
Пути передачи сигнала клетки




![]()
![]()
-аутокринная регуляция (саморегуляция биохимических процессов в клетки методами обратной связи, субстратной индукцией, а также с помощью свободно-радикальных метаболитов, супероксида и оксида азота)
-паракринная регуляция (тканевые гормоны, нейромедиаторы)
-эндокринная регуляция (истинные гормоны)
101.Механизм действия сигнальных молекул…,соприженных с G-белками и аденилатциклазой.
В зависимости от строения гормона существуют два типа взаимодействия. Если молекула гормона липофильна, (например, стероидные гормоны), то она может проникать через липидный слой наружной мембраны клеток-мишеней. Если молекула имеет большие размеры или является полярной, то ее проникновение внутрь клетки невозможно. Поэтому для липофильных гормонов рецепторы находятся внутри клеток-мишеней, а для гидрофильных — рецепторы находятся в наружной мембране.
Для получения клеточного ответа на гормональный сигнал в случае гидрофильных молекул действует внутриклеточный механизм передачи сигнала. Это происходит с участием веществ, которых называют «ВТОРЫМИ ПОСРЕДНИКАМИ». Молекулы гормонов очень разнообразны по форме, а «вторые посредники» — нет.
Надёжность передачи сигнала обеспечивает очень высокое сродство гормона к своему белку-рецептору.
Что такое посредники, которые участвуют во внутриклеточной передаче гуморальных сигналов? Это циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок — кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы — ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях.
Разберем более подробно механизмы действия гормонов и внутриклеточных посредников. Существует два главных способа передачи сигнала в клетки-мишени от сигнальных молекул с мембранным механизмом действия:
1. АДЕНИЛАТЦИКЛАЗНАЯ (ИЛИ ГУАНИЛАТЦИКЛАЗНАЯ) СИСТЕМЫ
2. ФОСФОИНОЗИТИДНЫЙ МЕХАНИЗМ
Основные компоненты: мембранный белок-рецептор, G-белок, фермент аденилатциклаза, гуанозинтрифосфат, протеинкиназы.
Кроме того, для нормального функционирования аденилатциклазной системы, требуется АТФ.
Схема аденилатциклазной системы представлена на рисунке:
Как видно из рисунка, белок-рецептор, G-белок, рядом с которым располагаются ГТФ и фермент (аденилатциклаза) встроены в мембрану клетки.
До момента действия гормона эти компоненты находятся в диссоциированнном состоянии, а после образования комплекса сигнальной молекулы с белком-рецептором происходят изменения конформации G-белка. В результате одна из субъединиц G-белка приобретает способность связываться с ГТФ.
Комплекс “G-белок-ГТФ” активирует аденилатциклазу. Аденилатциклаза начинает активно превращать молекулы АТФ в ц-АМФ.
ц-АМФ обладает способностью активировать особые ферменты — протеинкиназы, которые катализируют реакции фосфорилирования различных белков с участием АТФ. При этом в состав белковых молекул включаются остатки фосфорной кислоты. Главным результатом этого процесса фосфорилирования является изменение активности фосфорилированного белка. В различных типах клеток фосфорилированию в результате активации аденилат-циклазной системы подвергаются белки с разной функциональной активностью. Например, это могут быть ферменты, ядерные белки, мембранные белки. В результате реакции фосфорилирования белки могут становятся функционально активными или неактивными.
Такие процессы будут приводить к изменениям скорости биохимических процессов в клетке-мишени.
Активация аденилатциклазной систтемы длится очень короткое время, потому что G-белок после связывания с аденилатциклазой начинает проявлять ГТФ-азную активность. После гидролиза ГТФ G-белок восстанавливает свою конформацию и перестает активировать аденилатциклазу. В результате прекращается реакция образования цАМФ.
Кроме участников аденилатциклазной системы в некоторых клетках-мишенях имеются белки-рецепторы, связанные с G-белками, которые приводят к торможению аденилатциклазы. При этом комплекс “GTP-G-белок” ингибирует аденилатциклазу.

