
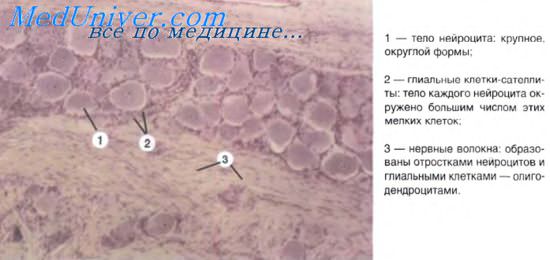
Способ передачи нервного импульса пресинаптической части синапса
Механизм химической передачи нервных импульсов через синапс сводится к тому, что содержимое синаптических пузырьков (медиатор) поступает в виде небольших порций (квантов) в синаптическую щель и взаимодействует затем с рецепторными белками постсинаптической мембраны. Это вызывает деполяризацию мембраны и возбуждение следующего нейрона. Ультраструктурные особенности синапса и механизм передачи импульсов определяют строгую однонаправленность передачи импульсов, что лежит в основе проведения импульсов по рефлекторным дугам.
В зависимости от того, какое вещество выполняет роль нейромедиатора, синапсы подразделяются на холинергические (медиатор — ацетилхолин), адренергические (адреналин и норадреналин), дофаминергические (дофамин), серотонинергические (серотонин), пептидергические (медиаторы — пептиды и аминокислоты, например, мет-энкефалин, гамма-аминомасляная кислота, глицин и др.).
Нейрохимические синапсы подразделяются в функциональном отношении на две противоположные по своему значению группы — возбуждающие и тормозные. Свойства этих синапсов зависят как от медиаторов, так и от ультраструктурных особенностей синапсов. Так, некоторые медиаторы (например, глутамат) характерны для возбуждающих синапсов, а в тормозных синапсах медиатором является гамма-аминомасляная кислота. Предполагают, что в возбуждающих синапсах к постсинаптической мембране прилежит электронно-плотное вещество, в связи с чем синапс приобретает асимметричное строение.
В пресинаптических частях таких соединений содержатся круглые синаптические пузырьки. Тормозные синапсы имеют симметричное строение. Синаптическая щель у них сужена и в пресинаптической части содержатся уплощенные синаптические пузырьки.
Кроме нейрохимических синапсов между нервными клетками (преимущественно между дендритами или телами нейронов) возникают электротонические синапсы. Последние у млекопитающих встречаются редко и по строению соответствуют щелевым контактам. Они проводят возбуждение благодаря формированию трансмембранных каналов — коннексонов.
Каждый нейрон на своей поверхности имеет огромное количество (до 10000) синапсов. Интеграция импульсов в пределах отдельного нейрона происходит так: от синапсов, расположенных на дендритах и теле, по его плазмолемме передается импульс к аксонному холмику (генераторному пункту нейрона), где путем суммирования всех возбуждающих и тормозных импульсов возникает результирующий потенциал действия.
Синаптические структуры обладают высокой чувствительностью к действию токсических факторов, в том числе психотропных отравляющих веществ. Их изменения играют важную роль в механизмах реактивности нервных тканей.
Рецепторные нервные окончания подразделяются на две группы: экстерорецепторы, воспринимающие раздражения из внешней среды, и интерорецепторы, служащие для восприятия раздражений из внутренней среды организма. В зависимости от специфичности раздражения, воспринимаемого рецептором, различают следующие чувствительные нервные окончания: механорецепторы, барорецепторы, хеморецепторы, терморецепторы, болевые рецепторы (ноцицепторы). Все рецепторы по особенностям их строения подразделяют на свободные и несвободные нервные окончания.
Первые из них состоят только из конечных разветвлений периферического отростка чувствительного нейрона; вторые — имеют в составе рецептора кроме нервного отростка глиальный компонент, а нередко и соединительнотканную капсулу. В последнем случае несвободные рецепторные окончания называют инкапсулированными. К числу таких рецепторов относятся пластинчатые, осязательные тельца, концевые колбы, мышечные веретена и др.
