электронная библиотека

§ 3.1.4. Строение клетки
Размеры клетки широко варьируют от 0,1 мкм (некоторые бактерии) до 155 мм (яйцо страуса). У всех клеток, независимо от их формы, размеров, функциональной нагрузки обнаруживается сходное строение (рис. 3.13).
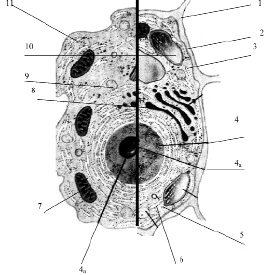
Рис. 3.13. Схема строения живой клетки: 1 – оболочка; 2 – мембрана; 3 – цитоплазма; 4 – ядро; 4а – ядрышко; 5 – рибосомы; 6 – эндоплазматическая сеть (ЭПС); 7 – митохондрии; 8 – комплекс гольджи; 9 – лизосомы; 10 – пластиды; 11 – клеточные включения
Снаружи клетка одета мембраной. Внутренняя часть клетки содержит многочисленные органоиды – структурные образования клетки, выполняющие определенные функции жизнедеятельности клетки.
1. Оболочка. Присутствует только у растительных клеток. Состоит из волокон целлюлозы. Функции оболочки: защита клетки от внешних повреждений, придает стабильную форму клетки, эластичность растительным тканям.
Повреждение наружной оболочки приводит к гибели клетки (цитолиз).
2. Мембрана. Тончайшая структура (75 Ǻ), состоит из двойного слоя молекул липидов и одного слоя белков. Такая структура обеспечивает уникальную эластичность и прочность мембране
 участие в обмене веществ. Эта функция связана с избирательной проницаемостью в клетку определенных веществ и выведение из нее продуктов обмена. В процессе питания в клетку могут проникать определенные растворы веществ (пиноцитоз) и твердые частицы (фагоцитоз).
участие в обмене веществ. Эта функция связана с избирательной проницаемостью в клетку определенных веществ и выведение из нее продуктов обмена. В процессе питания в клетку могут проникать определенные растворы веществ (пиноцитоз) и твердые частицы (фагоцитоз).
Явление фагоцитоза – поглощение клеткой твердых частиц – впервые было описано русским врачом Мечниковым. Фагоцитарная особенность лежит в основе процесса иммунитета. Особенно развита у лейкоцитов, клеток костного мозга, лимфатических узлов, селезенки, надпочечников и гипофиза.
Пиноцитоз – поглощение клеткой растворов – состоит в том, что мельчайшие пузырьки жидкости втягиваются через образующуюся воронку, проникают через мембрану и усваиваются клеткой.
3. Цитоплазма – внутренняя среда клетки. Представляет собой гелеобразную жидкость (коллоидная система), состоит на 80 % из воды, в которой растворены белки, липиды, углеводы, неорганические вещества. Цитоплазма живой клетки находится в постоянном движении (циклоз).
 транспортировка питательных веществ и утилизация продуктов обмена клетки;
транспортировка питательных веществ и утилизация продуктов обмена клетки;
 буферность цитоплазмы (постоянство физико-химических свойств) обеспечивает гомеостаз клетки, поддерживает постоянные нужные параметры жизнедеятельности;
буферность цитоплазмы (постоянство физико-химических свойств) обеспечивает гомеостаз клетки, поддерживает постоянные нужные параметры жизнедеятельности;
 поддержание тургора (упругость) клетки;
поддержание тургора (упругость) клетки;
 все биохимические реакции происходят только в водных растворах, что обеспечивается в среде цитоплазмы.
все биохимические реакции происходят только в водных растворах, что обеспечивается в среде цитоплазмы.
4. Ядро – обязательный органоид эукариотических клеток. Впервые было исследовано и описано Р. Броуном в 1831 г. В молодых клетках расположено в центре клетки, в старых – смещается в сторону. Снаружи ядро окружено мембраной с крупными порами, способными пропускать крупные макромолекулы. Внутри ядро заполнено клеточным соком – кариоплазмой, основная часть ядра заполнена хроматином – ядерным веществом, содержащим ДНК и белок. Перед делением хроматин образует палочковидные хромосомы. Причём, хромосомы одинакового строения (но содержащие разные ДНК!) образуют пары, зрительно воспринимаемые как одно целое (рис. 3.14).

