
- Дизентерия — симптомы и лечение
- Определение болезни. Причины заболевания
- Возбудитель дизентерии
- Эпидемиология
- Симптомы дизентерии
- Дизентерийную палочку по способу питания относят к
- Дизентерийную палочку по способу питания относят к
- Питание бактерий и грибов
- Питание бактерий
- Способы питания живых организмов
- Питание грибов
- Бесцветные серобактерии
- Эубактерии, окисляющие соединения серы: распространение в природе
- Железобактерии: общие сведения
- К сапротрофам относятся
Дизентерия — симптомы и лечение
Что такое дизентерия? Причины возникновения, диагностику и методы лечения разберем в статье доктора Александрова Павла Андреевича, инфекциониста со стажем в 13 лет.

Определение болезни. Причины заболевания
Шигеллёзы, или дизентерия — это группа острых/хронических заболеваний, вызываемых бактериями рода Шигелла, которые поражают желудочно-кишечный тракт (преимущественно дистальный отдел толстой кишки). Манифестные формы (клиническая картина) дизентерии: синдром общей инфекционной интоксикации, синдром поражения желудочно-кишечного тракта (дистальный колит), синдром обезвоживания. При отсутствии адекватного лечения приводит к серьёзным осложнениям и хронизации.
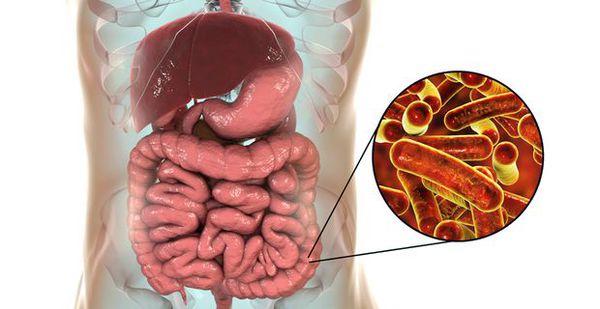
Возбудитель дизентерии
семейство — кишечные бактерии (Enterobacteriaceae)
род — Шигелла (Shigella)
виды — 4 основных:
- Dysenteriae (серовары — шигелла Григорьева-Шига, Штутцера-Шмитца, Ларджа-Сакса);
- Flexneri (Флекснера);
- Boydii (Боудии);
- Sonnei (Зонне).
Являются факультативно-анаэробными (способны существовать как при наличии, так и при отсутствии кислорода), граммотрицательными, неподвижными палочками. Хорошо растут на обычных питательных средах.
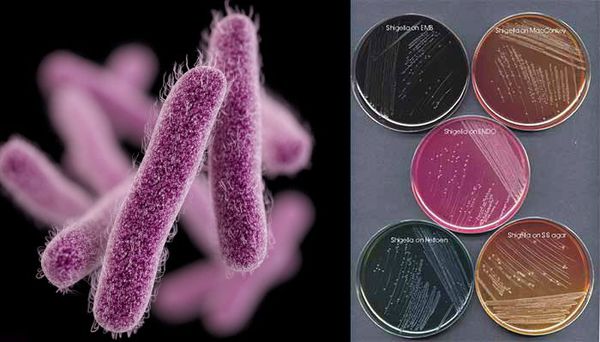
Имеют О-антиген (серологическая специфичность) и К-антиген (оболочечная структура). Клеточная стенка частично состоит из эндотоксина (липополисахарида), высвобождающегося при гибели микроорганизма. Бактерии вида Shigella Dysenteriae серовара Григорьева-Шига продуцируют высокоактивный цитотоксин, который нарушает синтез белка рибосомами клеток кишечного эпителия. Цитотоксин в совокупности с энтеротоксином (усиливающим секрецию жидкости и солей в просвет кишки) и нейротоксином (оказывающим нейротоксическое действие на сплетения Ауэрбаха) образует экзотоксином — токсин, выделяемый в процессе жизнедеятельности шигелл (Цитотоксин + энтеротоксин + нейротоксин = экзотоксин). Также у данной группы микроорганизмов происходит выделение гемолизинов — веществ, разрушающих эндотелий капилляров и вызывающий ишемию в тканях кишечника).
Факторы патогенности шигелл (механизм приспособления бактерий):
- адгезия (прикрепление к слизистой оболочке кишечника);
- инвазия (проникновение в эпителиальные клетки слизистой оболочки толстого кишечника);
- токсинообразование;
- внутриклеточное размножение.