Эффекторные нервные окончания подразделяются на двигательные и секреторные. Двигательный, или моторный, эффектор передает нервные импульсы на рабочие органы и ткани. В скелетных мышцах они называются нервно-мышечными (аксомышечными) окончаниями. Терминальное разветвление аксона двигательного нейрона заканчивается булавовидным расширением. Оно соответствует пресинаптической части нервно-мышечного синапса, содержит скопление синаптических пузырьков и митохондрий, ограничено пресинаптической мембраной характерного строения.
Плазмолемма мышечного волокна в этой области образует многочисленные складки и углубления. В постсинаптической части волокна находится зернистая саркоплазма с большим количеством митохондрий и овальных ядер. Синаптическая щель имеет ширину 50-100 нм. У человека медиатором в нервно-мышечных синапсах является ацетилхолин. Моторные окончания в гладкой мышечной ткани, а также секреторные эффекторы имеют вид тонких пучков аксонов или их одиночных терминалей, прилежащих к клеткам иннервируемых тканей.
Возрастные изменения нервных тканей связаны с постепенным уменьшением запаса нервных клеток, особенно — чувствительных нейронов, а также снижением уровня метаболических процессов, что выражается в закономерном накоплении включений липофусцина («пигмент изнашивания») в нейроплазме.
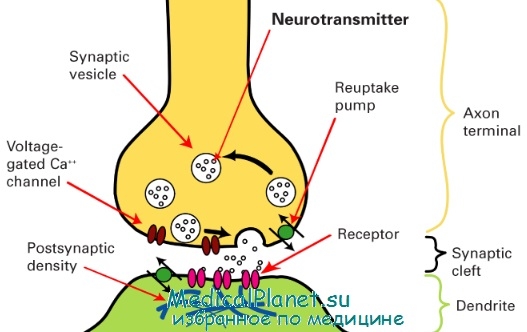 Схема строения синапса
Схема строения синапса
Источник
Способ передачи нервного импульса пресинаптической части синапса
Область контакта между двумя нейронами называют синапсом.
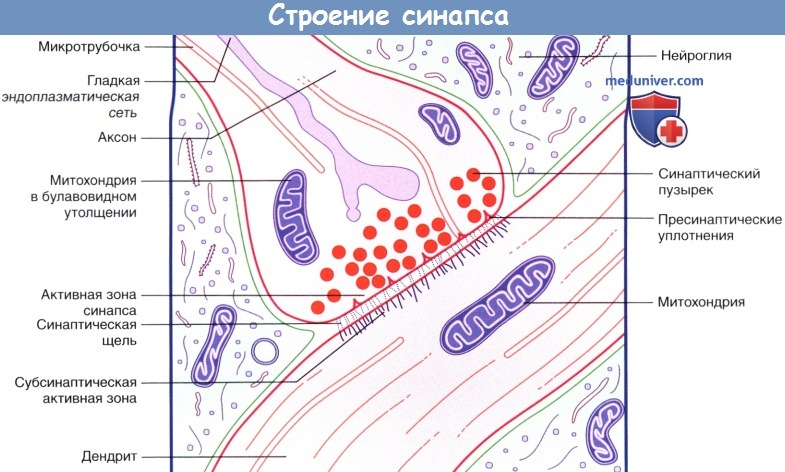 Внутреннее строение аксодендритического синапса.
Внутреннее строение аксодендритического синапса.
а) Электрические синапсы. Электрические синапсы в нервной системе млекопитающих встречаются редко. Они образованы щелевидными контактами (нексусами) между дендритами или сомами соприкасающихся нейронов, которые соединяются с помощью цитоплазматических каналов диаметром 1,5 нм. Процесс передачи сигнала происходит без синаптической задержки и без участия медиаторов.
Посредством электрических синапсов возможно распространение электротонических потенциалов от одного нейрона к другому. Вследствие тесного синаптического контакта модуляция проведения сигнала невозможна. Задача этих синапсов — осуществление одновременного возбуждения нейронов, выполняющих одинаковую функцию. Примером служат нейроны дыхательного центра продолговатого мозга, которые во время вдоха синхронно генерируют импульсы. Кроме того, примером могут служить нейронные цепи, управляющие саккадами, при которых точка фиксации взора перемещается от одного объекта внимания к другому.
б) Химические синапсы. Большинство синапсов нервной системы — химические. Функционирование таких синапсов зависит от высвобождения медиаторов. Классический химический синапс представлен пресинаптической мембраной, синаптической щелью и постсинаптической мембраной. Пресинаптическая мембрана — часть булавовидного расширения нервного окончания клетки, передающей сигнал, а постсинаптическая мембрана — часть клетки, получающей сигнал.
Медиатор высвобождается из булавовидного расширения посредством экзоцитоза, проходит через синаптическую щель и связывается с рецепторами на постсинаптической мембране. Под постсинаптической мембраной расположена субсинаптическая активная зона, в которой после активации рецепторов постсинаптической мембраны происходят разнообразные биохимические процессы.
В булавовидном расширении расположены содержащие медиаторы синаптические пузырьки, а также большое количество митохондрий и цистерны гладкой эндоплазматической сети. Применение традиционных методик фиксации при исследовании клеток позволяет различить на пресинаптической мембране пресинаптические уплотнения, ограничивающие активные зоны синапса, к которым при помощи микротрубочек направляются синаптические пузырьки.
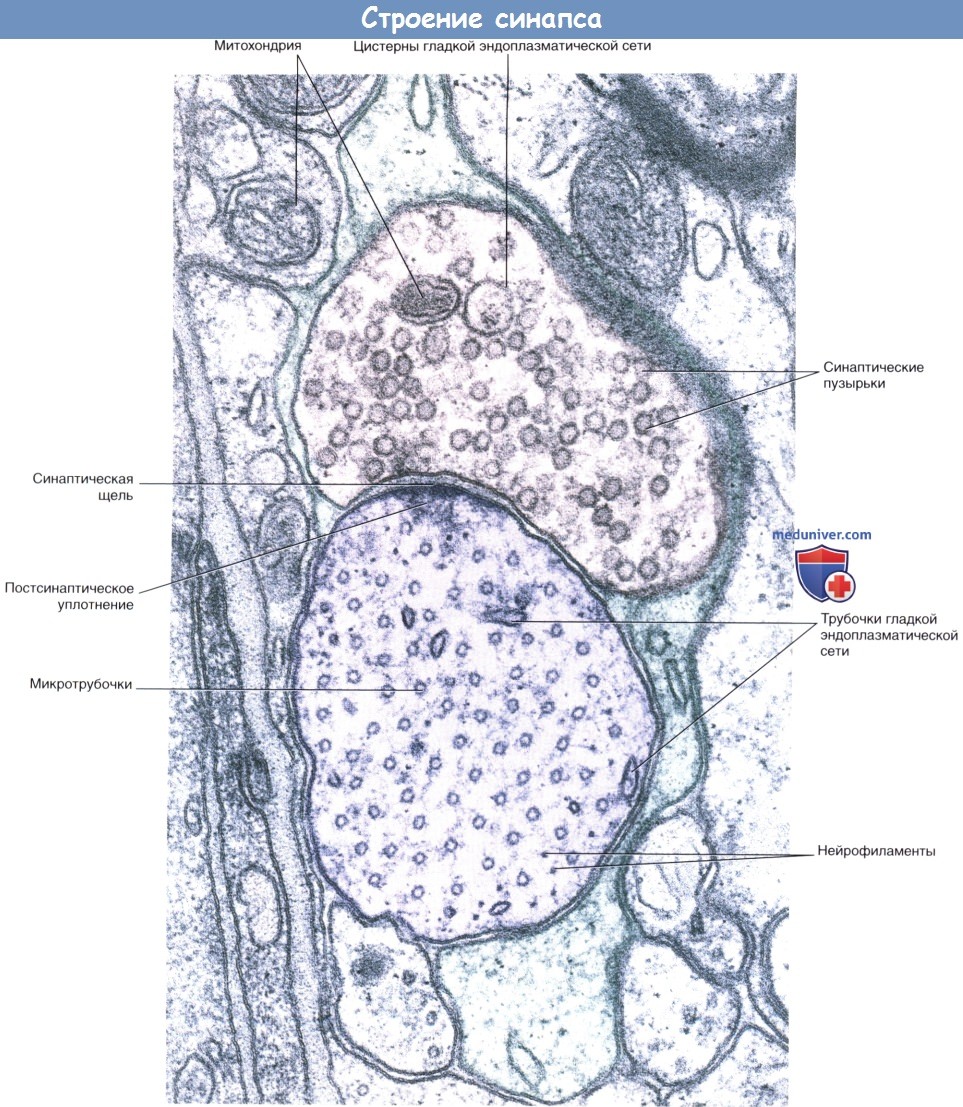 Аксодендритический синапс.
Аксодендритический синапс.
Срез препарата спинного мозга: синапс между концевым участком дендрита и, предположительно, двигательным нейроном.
Наличие округлых синаптических пузырьков и постсинаптического уплотнения характерно для возбуждающих синапсов.
Срез дендрита проведен в поперечном направлении, о чем свидетельствует наличие множества микротрубочек.
Кроме того, видны некоторые нейрофиламенты. Участок синапса окружен протоплазматическим астроцитом. 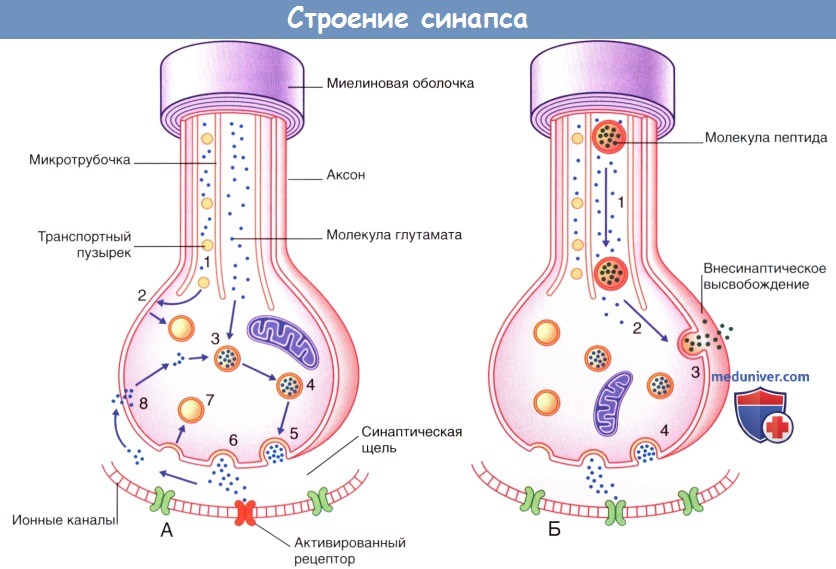 Процессы, происходящие в нервных окончаниях двух типов.
Процессы, происходящие в нервных окончаниях двух типов.
(А) Синаптическая передача небольших молекул (например, глутамата).
(1) Транспортные пузырьки, содержащие мембранные белки синаптических пузырьков, направляются вдоль микротрубочек к плазматической мембране булавовидного утолщения.
В это же время происходит перенос молекул ферментов и глутамата путем медленного транспорта.
(2) Мембранные белки пузырьков выходят из плазматической мембраны и формируют синаптические пузырьки.
(3) Глутамат погружается в синаптические пузырьки; происходит накопление медиатора.
(4) Пузырьки, содержащие глутамат, подходят к пресинаптической мембране.
(5) В результате деполяризации происходит экзоцитоз медиатора из частично разрушенных пузырьков.