Рис. 3.14. Хромосомный набор человеческой клетки перед началом деления
Структурирование всех хромосом в пары свидетельствует о том, что число хромосом – чётное. Поэтому, его часто обозначают 2n, где n – количество хромосомных пар, а соответствующий набор хромосом называют диплоидным. Например, у голубей n = 40 (80 хромосом), у мухи n = 6 (12 хромосом), у собаки n = 39 (78 хромосом), у аскариды n = 1 (2 хромосомы). У человека n = 23 (46 хромосом). Однако, в половых клетках число хромосом в два раза меньше. Поэтому набор хромосом в половых клетках называется гаплоидным. Клетки, не являющиеся половыми называются соматическими. Иногда клетки с гаплоидным набором хромосом называют гаплоидными клетками, а с диплоидным набором хромосом – диплоидными клетками.
При слиянии двух родительских гаплоидных половых клеток образуется диплоидная клетка, дающая начало новому организму с набором генов отца и матери
Совокупность всех хромосом ядра (а значит и генов) клетки называется генотип. Именно генотип определяет все внешние и внутренние признаки конкретного организма.
В соматических клетках 44 Х-образные хромосомы (22 пары) у женщин и мужчин идентичны (сходны по строению), их называют аутосомами. А 23-я пара имеет конфигурацию ХХ – у женщин и ХY – у мужчин. Эти пары хромосом именуются половыми хромосомами.
В половых клетках 22 хромосомы также одинаковые у яйцеклеток и у сперматозоидов, а 23-я хромосома конфигурации Х – у яйцеклетки и Х или Y – у сперматозоидов. Поэтому при слиянии половых клеток и образовании пар хромосом, 23-я пара будет ( <ХY>или <ХХ>) определять пол будущего ребенка.
Необходимо помнить, что хотя в соматических клетках набор хромосом диплоидный (2n), однако, перед началом деления клеток происходит репликация ДНК, то есть, удвоение их количества, а, значит, и удвоение
количества хромосом. Поэтому перед началом деления соматической клетки в ней насчитывается 4n хромосом (рис. 16). Она становится тетраплоидной.
– хранение генетической информации;
– контроль за всеми процессами, происходящими в клетке: делением, дыханием, питанием и др.
4а. Ядрышко – структура, содержащаяся в ядре. Ядро может содержат 1, 2 или более ядрышек. Функция ядрышка – формирование рибосом.
Следует отметить, что не все клетки имеют оформленное ядро. Клетки, имеющие ядро называются эукариотическими или эукариотами. Клетки, не имеющие ядра, называются прокариотическими или прокариотами. Функции ядра у прокариот несёт одна нить ДНК (именуется хромосома), в которой хранится вся генетическая информация. К прокариотам относятся бактерии и сине-зеленые водоросли. Как правило, у прокариотов отсутствуют и некоторые другие органоиды. Размеры прокариотических клеток меньше, чем размеры эукариот.
5. Рибосомы – самые мелкие органоиды клетки. Были обнаружены в 1954 г. Французским ученым Паладом. Рибосомы были обнаружены в цитоплазме, а также на гранулярной ЭПС и в ядре.
Функция рибосом: обеспечение биосинтеза белка.
6. Эндоплазматическая сеть. Представляет собой каналы и полости, ограниченные мембраной. Различают две разновидности ЭПС: гранулярная ЭПС и агранулярная ЭПС. Гранулярная ЭПС морфологически отличается от агранулярной наличием на ее поверхности многочисленных рибосом (на агранулярной ЭПС рибосомы отсутствуют).
Функции эндоплазматической сети:
– участие в синтезе органических веществ: на гранулярной ЭПС синтезируются белки, на агранулярной – липиды и углеводы;
– транспортировка продуктов синтеза ко всем частям клетки.
Несложно уяснить, что гранулярная ЭПС характерна для клеток, синтезирующих белки (например клетки желез внутренней секреции), агранулярная ЭПС характерна для клеток-производителей углеводов и липидов (например клетки жировой ткани).
7. Митохондрии – крупные органоиды, состоящие из двойного слоя мембран: наружная – гладкая, внутренняя образует многочисленные гребнеобразные складки – кристы. Внутри митохондрии заполнены жидкостью (матрикс).
Функции митохондрий: основная функция митохондрий – обеспечение клетки энергией. Этот процесс происходит за счет синтеза аденозинтрифосфорной кислоты (АТФ) (рис. 3.15), в которой фрагмент