Тинкториальные свойства (особенности микроорганизмов при окрашивании): быстро изменяют чувствительность к антибактериальным препаратам. Характерна высокая выживаемость во внешней среде (в воде, почве и пище при комнатной температуре сохраняются до 14 дней, в канализационных стоках — до 30 дней, при благоприятных условиях могут быть жизнеспособны до 4 месяцев). Способны к размножению в пищевых продуктах. При воздействии УФО (ультрафиолетового облучения) погибают за 10 минут, в 1% феноле — за 30 минут, при кипячении — мгновенно.
Интересная особенность: чем выше ферментативная активность возбудителя дизентерии, тем ниже вирулентность (минимальная доза возбудителя, способная заразить человека) и наоборот. [3] [6]
Эпидемиология
Антропоноз (повсеместное распространение).
Источник инфекции — человек (больной, носитель и лица с субклинической формой заболевания).
Минимальная инфицирующая доза для бактерии Григорьева-Шига — 10 микробных тел в 1 грамме вещества, для Флекснера — 10 2 бактерий, для Зонне — от 10 7 .
Длительное время в РФ превалировали случаи заражения, вызванные шигеллой Флекснера 2а, однако в настоящее время ввиду широкого развития туризма длительного преобладания какой-либо формы нет.
Механизм передачи — фекально-оральный (пищевой, водный, контактно-бытовой), из них преимущественно:
- Григорьева-Шига (контактно-бытовой путь);
- Зонне (с молоком и молочными продуктами);
- Флекснера (водный путь);
- Дизентериа (пищевой путь).
Наибольшую опасность представляют работники питания и водоснабжения. Больные заразны с начала болезни (конца инкубационного периода) и до трёх недель болезни.
Иммунитет непродолжителен и моноспецифичен (вырабатывается только к одному серотипу, который вызвал заболевание). [1] [4] [5]
Симптомы дизентерии
Инкубационный период — от 1 до 7 дней.
Начало заболевания острое (развитие основных синдромов наблюдается в первые сутки заболевания), характерна лихорадка постоянного типа.
- общей инфекционной интоксикации — в первую очередь;
- поражения желудочно-кишечного тракта (ЖКТ) — колит;
- дегидратационный — обезвоживание организма.
Источник
Дизентерийную палочку по способу питания относят к
Дизентерийную палочку по способу питания относят к
А) сапрофитам
Б) паразитам
В) хемотрофам
Г) автотрофам
2.Все бактерии способны к
А) быстрому размножению
Б) накоплению в их клетках ядовитых веществ
В) спорообразованию в неблагоприятных условиях
Г) развитию заболеваний при попадании в организм животного
3.Царство Бактерий объединяет организмы, тело которых состоит из
А) одной клетки с оформленным ядром и многими органоидами
Б) одной и множества ядерных клеток, но без оболочки из клетчатки и хлоропластов
В) одной или множества клеток с оболочкой из хитинообразного вещества, но без хлоропластов
Г) одной безъядерной клетки, не имеющей органоидов
4.К царству Бактерий относится
А) малярийный паразит
Б) хлорелла
В) кишечная палочка
Г) инфузория-туфелька
5.Хемосинтезирующими бактериями являются
А) железобактерии
Б) бактерии брожения
В) молочнокислые бактерии
Г) сине-зеленые (цианобактерии)
6.Группа бактерий, живущих в содружестве с другими организмами, -это
А) паразиты
Б) симбионты
В) консументы
Г) сапрофиты
7.Укажите главный признак бактерий
А) ядерное вещество не отделено от цитоплазмы
Б) отсутствует оболочка
В) нет рибосом
Г) наличие митохондрий
8.Клубеньковые бактерии вступают в симбиоз с растениями семейства
А) розоцветных
Б) пасленовых
В) крестоцветных
Г) бобовых
9.Назовите признак, характерный только для царства Бактерий
А) имеют клеточное строение
Б) дышат, питаются, размножаются
В) в клетках есть оформленное ядро
Г) в клетках отсутствует оформленное ядро
10.Наследственное вещество находится в цитоплазме клеток у
А) дизентерийной амебы
Б) малярийного паразита
В) кишечной палочки
Г) эвглены зеленой
Ответы: 1-Б, 2-В, 3-Г, 4-В, 5-А, 6-Б, 7-А, 8-Г, 9-Г, 10-В
Питание бактерий и грибов
Растения в природе выполняют функцию производителей органического вещества. Животные играют роль потребителей, питающихся органическими веществами. Бактерии и грибы — разрушители тел отмерших организмов.
Они разлагают мертвые тела до минеральных веществ, которые поступают в почву и вновь поглощаются растениями.
Питание бактерий
Большинство бактерий питаются готовыми органическими веществами. Разлагая их, они строят свои клетки. Такие бактерии называют гетеротрофами. Некоторые гетеротрофные бактерии питаются органическими веществами, содержащимися в мертвых телах, выделениях растений и животных.
Такие бактерии обитают в воде, почве, на коже, в кишечнике животных. Разлагая органические вещества, они возвращают в почву минеральные соли, а в атмосферу углекислый газ, необходимый для фотосинтеза.
Бактерии-паразиты поселяются в тканях человека, животных, растений и вызывают различные бактериальные болезни.
К ним относятся туберкулезная палочка, дизентерийная палочка, возбудители ангины, пневмонии.
На корнях фасоли, бобов, люпина .можно увидеть маленькие клубеньки, размером чуть больше булавочной головки. В них находятся очень полезные для растений клубеньковые бактерии.
Способы питания живых организмов
Они поглощают азот из воздуха и преобразуют его в доступные для растений азотные соединения. Клубеньковые бактерии снабжают растение-хозяина азотом, а сами извлекают из корней растения необходимые для жизнедеятельности вещества удобрением.
Когда растение отмирает, соединения азота остаются в почве и повышают ее плодородие. Подобные взаимовыгодные отношения между различными организмами называют симбиозом (от греч. «symbosis» — совместная жизнь).

Питание грибов
Грибы, так же, как и большинство бактерий, гетеротрофы.
Грибы отличаются друг от друга по типу питания.
Многие грибы обитают в почве. Они питаются, разрушая органические останки растений, животных, плодовые тела грибов. Некоторые грибы питаются органическими веществами, содержащимися в пищевых продуктах.
Другие паразитируют, поселяясь на живых организмах (растениях или животных), и питаются за их счет. Так, некоторые грибы могут поселиться на теле человека, вызывая различные заболевания, которые так и называют грибковыми.
Некоторые грибы получают органические вещества из корней деревьев.
При этом мицелии гриба вступает в тесный контакт с корнями деревьев. Гифы мицелия плотно оплетают корень и даже проникают внутрь ого клеток. Грибы поглощают из почвы воду и растворенные минеральные вещества, которые по их гифам поступают в корни деревьев. Таким образом, мицелий заменяет деревьям корневые волоски. А из корней деревьев гриб получает органические вещества, созданные растением и необходимые грибу для питания.
В результате образуется грибо-корень, или микориза (от греч. mykes — гриб и rhiza — корень).
Для питания грибы используют сложные органические вещества, которые не могут проникать в клетки через клеточные оболочки. Пищеварительный сок, который у животных выделяется в кишечник, у грибов выделяется наружу.
С помощью его ферментов происходит расщепление сложных органических веществ на более простые, которые всасываются всей поверхностью гриба.

Бесцветные серобактерии
Бесцветные серобактерии очень напоминают цианобактерии , являясь как бы их непигментированными аналогами. На основании морфологических признаков делятся на две группы: одна представлена одноклеточными формами (роды Achromatium , Macromonas и др.), в составе другой объединены нитчатые организмы (роды Beggiatoa , Thiothrix , Thioploca ).
Одноклеточные бесцветные серобактерии — подвижные или неподвижные, различающиеся размерами, формами. Нитчатые организмы представлены также неподвижными или способными к скользящему движению видами (рис. 2 , 1).

Единственный общий признак группы — способность откладывать серу в периплазматическом пространстве клеток. Вопрос о значении, которое имеет окисление восстановленных соединений серы для этой группы бактерий, имеет длинную историю.
С.Н.Виноградский, наблюдая в 1887-1889 гг. в клетках Beggiatoa при выращивании на среде с H2S отложение гранул серы и их последующее исчезновение после исчерпания сероводорода из среды, пришел к выводу, что энергия, освобождающаяся при окислении H2S до S0 и затем до SO4 с участием О2, используется этим организмом для ассимиляции СО2.
Таким образом, работая с Beggiatoa , Виноградский сформулировал положение о принципиально новом способе существования организмов -хемолитоавтотрофии . Однако позднее выяснилось, что культуры, с которыми работал Виноградский, были нечистыми. И до сих пор большинство представителей этой группы не выделены в виде чистых культур, что затрудняет изучение их физиологии. Для некоторых бесцветных серобактерий, в том числе и для Beggiatoa , были получены данные в пользу того, что окисление H2S может быть связано с получением клеткой энергии.
В то же время показано, что важная физиологическая особенность бесцветных серобактерий — образование ими значительных количеств перекиси водорода .
Более 80-90% потребленного клетками в процессе дыхания О2 восстанавливается лишь до Н2О2. Накоплению в клетках перекиси водорода способствует низкая каталазная активность, обнаруженная у этих организмов. Была выявлена определенная связь между окислением H2S и кислородным метаболизмом бесцветных серобактерий. Оказалось, что окисление соединений серы используется этими организмами для удаления Н2О2. Отложение молекулярной серы является, таким образом, результатом окисления сульфидов среды перекисью водорода, образующейся в клетке.
Перекисный механизм окисления восстановленных соединений серы исключает возможность использования организмами энергии этого процесса.
Вопрос о способности бесцветных серобактерий существовать автотрофно также пока не доказан: чистые культуры могут расти только в присутствии органических соединений; не обнаружено типичных для эубактерий механизмов автотрофной ассимиляции СО2.
Все это заставляет склоняться в пользу того, что бесцветные серобактерии могут существовать только хемогетеротрофно . В микроаэробных условиях некоторые штаммы Beggiatoa обнаруживают способность к азотфиксации.
Эубактерии, окисляющие соединения серы: распространение в природе
Окисление неорганических восстановленных соединений серы с помощью фототрофных и хемотрофных эубактерий является одним из звеньев круговорота серы в природе.
В первом случае процесс протекает в анаэробных условиях, во втором — в аэробных . Хемотрофы, окисляющие серу, обитают в морских и пресных водах, содержащих О2, в аэробных слоях почв разного типа.
Поскольку эта группа объединяет организмы с разными физиологическими свойствами, ее представителей можно обнаружить в кислых горячих серных источниках, кислых шахтных водах, в водоемах со щелочной средой и высокой концентрацией NaCl.
Хемолитоавтотрофные серобактерии обнаружены на глубине 2600-6000 м в местах, где на поверхность дна океана из недр земной коры выходят горячие источники. Вода источников, называемая гидротермальной жидкостью, имеет температуру до 350 градусов по С, не содержит совсем О2 и NO3, но обогащена H2S, CO2 и NH4+.
На дне океана гидротермальная жидкость смешивается с окружающей морской водой, имеющей температуру 2 градуса по С, которая, наоборот, не содержит H2S и характеризуется достаточно высокими уровнями О2 и NO3-.
Эти области отличаются также высоким давлением и полным отсутствием света.
К удивлению исследователей, вокруг выходов гидротермальной жидкости были обнаружены плотные скопления необычных беспозвоночных животных, среди которых преобладали гигантские живущие в трубках черви Riftia pachyptila длиной до 2,5 м и толщиной до 5 см, крупные белые двустворчатые моллюски Calyptogena magnified и мидии Bathymodiolus thermophilus . Имелись там креветки, крабы и рыбы в немалых количествах.
Как объяснить наличие таких «оазисов» жизни в «пустыне», которой до недавнего времени считали дно океана на большой глубине?
Из проб воды, взятых у гидротермальных выходов, выделены бактерии, среди которых некоторые виды были Н2S-окисляющими хемолитоавтотрофами , идентифицированными как представители родов Thiomicrospira и Thiobacillus .
Такие бактерии могут составить первое звено трофической цепи в экосистеме гидротермальных источников, обеспечивая пищей различные виды животных.
Однако вскоре обнаружилось, что одно из преобладающих животных Riftia pachyptila не может питаться частичками пищи, поскольку представляет собой просто замкнутый мешок без ротового, анального отверстий и пищеварительной системы.
На переднем конце тела животного располагаются ярко окрашенные щупальца. В мешке заключены внутренние органы, самый крупный из них, занимающий почти всю полость тела, — трофосома, в которой обнаружено множество бактерий, окисляющих H2S, запасающих энергию в молекулах АТФ и использующих ее затем для фиксации СО2 в восстановительном пентозофосфатном цикле .
Бактерии локализованы внутри клеток трофосомы. Riftia pachyptila получает от бактерий органические соединения, а в обмен поставляет им необходимые для осуществления хемолитоавтотрофного метаболизма вещества (СО2, О2, H2S), поглощая их из внешней среды щупальцами (темно-красный цвет щупалец обусловлен присутствием большого количества крови, богатой гемоглобином), откуда они по кровеносной системе переносятся в трофосому к бактериям.