(6) Высвобождающийся медиатор распространяется диффузно в области синаптической щели и активирует специфические рецепторы на постсинаптической мембране.
(7) Мембраны синаптических пузырьков транспортируются обратно в клетку путем эндоцитоза.
(8) Происходит частичный обратный захват глутамата в клетку для повторного использования.
(Б) Передача нейропептидов (например, субстанции Р), осуществляющаяся одновременно с синаптической передачей (например, глутамата).
Совместная передача этих веществ происходит в центральных нервных окончаниях униполярных нейронов, обеспечивающих болевую чувствительность.
(1) Синтезированные в комплексе Гольджи (в области перикариона) пузырьки и предшественники пептидов (пропептиды) транспортируются к булавовидному расширению путем быстрого транспорта.
(2) При их попадании в область булавовидного утолщения завершается процесс формирования молекулы пептида, и пузырьки транспортируются к плазматической мембране.
(3) Деполяризация мембраны и перенос содержимого пузырьков в межклеточное пространство путем экзоцитоза.
(4) Одновременно с этим происходит высвобождение глутамата.
1. Активация рецепторов. Молекулы медиаторов проходят через синаптическую щель и активируют рецепторные белки, расположенные парами на постсинаптической мембране. Активация рецепторов запускает ионные процессы, которые приводят к деполяризации постсинаптической мембраны (возбуждающее постсинаптическое действие) или гиперполяризации постсинаптической мембраны (тормозящее постсинаптическое действие). Изменение электротонуса передается в сому в виде затухающего по мере распространения электротонического потенциала, за счет которого происходит изменение потенциала покоя в начальном сегменте аксона.
Ионные процессы подробно описаны в отдельной статье на сайте. При преобладании возбуждающих постсинаптических потенциалов начальный сегмент аксона деполяризуется до порогового уровня и генерирует потенциал действия.
Наиболее распространенный возбуждающий медиатор ЦНС — глутамат, а тормозной — гамма-аминомасляная кислота (ГАМК). В периферической нервной системе медиатором для двигательных нейронов поперечно-полосатой мускулатуры служит ацетилхолин, а для чувствительных нейронов — глутамат.
Последовательность процессов, происходящих в глутаматергических синапсах, показана на рисунке ниже. При передаче глутамата совместно с другими пептидами высвобождение пептидов осуществляется внесинаптическим путем.
Большинство чувствительных нейронов помимо глутамата выделяет и другие пептиды (один или несколько), высвобождающиеся в различных участках нейрона; однако основная функция этих пептидов — модуляция (повышение или снижение) эффективности синаптической передачи глутамата.
Кроме того, нейротрансмиссия может происходить путем диффузной внесинаптической передачи сигнала, характерной для моноаминергических нейронов (нейронов, использующих биогенные амины для обеспечения нейротрансмиссии). Выделяют две разновидности моноаминергических нейронов. В одних нейронах осуществляется синтез катехоламинов (норадреналина или дофамина) из аминокислоты тирозина, а в других — серотонина из аминокислоты триптофана. Например, дофамин высвобождается как в синаптической области, так и из варикозных утолщений аксона, в которых также происходит синтез этого нейромедиатора.
Дофамин проникает в межклеточную жидкость ЦНС и до момента деградации способен активировать специфические рецепторы на расстоянии до 100 мкм. Моноаминергические нейроны присутствуют во многих структурах ЦНС; нарушение передачи импульса этими нейронами приводит к различным заболеваниям, среди которых выделяют болезнь Паркинсона, шизофрению и глубокую депрессию.
Оксид азота (газообразная молекула) также участвует в диффузной нейропередаче в глутаматергической системе нейронов. Избыточное влияние оксида азота оказывает цитотоксическое действие, особенно в тех участках, кровоснабжение которых нарушено за счет тромбоза артерий. Глутамат также является потенциально цитотоксическим нейромедиатором.
В отличие от диффузной нейротрансмиссии, традиционную синаптическую передачу сигнала ввиду ее относительной стабильности называют «проводниковой».