Когда останавливается образование цАМФ, реакции фосфорилирования в клетке прекращаются не сразу: пока продолжают существовать молекулы цАМФ — будет продолжаться и процесс активации протеинкиназ. Для того, чтобы прекратить действие цАМФ, в клетках существует специальный фермент — фосфодиэстераза, который катализирует реакцию гидролиза 3′,5′-цикло-АМФ до АМФ.
Некоторые вещества, обладающие ингибирующим действием на фосфодиэстеразу, (например, алкалоиды кофеин, теофиллин), способствуют сохранению и увеличению концентрации цикло-АМФ в клетке. Под действием этих веществ в организме продолжительность активации аденилатциклазной системы становится больше, то есть усиливается действие гормона.
102.Механизм действия сигнальных молекул…,соприженных с G-белками и фосфолипазой С
В зависимости от строения гормона существуют два типа взаимодействия. Если молекула гормона липофильна, (например, стероидные гормоны), то она может проникать через липидный слой наружной мембраны клеток-мишеней. Если молекула имеет большие размеры или является полярной, то ее проникновение внутрь клетки невозможно. Поэтому для липофильных гормонов рецепторы находятся внутри клеток-мишеней, а для гидрофильных — рецепторы находятся в наружной мембране.
Для получения клеточного ответа на гормональный сигнал в случае гидрофильных молекул действует внутриклеточный механизм передачи сигнала. Это происходит с участием веществ, которых называют «ВТОРЫМИ ПОСРЕДНИКАМИ». Молекулы гормонов очень разнообразны по форме, а «вторые посредники» — нет.
Надёжность передачи сигнала обеспечивает очень высокое сродство гормона к своему белку-рецептору.
Что такое посредники, которые участвуют во внутриклеточной передаче гуморальных сигналов? Это циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок — кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы — ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях.
Разберем более подробно механизмы действия гормонов и внутриклеточных посредников. Существует два главных способа передачи сигнала в клетки-мишени от сигнальных молекул с мембранным механизмом действия:
1. АДЕНИЛАТЦИКЛАЗНАЯ (ИЛИ ГУАНИЛАТЦИКЛАЗНАЯ) СИСТЕМЫ
2. ФОСФОИНОЗИТИДНЫЙ МЕХАНИЗМ
Инозитолтрифосфат -это вещество, которое является производным сложного липида — инозитфосфатида. Оно образуется в результате действия специального фермента — фосфолипазы “С”, который активируется в результате конформационных изменений внутриклеточного домена мембранного белка-рецептора.
Этот фермент гидролизует фосфоэфирную связь в молекуле фосфатидил-инозитол-4,5-бисфосфата и в результате образуются диацилглицерин и инозитолтрифосфат.
Известно, что образование диацилглицерина и инозитолтрифосфата приводит к увеличению концентрации ионизированного кальция внутри клетки. Это приводит к активации многих кальций-зависимых белков внутри клетки, в том числе активируются различные протеинкиназы. И здесь, как и при активации аденилатциклазной системы, одной из стадий передачи сигнала внутри клетки является фосфорилирование белков, которое в приводит к физиологическому ответу клетки на действие гормона.
В работе фосфоинозитидного механизма передачи сигналов в клетке-мишени принимает участие специальный кальций-связывающий белок — кальмодулин. Это низкомолекулярный белок (17 кДа), на 30% состоящий из отрицательно заряженных аминокислот (Глу, Асп) и поэтому способный активно связывать Са +2 . Одна молекула кальмодулина имеет 4 кальций-связывающих участка. После взаимодействия с Са +2 происходят конформационные изменения молекулы кальмодулина и комплекс “Са +2 -кальмодулин” становится способным регулировать активность (аллостерически угнетать или активировать) многие ферменты — аденилатциклазу, фосфодиэстеразу, Са +2 ,Мg +2 -АТФазу и различные протеинкиназы.