Таким образом, отношения между R. pachyptila и серобактериями — типичный пример внутриклеточного симбиоза мутуалистической природы.
Исследования других животных, обитающих у гидротермальных источников, показали, что R. pachyptila — не единственный вид, симбиотически связанный с хемолитоавтотрофными бактериями. МоллюскиCalyptogena magnifica и Bathymodiolus thermophilus также содержат хемосинтетических эндосимбионтов, но бактерии у них обитают в жабрах, где могут легко получать О2 и СО2 из проходящего сквозь жабры потока воды, a H2S моллюск поглощает своей вытянутой в длину ногой, которая погружена в источник, где концентрация сульфида наиболее высокая. Из ноги H2S переносится с кровью в жабры к бактериям.
Симбиозы, подобные описанному выше, обнаружены в других местах, богатых H2S, в том числе в мангровых и травяных соленых болотах, у мест просачивания нефти, в районах сброса сточных вод.
Число видов беспозвоночных, в которых найдены такие эндосимбионты, достаточно велико, и список этот постоянно растет.
Важное следствие открытия симбиозов, компонентом которых являются хемолитоавтотрофные бактерии, — существование экосистем, в которых первичными продуцентами служат не фотоавтотрофные, а хемолитоавтотрофные организмы.
Окисление восстановленных соединений серы до сульфатов , осуществляемое этими бактериями, приводит к подкислению окружающей среды, что может иметь положительные и отрицательные последствия.
Подкисление почвы приводит к переводу некоторых соединений, например фосфатов, в растворимую форму, что делает их доступными для растений. Окисление нерастворимых сульфидных минералов, сопровождающееся переводом металлов в растворимую форму, облегчает их добычу. Однако накопление серной кислоты в результате деятельности этих бактерий может приводить к порче и разрушению различных сооружений.
Железобактерии: общие сведения
Способность осаждать окислы железа и марганца на поверхности клеток присуща многим эубактериям, различающимся морфологическими и физиологическими признаками и принадлежащим к разным таксономическим группам.
В вопросе о том, какие организмы следует относить к железобактериям, нет единого мнения. С. Н. Виноградский впервые термин «железобактерии» применил для обозначения организмов, использующих энергию окисления Fe++ до Fe+++ для ассимиляции СО2, т.е. способных существовать хемолитоавтотрофно . X.Молиш к железобактериям относил все организмы, откладывающие вокруг клеток окислы железа или марганца независимо от того, связан ли этот процесс с получением клеткой энергии.
Накопление окислов железа и марганца на поверхности бактериальных клеток — результат двух взаимосвязанных процессов: аккумуляции (поглощения) клетками этих металлов из раствора и окисления , сопровождающегося обильным отложением нерастворимых окислов на поверхности бактерий.
Процесс аккумуляции тяжелых металлов из растворов в основе имеет физико-химическую природу и в значительной мере обусловлен химическим составом и свойствами поверхностных структур клетки.
Он включает связывание металлов внеклеточными структурами (капсулы, чехлы, слизистые выделения), клеточной стенкой и ЦПМ . Сорбционные свойства поверхностных клеточных структур определяются в большой степени суммарным отрицательным зарядом молекул, входящих в их состав.
Поглощение металлов приводит к значительному концентрированию их вокруг клеток по отношению к среде. Коэффициент накопления для железа и марганца может достигать значений 100000-1000000.
Как известно, Fe++ подвергается быстрому химическому окислению молекулярным кислородом при рН больше 5,5, что приводит к образованию нерастворимого Fe(OH)3.
Последний вместе с Fe++ неспецифически связывается клеточными кислыми экзополимерами. Подобный тип накопления железа не зависит от метаболической активности клеток.
Мn++ более устойчив к окислению О2, чем Fe++.
Его химическое окисление (Мn++ переходит в Мn++++) молекулярным кислородом с заметной скоростью происходит только при рН больше 8,5. Поэтому в нейтральной среде окисление марганца имеет только ферментативную природу.
Окисление Fe++ и Мn++ с последующим отложением нерастворимых окислов вокруг бактериальных клеток может быть результатом взаимодействия ионов металлов с продуктами бактериального метаболизма, в частности с Н2О2, образующейся в процессе окисления органических веществ при переносе электронов подыхательной цепи . Перекись водорода , возникающая в качестве промежуточного или конечного продукта окисления, выделяется из клеток и накапливается в окружающих их структурах.
В нейтральной или слабокислой среде окисление Fe++ до Fe+++ происходит в результате непосредственного взаимодействия с Н2О2:
2Fe+++ Н2О2 + 2Н+ переходит в 2Fe+++ + 2Н2О
Окисление марганца при взаимодействии с Н2О2 осуществляется при участии каталазы, выполняющей пероксидазную функцию.
Мn++ в этом случае служит донором электронов:
Мn++ + 2Н2О2→ МnО2 + 2Н2О
Описанные выше процессы протекают в капсулах, чехлах, слизистых выделениях, на поверхности клеточной стенки, в которых концентрируются все компоненты реакции: восстановленные формы железа и марганца, перекись водорода, каталаза.
Физиологический смысл процессов окисления Fe++ и Мn++ с участием Н2О2 — детоксикация вредного продукта метаболизма. Ни в одном случае окисление железа и марганца не приводит к получению бактериями энергии.
Наконец, среди железобактерий есть организмы, у которых окисление Fe++ связано с получением энергии.
В этом случае отложение окислов железа служит показателем активности энергетических процессов. Возможность получения энергии бактериями при окислении Мn++ экспериментально не доказана.
В изучении железобактерий в последнее время достигнуты большие успехи, связанные с получением чистых культур ряда этих организмов.
Стало понятным, что это разнообразная группа бактерий, способных окислять и откладывать окислы железа и/или марганца вне или иногда внутри клетки. (К железобактериям, откладывающим железо внутри клетки, относятся магниточувствительные бактерии).
На основании морфологических характеристик все железобактерии могут быть разделены на две группы: нитчатые железобактерии и одноклеточные железобактерии .
Эубактерий, описанные в этом разделе, широко распространены в природе и могут существовать в большом диапазоне условий.
Облигатные ацидофилы обнаружены в подземных водах сульфидных месторождений, кислых водах железистых источников и кислых озерах с высоким содержанием закисного железа. Нитчатые формы также занимают вполне определенные экологические ниши. Представители рода Leptothrix — обитатели олиготрофных железистых поверхностных вод, Sphaerotilusпредпочитают среды с высоким содержанием органических веществ.
К сапротрофам относятся
Бактерии — паразиты болезнетворные (туберкулезная палочка, дизентерийная палочка ) P.S. Бактерии — доядерные организмы (прокариоты) . Бактерии — в основном гетеротрофы по способу питания. Подавляющее большинство бактерий — сапрофиты (сапротрофы) — разрушители органических веществ. Существование бактерий — автотрофов, способных образовывать органические вещества из неорганических. Бактерии — паразиты болезнетворные (туберкулезная палочка, дизентерийная палочка и др.) .
Образование бактериями спор — приспособление к перенесению неблагоприятных условий.
Палочка Коха. Сапрофит — питается гнилостными останками. Автотроф — питается посредством фотосинтеза. Паразит — живет в теле хозяина, при этом организм хозяина несёт ущерб, а организм паразита пользу.
Вы в вопросе намешали праведное с грешным, Сапрофиты у людей не вызывают заболеваний. автотроф— это разделени бактерий по способу питания бактерий.
Латинский префикс «myco-» означает гриб (воск) , в данном случае понятие отражает «воскоподобные» компоненты, составляющие часть клеточной стенки микобактериальных клеток.
Аэробны и неподвижны. не образует спор и капсул, образуют L-формы. Относится к внутриклеточной инфекции. Принято считать грам-положительными бактериями.
Микобактерии туберкулёза нечувствительны к рассеянному солнечному свету и могут более года существовать во внешней среде без потери жизнеспособности. Коротковолновое ультрафиолетовое изучение оказывает универсальное бактерицидное действие на все микроорганизмы. Однако в реальных условиях, когда микобактерии туберкулёза находятся во взвешенном состоянии в виде клеточных агломератов с пылевыми частицами, их устойчивость к ультрафиолетовому излучению возрастает.
Не выделяет эндо- и экзотоксинов, поэтому при инфицировании ярких клинических симптомов как правило не возникает. По мере размножения и формирования повышенной чувствительности тканей к туберкулопротеидам (белки-носители антигенных свойств) возникают первые признаки инфицирования (положительная реакция на туберкулин).
Источник