в) Резюме. Мультиполярные нейроны ЦНС состоят из сомы, дендритов и аксона; аксон образует коллатеральные и терминальные ветви. В соме расположены гладкая и шероховатая эндоплазматическая сети, комплексы Гольджи, нейрофиламенты и микротрубочки. Микротрубочки пронизывают нейрон на всем протяжении, принимают участие в процессе антероградного транспорта синаптических пузырьков, митохондрий и веществ для построения мембран, а также обеспечивают ретроградный транспорт «маркерных» молекул и разрушенных органелл.
Существует три вида химических межнейрональных взаимодействий: синаптическое (например, глутаматергическое), внесинаптическое (пептидергическое) и диффузное (например, моноаминергическое, серотонинергическое).
Химические синапсы классифицируют по анатомическому строению на аксодендритические, аксосоматические, аксоаксональные и дендро-дендритические. Синапс представлен пре- и постсинаптическими мембранами, синаптической щелью и субсинаптической активной зоной.
Электрические синапсы обеспечивают одновременную активацию целых групп нейронов, образуя между ними электрические связи за счет щелевидных контактов (нексусов).
![]() Диффузная нейротрансмиссия в головном мозге.
Диффузная нейротрансмиссия в головном мозге.
Аксоны глутаматергического (1) и дофаминергического (2) нейронов образуют плотные синаптические контакты с отростком звездчатого нейрона (3) полосатого тела.
Дофамин высвобождается не только из пресинаптической области, но и из варикозного утолщения аксона, откуда диффузно распространяется в межклеточное пространство и активирует дофаминовые рецепторы дендритного ствола и стенки перицита капилляра. 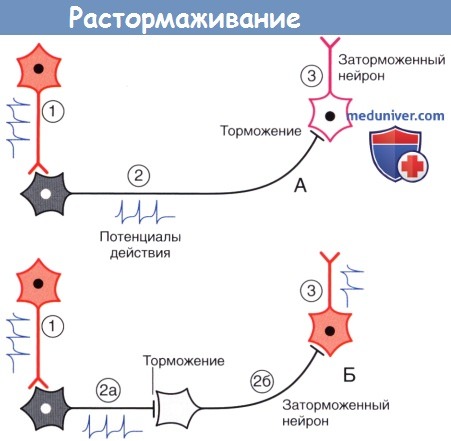 Растормаживание.
Растормаживание.
(А) Возбуждающий нейрон 1 активирует тормозной нейрон 2, который в свою очередь затормаживает нейрон 3.
(Б) Появление второго тормозного нейрона (2б) оказывает противоположное влияние на нейрон 3, поскольку происходит торможение нейрона 2б.
Спонтанно-активный нейрон 3 генерирует сигналы в условиях отсутствия тормозных влияний.
2. Лекарственные средства — «ключи» и «замки». Рецептор можно сравнить с замком, а медиатор — с подходящим к нему ключом. В том случае, если процесс высвобождения медиатора нарушится с возрастом или в результате какого-либо заболевания, лекарственное средство может сыграть роль «запасного ключа», выполняющего аналогичную медиатору функцию. Такое лекарственное средство называют агонистом. В то же время в случае чрезмерной продукции медиатор может быть «перехвачен» блокатором рецептора — «фальшивым ключом», который свяжется с «замком»-рецептором, но при этом не вызовет его активацию.
3. Торможение и растормаживание. Функционирование спонтанно-активных нейронов сдерживается под влиянием тормозных нейронов (обычно, ГАМКергических). Деятельность тормозных нейронов, в свою очередь, может быть ингибирована воздействующими на них другими тормозными нейронами, в результате чего происходит растормаживание клетки-мишени. Процесс растормаживания — важная особенность нейрональной активности в базальных ганглиях.
4. Редкие виды химических синапсов. Выделяют два типа аксоаксональных синапсов. В обоих случаях булавовидное утолщение образует тормозной нейрон. Синапсы первого типа образуются в области начального сегмента аксона и передают мощное ингибирующее влияние тормозного нейрона. Синапсы второго типа образуются между булавовидным утолщением тормозного нейрона и булавовидными утолщениями возбуждающих нейронов, что приводит к угнетению высвобождения медиаторов. Этот процесс получил название пресинаптического торможения. В этом плане традиционный синапс обеспечивает постсинаптичсекое торможение.
Дендро-дендритические (Д-Д) синапсы образуются между дендритными шипиками дендритов смежных шипиковых нейронов. Их задача — не генерирование нервного импульса, а изменение электротонуса клетки-мишени. В последовательных Д-Д-синапсах синаптические пузырьки располагаются только в одном дендритном шипике, а в реципрокном Д-Д-синапсе— в обоих. Возбуждающие Д-Д-синапсы изображены на рисунке ниже. Тормозные Д-Д-синапсы широко представлены в переключающих ядрах таламуса.
Кроме того, выделяют немногочисленные сомато-дендритические и сомато-соматические синапсы.
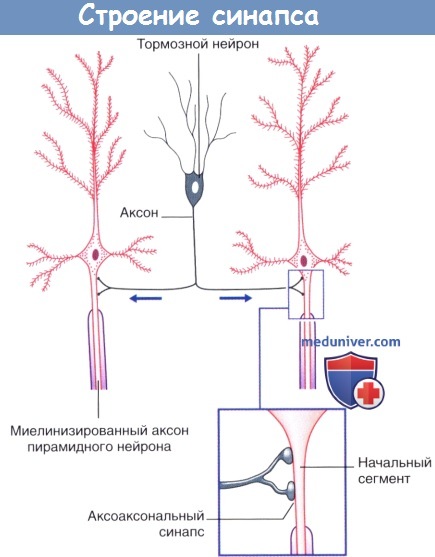 Аксоаксональные синапсы коры головного мозга.
Аксоаксональные синапсы коры головного мозга.
Стрелками указано направление проведения импульсов. 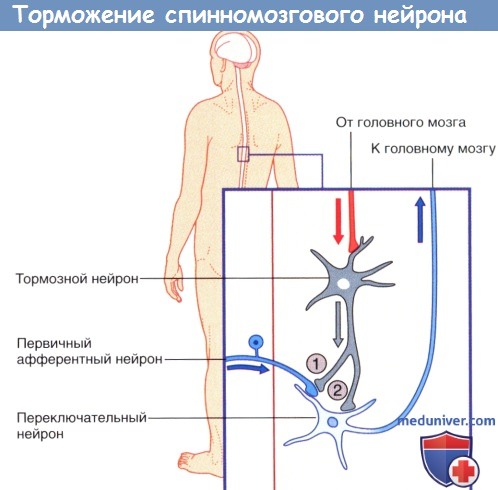 (1) Пресинаптическое и (2) постсинаптическое торможение спинномозгового нейрона, направляющегося к головному мозгу.
(1) Пресинаптическое и (2) постсинаптическое торможение спинномозгового нейрона, направляющегося к головному мозгу.
Стрелками указано направление проведения импульсов (возможно торможение переключательного нейрона под действием тормозных влияний). 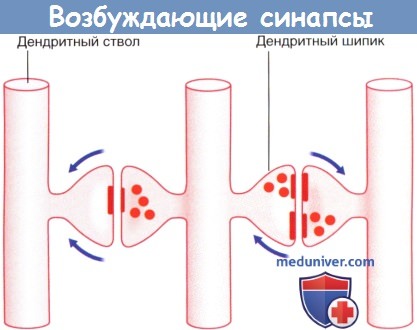 Возбуждающие дендро-дендритические синапсы. Изображены дендриты трех нейронов.
Возбуждающие дендро-дендритические синапсы. Изображены дендриты трех нейронов.
Реципрокный синапс (справа). Стрелками указано направление распространения электрото-нических волн.
Учебное видео — строение синапса
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
Источник