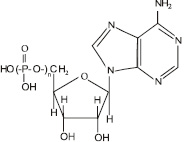
Рис. 3.15. Структурная формула аденозинфосфорных кислот. Для аденозинтрифосфорной кислоты n = 3, для аденозиндифосфорной кислоты n = 2, для аденозинмонофосфорной кислоты n = 1
При взаимодействии молекулы аденозинтрифосфорной кислоты с водой отщепляется один остаток фосфорной кислоты, в результате чего образуется аденозиндифосфорная кислота – АДФ и выделяется огромное количество энергии:
АТФ + Н2О = АДФ + Н3РО4 + 10 000 калорий.
Впоследствии от АДФ может отщепляться еще один остаток фосфорной кислоты, образуя АМФ – аденозинмонофосфорную кислоту.
АДФ + Н2О = АМФ + Н3РО4 + 10 000 калорий[37].
Освободившаяся энергия используется для жизнедеятельности клетки (КПД процесса превышает 80 %!).
Наряду с распадом АТФ и выделением энергии в клетке постоянно происходит синтез АТФ и накопление энергии (обратные реакции).
Количество митохондрий в клетке зависит от потребности последней в энергии. Так, в клетках кожи человека находится в среднем 5–6 митохондрий, в клетках мышц – до 1000, в клетках печени – до 2500!
8. Комплекс Гольджи. Итальянский ученый Гольджи обнаружил и описал структуру клетки, напоминающую стопки мембран, цистерны, пузырьки и трубочки. Расположена эта система чаще всего возле ядра.
Функции комплекса Гольджи: в полостях комплекса накапливаются всевозможные продукты обмена клетки, которые по каким-либо причинам не вывелись наружу. В последствии эти продукты могут быть использованы клеткой для процессов жизнедеятельности. Из пузырьков и цистерночек комплекса Гольджи в растительных клетках образуются вакуоли, заполненные клеточным соком.
9. Лизосомы – мелкие органоиды. Представляют собой пузырьки, окруженные мембраной. Внутри лизосомы заполнены пищеварительными ферментами (обнаружено 12 ферментов), которые расщепляют и переваривают крупные макромолекулы (белки, полисахариды, нуклеиновые кислоты).
Функции лизосом: растворение и переваривание макромолекул. Лизосомы участвуют в фагоцитозе. Понятно, что основная функция по перевариванию поступающих в клетку частиц принадлежит лизосомам.
10. Пластиды. Эти органоиды характерны только для растительных клеток. Форма напоминает двояковыпуклую линзу. Структура пластид напоминает таковую у митохондрий: двойной слой мембраны. Наружная – гладкая, внутренняя образует складки, называемые тилакоидами. На тилакоидах происходит основной жизненно важный для всех зеленых растений процесс – фотосинтез:

Пластиды бывают трех типов:
1) Хлоропласты – зеленые пластиды. Их цвет обусловлен наличием хлорофилла. Хлорофилл – основное вещество хлоропластов (имеет зеленый цвет). Только благодаря хлорофиллу возможен процесс фотосинтеза (см. раздел 4.2). Хлоропласты придают зеленый цвет растительным организмам.
2) Хромопласты – пластиды, имеющие различные окраски: от ярко-желтого до пурпурно-багряного. Наличие различных пигментов окрашивают плоды, цветки и осенние листья растений в соответствующие цвета. Этот факт особенно важен для привлечения насекомых к цветкам, как природный индикатор созревания плодов и др.
3) Лейкопласты – бесцветные пластиды, в которых происходит накопление запасных питательных веществ (например, крахмала).
Некоторые виды пластид могут переходить друг в друга: например, переход хлоропластов в хромопласты: созревание томатов, яблок, вишни, и т. д.; изменение окраски листьев в осенний период времени. Лейкопласты могут переходить в хлоропласты: позеленение картофеля на свету. Это доказывает общность происхождения пластид.
11. Клеточные включения. Вакуоли. Это непостоянные и необязательные составляющие клетки. Они могут появляться и исчезать в течение всей жизни клетки. К ним относятся капли жира, зерна крахмала и гликогена, кристаллы щавелево-кислого кальция и др. Жидкие продукты обмена называются клеточным соком и накапливаются они в вакуолях. В клеточном соке растворены сахара, минеральные соли, пигменты и т. д. Чем старше клетка, тем больше клеточного сока накапливает клетка. Молодые клетки практически не содержат вакуолей.
Помимо перечисленного некоторые специализированные клетки обладают специальными органоидами. К ним относятся:
– реснички и жгутики, представляющие собой выросты мембраны клетки, осуществляющие движения клетки. Они имеются у одноклеточных организмов и многоклеточных (кишечный эпителий, сперматозоиды, эпителий дыхательных путей);
– миофибриллы – тонкие нити мышечных клеток, участвующие в сокращении мышц;
– нейрофибриллы – органоиды, характерные для нервных клеток и участвующие в проведении нервных импульсов. Кроме того, в состав клеток входят центриоли – две (иногда более) цилиндрические структуры диаметром около 0,1 мкм и длиной 0,3 мкм. Место расположения центриолей в период между делениями клетки считается серединой клеточного центра. При делении клетки центриоли расходятся в противоположные стороны – к полюсам, определяя ориентацию веретена деления (рис. 16).
Следует иметь в виду, что, хотя животные и растительные клетки имеют много общего, но между ними существуют и серьёзные различия (табл. 3.1).
Более общая классификация клеток представлена на рис. 3.16.
Одно из основных отличий бактерий от архей, состоит в химическом составе мембраны. Бактерии отделены от внешней среды двойным слоем липидов (жиров и жироподобных веществ). Мембраны архей состоят из терпеновых спиртов.
Источник
Прокариоты. Строение, форма клеток, размножение, питание
» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»>
Прокариоты
- 2.3.1. Строение бактерий
- 2.3.2. Форма клеток
- 2.3.3. Размножение
- 2.3.4. Питание
- 2.3.5. Рост популяции бактерий
К царству прокариот относятся организмы, которых обычно называют бактериями. Это — наидревнейшая группа, появившаяся примерно 3,5 млрд. лет назад; к тому же это и мельчайшие организмы, обладающие клеточной структурой. Свойства прокариот суммированы в табл. 2.2. Как правило, прокариоты представлены одиночными клетками, хотя сине-зеленые водоросли (цианобактерии, Cyanobacteria) могут образовывать цепочки клеток, называемые нитями.
Некоторые бактерии прилипают друг к другу, образуя характерные скопления, напоминающие гроздья винограда (рис. 2.10), однако объединившиеся клетки остаются абсолютно независимыми друг от друга. Индивидуальную бактериальную клетку можно увидеть только с помощью микроскопа, почему их и называют микроорганизмами. Наука, изучающая бактерий — бактериология — составляет важную ветвь микробиологии.

Бактерии различаются по своим размерам: их длина колеблется от 0,1 до 10 мкм, а диаметр в среднем составляет — 1 мкм. Таким образом, в бактериальной клетке достаточно места, чтобы поперек нее уместилось 200 молекул глобулярных белков среднего размера (5 нм в диаметре).
Поскольку такие молекулы способны диффундировать примерно на расстояние 60 мкм в секунду, никаких специальных механизмов транспорта этим организмам не нужно.
Бактерий можно обнаружить повсюду: в почве, и в пыли, в воде и в воздухе, внутри и на поверхности животных и растений. Некоторые бактерии поселяются в горячих источниках с температурой 78 °С или выше. Другие способны выжить при очень низких температурах и даже пережить определенные периоды замораживания во льду. Встречаются бактерии и в глубоких расселинах на дне океана при очень высоком давлении и температуре 360 °С. С них начинаются уникальные пищевые цепи в этих областях океана.
Число бактерий невообразимо велико; установлено, что в одном грамме плодородной почвы содержится 2,5 млрд. бактерий; в 1 см 3 свежего молока их содержание может превышать 3 млрд. Вместе с грибами бактерии имеют жизненно важное значение для всех других организмов, поскольку, разрушая в результате своей жизнедеятельности органические вещества, они обеспечивают циркуляцию биогенных элементов в природе. Кроме того, они приобретают все более важное значение в жизни человека, и не только потому, что некоторые из них являются возбудителями различных болезней, но и потому, что в силу разнообразия протекающих в них биохимических реакций они могут использоваться во многих биотехнологических процессах.
2.3.1. Строение бактерий
На рис. 2.5 показано строение обобщенной бактерии — типичной прокариотической клетки. На рис. 2.6, А–Г изображена широко известная палочковидная бактерия Escherichia coli. Обычно она совершенно безвредна. Ее наличие в воде может использоваться в качестве очень надежного показателя загрязнения воды фекалиями. Из всех бактерий E. coli изучена лучше всего. Кроме того, это одна из бактерий, генетическая карта которых установлена полностью. Обратите внимание, что у E. coli намного меньше видимых внутриклеточных структур, чем в эукариотической клетке (рис. 5.10 и 5.11). На рис. 2.7 показана другая палочковидная бактерия, у которой в отличие от E. coli имеется жгутик.
Клеточная стенка
Клеточная стенка бактерий — структура довольно прочная и позволяет клетке сохранять свою форму; это обусловлено наличием в ней муреина — молекулы, построенной из параллельных полисахаридных цепей, перекрестно связанных через регулярные интервалы короткими цепями аминокислот. Таким образом, каждая клетка окружена как бы сетчатым мешком, представляющим на деле одну огромную молекулу. Клеточная стенка предохраняет клетку от разрыва при поступлении в нее воды (например, в результате осмоса). Ионы воды и малые молекулы попадают в клетку через мельчайшие поры в клеточной стенке.
В 1884 г. датский биолог Кристиан Грам разработал метод окрашивания, с помощью которого было установлено, что бактерии подразделяются на две естественные группы, что, как теперь стало известно, обусловлено различиями в строении их клеточной стенки. Одни бактерии, окрашивающиеся по Граму, получили название грамположительных, другие, не окрашивающиеся, — грамотрицательных.




У грамположительных бактерий, таких как Staphylococcus, Bacillus и Lactobacillus в муреиновую сетку встроены другие компоненты, в основном полисахариды и белки, что делает клеточную стенку сравнительно толстой. У грамотрицательных бактерий, таких как Salmonella, E.coli и Azotobacter, клеточная стенка тоньше и имеет более сложное строение (рис. 2.8). Муреиновый слой у этих бактерий снаружи покрыт гладким тонким мембраноподобным слоем липидов и полисахаридов, защищающим клетки от лизоцима — антибактериального фермента, содержащегося в слезах, слюне и других биологических жидкостях, а также в белке куриного яйца.
Лизоцим расщепляет полисахаридный каркас муреина, что приводит к продырявливанию клеточной стенки и лизису клетки, т. е. к ее осмотическому набуханию и разрыву. Липидно-полисахаридный слой обусловливает также устойчивость грамотрицательных бактерий к пенициллину. Этот антибиотик блокирует образование перекрестных сшивок в муреине растущих грамположительных бактерий, что делает их клетки более чувствительными к осмотическому шоку.
Плазматическая мембрана, мезосомы и фотосинтетические мембраны
Как и у всех других организмов, живое вещество бактериальной клетки окружено полупроницаемой мембраной. По строению и функциям плазматическая мембрана бактериальных клеток не отличается от плазматических мембран эукариотических клеток (разд. 5.9). Она служит также местом локализации дыхательных ферментов, а у некоторых бактерий она образует мезосомы и(или) фотосинтетические мембраны.
Мезосомы— складчатые структуры, представляющие собой впячивания плазматической мембраны клетки (рис. 2.5). Во время клеточного деления мезосомы, по-видимому, ассоциируются с ДНК, что обеспечивает разделение двух дочерних молекул ДНК после репликации и способствует образованию перегородки между дочерними клетками.
У фотосинтезирующих бактерий в мешковидных, трубчатых или пластинчатых впячиваниях плазматической мембраны содержатся фотосинтетические пигменты (в том числе обязательно бактериохлорофилл). Сходные мембранные образования участвуют и в фиксации азота.
Генетический материал (бактериальная «хромосома»)
Бактериальная ДНК представляет собой одиночную кольцевую молекулу длиной около 1 мм (т. е. она значительно длиннее, чем сама клетка), состоящую примерно из 5 млн. пар оснований.
Суммарное содержание ДНК (геном), а следовательно, и количество закодированной в ней информации, в бактериальной клетке значительно меньше, чем в эукариотической: в типичном случае у бактерии ДНК содержит несколько тысяч генов, что в 500 раз меньше, чем в клетке человека (см. также табл. 2.2 и рис. 2.5).
Рибосомы
Рибосомы служат местом синтеза белков (см. табл. 2.2 и рис. 5.5).
Капсулы
У некоторых бактерий слизистые или клейкие секреты образуют капсулы; капсулы хорошо видны после негативного контрастирования (когда окрашивают не препарат, а фон). Иногда эти секреты служат для формирования колоний из одиночных бактерий. С помощью секретов бактерии приобретают способность прилипать к различным поверхностям, таким как зубы, частицы ила или скалы. Кроме того, капсулы обеспечивают дополнительную защиту для бактериальной клетки. Так, например, капсулированные штаммы пневмококков свободно размножаются в организме человека, вызывая воспаление легких, тогда как некапсулированные штаммы легко атакуются и разрушаются фагоцитами и поэтому совершенно безвредны.
Споры
Некоторые бактерии, главным образом относящиеся к родам Clostridiumи Bacillus, образуют эндоспоры (т. е. споры, которые располагаются внутри клеток). Споры представляют собой толстостенные долгоживущие образования, отличающиеся очень высокой устойчивостью, особенно к нагреванию, коротковолновому облучению и высушиванию. Локализация спор в клетке бывает различной и служит важным признаком для идентификации и классификации бактерий (см. рис. 2.10).
Жгутики
Многие бактерии подвижны, что обусловлено наличием у них одного или нескольких жгутиков. Жгутик — это простой полый цилиндр, образуемый одинаковыми белковыми молекулами.
Несмотря на волнистую форму, они довольно жестки (рис. 2.7). Подвижность бактерий достигается вращением основания жгутика; получается, что жгутик как бы ввинчивается в среду, не совершая беспорядочных биений, и таким образом продвигает бактерию за собой. В качестве примеров бактерий, имеющих жгутики, приведем Rhizobium (один жгутик) и Azotobacter (много жгутиков); обе бактерии участвуют в круговороте азота в природе.
Подвижные бактерии могут передвигаться в ответ на определенные раздражители, т. е. они способны к таксису. Аэробные бактерии, например, перемещаются в направлении увеличения концентрации кислорода в среде (проявляют положительный аэротаксис), а подвижные фотосинтезирующие бактерии плывут к свету (проявляют положительный фототаксис).
Жгутики лучше всего видны в электронном микроскопе при использовании метода напыления (рис. 2.7).
Пили
На клеточной стенке некоторых грамотрицательных бактерий видны многочисленные тонкие палочковидные выросты, которые называются пили, или фимбрии (рис. 2.7). Пили короче и тоньше жгутиков и служат для прикрепления к специфическим клеткам или поверхностям. Известны различные типы пилей, но наибольший интерес вызывают F-пили, участвующие в половом размножении (разд. 2.3.3).
Плазмиды
Помимо единственной молекулы ДНК, имеющейся у всех бактерий, у некоторых из них обнаруживается еще одна или более плазмид (рис. 2.9).

Плазмида — это небольшая кольцевая молекула дополнительной ДНК, способная к саморепликации. Плазмида несет в себе всего несколько генов, обусловливающих повышенную выживаемость клеток. Некоторые плазмиды делают клетку устойчивой к антибиотикам.
Например, в клетках некоторых стафилококков содержится плазмида, несущая ген пенициллиназы — фермента, расщепляющего пенициллин.
В результате клетка оказывается устойчивой к пенициллину. Распространение таких генов при конъюгации находит важное применение в медицине. Известны и другие плазмидные гены, в частности гены,
- 1) придающие устойчивость к дезинфицирующим средствам;
- 2) вызывающие различные болезни;
- 3) отвечающие за сбраживание молока молочнокислыми бактериями при сыроварении;
- 4) придающие способность использовать в качестве пищи такие сложные химические вещества, как углеводороды, и поэтому потенциально пригодные для использования в борьбе с нефтяными пятнами и для образования белка из нефтепродуктов.
2.3.2. Форма клеток
Форма бактериальной клетки является одним из важнейших систематических признаков. Четыре основных типа клеток приведены на рис. 2.10. На этом же рисунке указаны как полезные, так и болезнетворные бактерии.

2.3.3. Размножение
Индивидуальный рост и бесполое размножение
Соотношение поверхность/объем у бактериальных клеток очень велико, что способствует быстрому поглощению питательных веществ из окружающей среды за счет диффузии и активного транспорта. Поэтому в благоприятных условиях бактерии способны расти очень быстро. Рост бактериальных клеток в большой степени зависит от таких факторов среды, как температура, наличие питательных веществ, pH среды и концентрация ионов. Кроме того, облигатным аэробам необходим кислород, а облигатным анаэробам необходимо, чтобы его не было.
Достигнув определенных размеров, диктуемых соотношением объемов ядра и цитоплазмы, бактерии переходят к бесполому размножению путем простого деления, т. е. путем деления на две идентичные дочерние клетки (рис. 2.11).

Клеточному делению предшествует репликация ДНК, причем до тех пор, пока процесс репликации не завершится, мезосомы могут удерживать ДНК в определенном положении (рис. 2.5 и 2.6, В). Мезосомы могут прикрепляться и к новым перегородкам, образующимся между дочерними клетками, участвуя каким-то образом в синтезе материала клеточной стенки. У самых быстрорастущих бактерий деление происходит через каждые 20 мин.
Половое размножение
В 1946 г. у бактерий было обнаружено половое размножение, но в самой примитивной форме. Гамет в данном случае не образуется, однако наиважнейшее событие полового размножения, а именно обмен генетическим материалом, происходит и в этом случае. Этот процесс называется генетической рекомбинацией. Генетическая рекомбинация впервые была обнаружена при изучении E.coli. В норме при наличии в среде достаточного количества глюкозы и неорганических солей E.coli сама синтезирует все необходимые ей аминокислоты. В результате облучения этих бактерий у них иногда возникают случайные мутации. Были выделены два типа мутантов: один, не способный синтезировать биотин (витамин) и аминокислоту метионин, и другой — не способный синтезировать аминокислоты треонин и лейцин. В среду, не содержавшую всех четырех факторов роста, помещали по 10 8 клеток каждого мутантного штамма. Теоретически клетки не должны были расти на этой среде. Однако все же было получено несколько сотен колоний (каждая колония возникает из одной исходной клетки), причем оказалось, что в таких клетках имеются все гены, необходимые для образования этих четырех факторов роста. Следовательно, в клетках каким-то образом произошел обмен генетической информацией, но выделить вещество, ответственное за этот процесс, в то время не удалось. В конце концов было установлено (при помощи электронного микроскопа), что клетки E.coli могут непосредственно контактировать друг с другом, т. е. у них может происходит конъюгация (рис. 2.12).

Таким образом, при конъюгации происходит перенос ДНК между клетками в результате прямого контакта. Одна клетка в этом случае служит донором («мужская» клетка), другая — реципиентом («женская» клетка). Способность клетки служить донором определяется генами, содержащимися в особой плазмиде, называемой половым фактором или F-фактором (F от англ. fertility — плодовитость). В этих генах закодирован белок специфических пилей, называемых F-пилями или половыми пилями. F-пили участвуют в межклеточном контакте при конъюгации. Пили — структуры полые и предполагается, что именно по этим пилям осуществляется перенос ДНК от донора (F + ) к реципиенту (F – ).
Процесс этот показан на рис. 2.13.

Обратите внимание на то, что донорная клетка сохраняет F-фактор, а реципиентная клетка его приобретает и становится F + . Процесс этот протекает медленно, и поэтому прежде чем перенос F-плазмиды завершится, клетка, бывшая изначально F – , успевает реплицироваться один или несколько раз, и в результате в популяции всегда сохраняются F – клетки. F-фактор вызывает особенно большой интерес и потому, что время от времени, примерно в 1 случае из 100 000, он встраивается в молекулу основной ДНК клетки-хозяина. Тогда при конъюгации происходит перенос не только F-фактора, но и всей остальной ДНК. Этот процесс занимает примерно 90 мин, но клетки могут расходиться и раньше, чем произойдет полный обмен ДНК. Такие штаммы постоянно передают всю или большую часть своей ДНК другим клеткам. Эти штаммы называют
Hfr-штаммами (от англ. H — High — высокая, f — frequency — частота, r — recombination — рекомбинация), потому что донорная ДНК таких штаммов рекомбинирует с ДНК реципиента.
2.3.4. Питание
Питание — это процесс приобретения энергии и веществ. Основываясь на природе необходимого источника энергии или источника углерода — наиважнейшего элемента для роста, — живые организмы можно подразделить на несколько групп. Для синтеза органических соединений живые организмы способны использовать только два вида энергии: энергию света и энергию химических связей. Организмы, использующие световую энергию, называются фототрофами, а организмы, использующие только химическую энергию — хемотрофами. Фототрофы осуществляют фотосинтез.
Как уже говорилось, организмы разделяют также на автотрофные и гетеротрофные — в зависимости от того, какой источник углерода они используют: неорганическое соединение (диоксид углерода) или разнообразные органические вещества. Таким образом, можно выделить четыре типа питания (табл. 2.3). Среди бактерий встречаются представители всех четырех типов. Наибольшую группу образуют хемогетеротрофные бактерии.
Хемогетеротрофные бактерии
Бактерии этого типа получают энергию из поступающих с пищей химических соединений.
Они способны использовать огромное множество различных веществ. Среди хемогетеротрофных бактерий можно выделить три основные группы, а именно сапротрофы, мутуалисты и паразиты.
Сапротрофы представлены организмами, извлекающими питательные вещества из мертвого разлагающегося материала. Для разложения органического материала сапротрофы выделяют на него ферменты. Таким образом, переваривание пищи у них происходит вне организма. Образующиеся при этом растворимые продукты поступают в тело сапротрофа и там ассимилируются.
Сапротрофные бактерии и грибы составляют группу редуцентов. Им принадлежит важная роль в разложении органического материала и возврате элементов в природные круговороты. Они образуют гумус из животных и растительных остатков, но при этом они способны и наносить вред, разрушая нужные человеку материалы, особенно пищевые продукты. Значение сапротрофов в биосфере рассматривается в гл. 10.
Мутуализмом (или симбиозом) называют любую форму тесной взаимосвязи между двумя живыми организмами, выгодной для обоих партнеров. Примером бактериального мутуализма может служить Rhizobium — бактерия, способная фиксировать азот и живущая в корневых клубеньках бобовых растений, например гороха и клевера, или Escherichia coli, обитающая в кишечнике человека и, вероятно, поставляющая человеку витамины группы B и K.
Паразитом называют любой организм, живущий внутри тела или на теле другого организма (хозяина), от которого он получает пищу и, как правило, убежище. Хозяевами могут служить представители самых различных видов, причем паразиты наносят ощутимый вред своим хозяевам. Паразиты, вызывающие болезни, называют патогенами. Некоторые из них приведены на рис. 2.10. Одни паразиты, называемые облигатными, могут жить и расти только в живых клетках. Другие, называемые факультативными, заражают хозяина, вызывают его гибель и затем живут на его остатках как сапротрофы. Паразиты отличаются чрезвычайной разборчивостью в пище, поскольку они нуждаются во «вспомогательных факторах роста», которые не способны синтезировать сами, но могут получать только от своих хозяев.
Фотоавтотрофные бактерии
Примерами фотоавтотрофных бактерий могут служить цианобактерии, называемые также сине-зелеными бактериями. Водоросли и растения также являются фотоавтотрофами. Все они осуществляют фотосинтез и используют углекислый газ (CO2) в качестве единственного источника углерода (табл. 2.3).

Процесс фотосинтеза впервые появился у бактерий, возможно именно у цианобактерий. Как мы увидим далее, хлоропласты водорослей и наземных растений представляют собой, по-видимому, потомков некогда свободноживущих фотосинтезирующих бактерий, поселившихся в свое время в гетеротрофных клетках.
Цианобактерии широко распространены в поверхностных водах морей и пресных водоемов. Кроме того, они обнаружены в слизистых подушковидных образованиях на затененных почвах, на скалах, в иле, на древесине и в некоторых живых организмах. Большинство цианобактерий представлены одиночными клетками, хотя некоторые из них объединяются, образуя покрытые слизью нити, например Anabaena и Spirulina. В отличие от большинства бактерий они, подобно водорослям и растениям, способны к фотосинтезу, а, следовательно, и к выделению кислорода из воды. На рис. 2.14 показано строение типичной цианобактерии Anabaena.

Как видно из рисунка, толщу цитоплазмы характерным образом пронизывают фотосинтетические мембраны, на которых располагаются фотосинтетические пигменты. Пигменты представлены хлорофиллом а, похожим на пигмент растений и водорослей, и специфическим сине-зеленым пигментом фикоцианином. Клетки цианобактерий, как правило, крупнее клеток других бактерий. Способность цианобактерий выделять кислород в процессе фотосинтеза, наличие у них фотосинтетических мембран, и хлорофилл а свидетельствуют о том, что цианобактерии вполне могут быть эволюционным звеном между остальными бактериями и эукариотами.
Некоторые цианобактерии, такие как Anabaena, способны фиксировать азот. Иными словами, они способны превращать содержащийся в воздухе газообразный азот в аммиак, который затем может быть использован для синтеза аминокислот, белков и других азотсодержащих органических соединений. Этот процесс происходит в специализированных клетках, называемых гетероцистами, которые образуются при недостатке азота. Гетероцисты экспортируют содержащиеся в них азотистые вещества в соседние клетки в обмен на другие питательные вещества, например углеводы.
Хемоавтотрофные бактерии
Эти организмы чаще называют хемосинтезирующими бактериями. В качестве источника углерода они используют CO 2 (диоксид углерода), но энергию получают в результате химических реакций. Высвобождение необходимой энергии происходит при окислении таких неорганических веществ, как аммиак и нитриты. Некоторые хемоавтотрофные бактерии играют важную роль в круговороте азота, участвуя в процессе, называемом нитрификацией. Процесс нитрификации протекает в две стадии. На первом этапе аммиак окисляется до нитрита, что сопровождается выделением энергии. Этот этап осуществляется такими, например, бактериями, как Nitrosomonas. На втором этапе образовавшийся нитрит окисляется до нитрата с высвобождением дополнительной энергии. Этот этап осуществляется, например, Nitrobacter

2.3.5. Рост популяции бактерий
2.1. Рассмотрим ситуацию, когда одиночная бактериальная клетка помещена в питательную среду и находится в условиях, оптимальных для роста.
Перепишите табл. 2.4 и заполните ее, исходя из предположения, что эта клетка и все ее потомки делятся каждые 20 мин.
На основе данных заполненной вами таблицы постройте графики.
По вертикальной оси отложите число бактерий (кривая А) и десятичный логарифм этого числа (кривая Б), а по горизонтальной оси — время.
Что можно сказать о форме этих кривых?

Когда число клеток увеличивается, как видно из заполненной вами табл. 2.4, говорят о логарифмическом, экспоненциальном или геометрическом росте. В этом случае мы получим экспоненциальный ряд чисел. Это гораздо легче понять, если посмотреть на строку В в табл. 2.4, где число бактерий выражено в виде числа 2, возведенного в соответствующую степень. Показатель степени можно назвать логарифмом или экспонентной числа 2.
Логарифмы, или экспоненты, образуют линейный ряд 0, 1, 2, 3 и т. д., соответствующий числу генераций.
Вернемся к табл. 2.4; вместо чисел, расположенных в строке А, можно записать их логарифмы по основанию 2 следующим образом:

Сравните строки В и Г. Однако обычно пользуются десятичными логарифмами (см. строку Б). В этом случае 1 = 10 0 , 2 = 10 0,3 , 4 = 10 0,6 и т. д. Кривая, полученная на основе данных строки А (табл. 2.4.), называется логарифмической или экспоненциальной кривой. Такую кривую можно преобразовать в прямую, построив график изменения числа клеток во времени. Тогда в идеальных условиях рост бактерий теоретически должен быть экспоненциальным. Сравним эту математическую модель с кривой роста реальной популяции бактерий, изображенной на рис. 2.15. Отчетливо видны четыре фазы роста.

1. Во время лаг-фазы бактерии адаптируются к новой среде обитания, и поэтому рост пока еще не достигает максимальной скорости. В этот период у бактерий могут, например, синтезироваться новые ферменты, необходимые для усвоения тех питательных веществ, которые содержатся в новой среде.
2. Логарифмическая фаза — это фаза, когда бактерии растут с максимальной скоростью, число клеток увеличивается почти экспоненциально, а кривая роста представляет собой практически прямую.
3. В конце концов рост колонии начинает замедляться, и культура входит в стационарную фазу, когда скорость роста равна нулю и когда резко возрастает конкуренция за пищевые ресурсы. Образование новых клеток замедляется, а затем совсем прекращается. Увеличение числа клеток компенсируется одновременной гибелью других клеток, поэтому число жизнеспособных клеток остается постоянным (табл. 2.5). Переход к этой фазе обусловлен действием нескольких факторов: снижением концентрации питательных веществ в среде, накоплением токсичных продуктов метаболизма, а в случае аэробных бактерий и уменьшением содержания кислорода в среде.
4. Во время последней фазы — фазы замедления роста — ускоряется гибель клеток и прекращается их размножение.
Источник