В разных клетках при воздействии комплекса “Са +2 -кальмодулин” на изоферменты одного и того же фермента (например, на аденилатциклазу разного типа) в одних случаях наблюдается активация, а в других — ингибирование реакции образования цАМФ. Такие различные эффекты происходят потому, что аллостерические центры изоферментов могут включать в себя различные радикалы аминокислот и их реакция на действие комплекса Са +2 -кальмодулин будет отличаться.
Таким образом, в роли «вторых посредников» для передачи сигналов от гормонов в клетках-мишенях могут быть:
Источник
Способ передачи сигнальных молекул при котором изменение функции клетки происходит
Все клетки, от прокариот до клеток растений и животных, способны воспринимать сигналы извне и реагировать на них стереотипным образом, который позволяет им выживать, приспосабливаться и функционировать в соответствии с потребностями всего организма.
Эта реакция клеток выражается не просто в форме неких физических или химических явлений или изменений в их локальном окружении. Скорее, следут сказать, что клетка экспрессирует целый ансамбль сенсорных белков, или рецепторов, узнающих специфические внешние сигналы.
В ответ на эти сигналы рецепторы активируют разнообразные внутриклеточные регуляторные белки, которые, в свою очередь, обеспечивают адекватную реакцию клетки. Процесс узнавания внешнего сигнала и передачи соответствующей информации на внутриклеточные мишени называется преобразованием сигнала.
Клетка реагирует на все виды сигналов. Бактерии реагируют на питательные ингредиенты, токсины, тепло, холод, свет и химические сигналы, исходящие от других бактерий.
Клетки многоклеточных организмов экспрессируют рецепторы, специфичные по отношению к гормонам, нейромедиаторам, аутокринным и паракринным агентам (гормоноподобные вещества, секретируемые самой клеткой или ее окружением), к запаху и к молекулам, регулирующим рост или дифференцировку, а также к белкам, расположенным на поверхности клеток. Типичная клетка млекопитающих экспрессирует около пятидесяти различных рецепторов, которые проявляют чувствительность к разнообразным сигналам, а всего у млекопитающих экспрессируется несколько тысяч рецепторов.
Несмотря на большое разнообразие клеток и огромное количество веществ, которые они узнают, основные классы белков и процессы, участвующие в системе преобразования сигнала в клетках, достаточно консервативны.
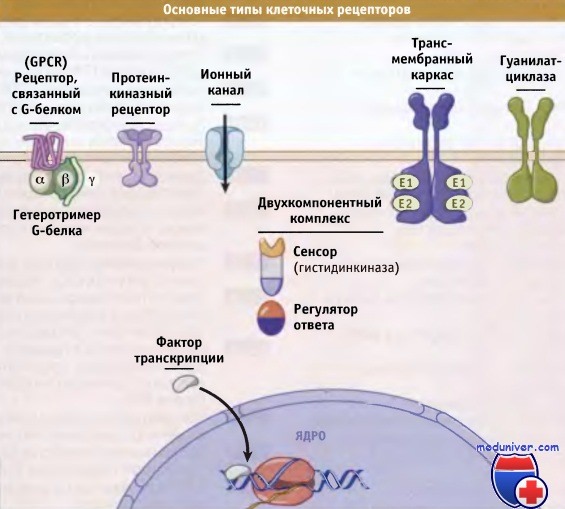
• Рецепторы, связанные с G-белком, состоящие из семи спиралей, проходящих через мембрану. Они обеспечивают активацию гетеротримерных ГТФ-связывающих белков, которые называются G-белками. Эти белки связаны с внутренней стороной плазматической мембраны и передают сигналы на разные внутриклеточные белки.
• Протеинкиназные рецепторы часто представляют собой димеры белков, проходящих через мембрану, которые фосфорилируют внутриклеточные субстраты и, таким образом, изменяют форму и функции белков-мишеней. Эти протеинкиназы часто содержат домены, ответственные за взаимодействие с другими белками и принимающие участие в организации комплексов сигнальных белков, расположенных на внутренней поверхности плазматической мембраны.
• Фосфопротеинфосфатазы аннулируют эффект протеинкиназ, удаляя фосфатные группы.
• Прочие ферменты, молекулы которых проходят через мембрану, например, гуанилатциклаза, обладают структурой, близкой к рецепторным проте-инкиназам, однако характеризуются различными ферментативными активностями. Гуанилатциклаза катализирует превращение ГТФ в 3’5′-циклоГТФ, который участвует в распространении сигнала.
• Ионные каналы, хотя и имеют ряд отличий в деталях структуры, обычно представляют собой олигомеры субъединиц, из которых каждая состоит из нескольких трансмембранных сегментов. Эти субъединицы способны менять свою конформацию и относительное расположение, тем самым регулируя ионные потоки через центральную пору.
• Двухкомпонентные системы могут обладать трансмембранным расположением или локализуются в цитозоле. Число субъединиц у них также варьирует, но каждая двухкомпонентная система содержит гистидинкиназный домен или субъединицу, находящуюся под контролем сигнальной молекулы, и регулятор ответа, в структуре которого присутствует остаток аспарагиновой кислоты (Asp), способный фосфорилироваться.
• Некоторые рецепторы представляют собой трансмембранный каркас, который меняет конформацию или обладает способностью к олигомеризации своих доменов, расположенных с клеточной стороны. Это происходит в ответ на появление внеклеточных сигнальных молекул или лигандов. Таким образом, происходит отбор регуляторных белков, взаимодействующих между собой в определенных местах мембраны.
• Ядерные рецепторы представляют собой факторы транскрипции, часто гетеродимеры, которые находятся в цитоплазме до момента их активации агонистами, или постоянно присутствуют в ядре. Биохимические процессы, участвующие в преобразовании сигнала в разных клетках, поразительно сходны. Для обнаружения и обработки сигнала, клетки бактерий, грибов, растений и животных используют аналогичные белки и мультибелковые комплексы.
Например, у растений, грибов и в клетках животных обнаружены обладающие близкой структурой гетеротримерные G-белки и соответствующие им рецепторы, сохранившиеся в ходе эволюции. Аналогичным образом, в бактериях, грибах и у животных обнаружен 3’5′-циклический АМФ (цАМФ), представляющий собой внутриклеточную сигнальную молекулу; во всех клетках эукариот аналогичную роль играет Са2+. Во всех клетках в регуляции активности ферментов участвуют протеинкиназы и фосфопротеинфосфатазы.
Хотя основные биохимические компоненты и процессы передачи сигнала достаточно консервативны и используются многократно, они часто вовлекаются в самые разнообразные процессы и принимают участие в реализации множества различных физиологических реакций. Например, цАМФ образуется в бактериях, грибах и у млекопитающих с участием близких по свойствам ферментов, и в каждом организме белки-мишени для этого метаболита различны; так, для некоторых видов миксомицетов он является феромоном.
Часто для регуляции определенного процесса, например транскрипции, ионного транспорта, подвижности и метаболизма, клетки используют одни и те же сигнальные белки. Такие системы передачи сигнала собираются в сигнальные сети с тем, чтобы клетка могла координировать реакции на множественные сигналы с выполняемыми в данный момент функциями. В настоящее время в последовательностях отдельных процессов системы передачи сигнала, и в сети в целом, можно обнаружить присутствие элементов, характерных для компьютерных сетей.
Это усилители, логические шлюзы, контроль обратной и прямой связи, и память.
В дальнейших статьях вначале рассматриваются принципы организации клеточных систем передачи сигнала и общие вопросы стратегии, а затем обсуждаются соответствующие биохимические компоненты и процессы, а также пути конкретной реализации изложенных принципов.
 Рецепторы группируются в относительно небольшую группу семейств,
Рецепторы группируются в относительно небольшую группу семейств,
которые характеризуются общими механизмами действия и общностью структуры.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник



