
- Способ передачи наследственной информации называется
- Биология. 10 класс
- Хранение, передача и реализация наследственной информации в клетке. Ген. Геном. Реакции матричного синтеза
- Необходимо запомнить
- Реализация генетической информации
- Содержание
- Информация, содержащаяся в биологических последовательностях
- Универсальные способы передачи биологической информации
- Репликация ДНК (ДНК → ДНК)
- Транскрипция (ДНК → РНК)
- Трансляция (РНК → белок)
- Специальные способы передачи информации
- Обратная транскрипция (РНК — ДНК)
- Репликация РНК (РНК — РНК)
- Прямая трансляция белка на матрице ДНК (ДНК — белок)
- Эпигенетические изменения
- Прионы
- История возникновения термина «догма»
- См. также
- Примечания
- Ссылки
- Полезное
- Смотреть что такое «Реализация генетической информации» в других словарях:
Способ передачи наследственной информации называется
§ 15. ПЕРЕНОС ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ В КЛЕТКЕ:
РЕПЛИКАЦИЯ, ТРАНСКРИПЦИЯ
В процессе размножения клеток генетическая информация передается от одного поколения клеток другому. При этом все клетки получают одинаковую информацию. Это, возможно, вследствие того, что перед делением клетки осуществляется репликация (удвоение) ДНК, в результате образуются две идентичные молекулы ДНК, которые и передаются потомкам. В структуре ДНК заложена способность этой молекулы к копированию.
Закодированная в ДНК генетическая информация реализуется в результате экспрессии генов. Экспрессия генов включает транскрипцию (копирование информации с ДНК на синтезируемую РНК) и последующую трансляцию (синтез на матрице РНК соответствующего белка).
Возможен поток информации и в направлении от РНК к ДНК, этот процесс носит название обратная транскрипция. В то же время информация не передается от белков нуклеиновым кислотам. Однако следует отметить, что белки играют важную роль в осуществлении процессов передачи информации, как между нуклеиновыми кислотами, так и от нуклеиновых кислот к белкам.
Описанные информационные взаимоотношения между ДНК, РНК и белками могут быть представлены в виде схемы:
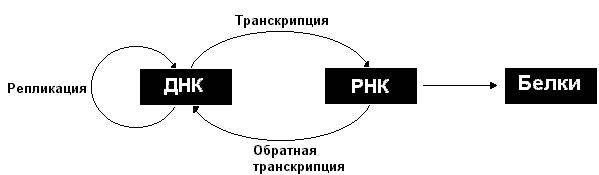
Рассмотрим более подробно процессы репликации, транскрипции и трансляции.
Интересно знать! В 1982 году Р.Д.Пальмитер с коллегами опубликовали фотографию, на которой рядом сидели две мыши. Одна из них была трансгенной, в ее ДНК встроили ген гормона роста крысы, другая была обычной мышкой. Трансгенная мышь в сравнении с обычной имела в два раза большие размеры. Причиной тому была экспрессия гена гормона роста, в результате которой у мыши синтезировался соответствующий гормон, который и определил ее гигантские размеры.
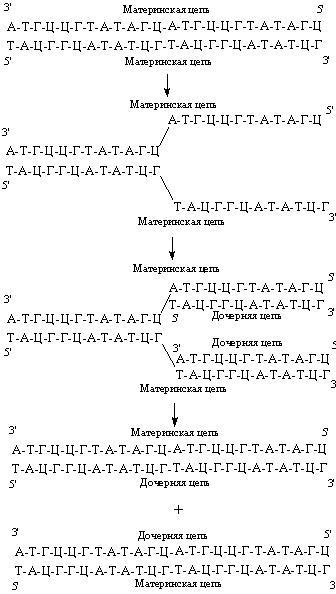
Рис. 44. В результате репликации образуются две дочерние молекулы ДНК.
Репликация ДНК
В основе репликации, или удвоения, ДНК лежит принцип комплементарности. Репликация начинается с разделения цепей, каждая из которых становится матрицей, определяющей нуклеотидную последовательность новой комплементарной цепи (рис. 44). В результате репликации образуются две дочерние молекулы ДНК, нуклеотидные последовательности которых идентичны между собой и с материнской молекулой ДНК. При этом две дочерние молекулы ДНК содержат одну вновь синтезированную (дочернюю) и одну материнскую цепи ДНК.
Субстратом для синтеза ДНК являются дезоксинуклеотид-5’-трифосфаты: дАТФ, дГТФ, дЦТФ и дТТФ. Уравнение этой реакции в простейшей виде выглядит так:
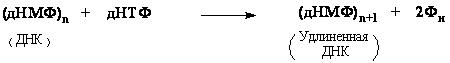
Данную реакцию катализирует фермент ДНК-полимераза. Она последовательно добавляет нуклеотиды к 3’-концу полинуклеотидной цепи. Следовательно, синтез новой цепи происходит в направлении от 5’-конца к 3’-концу. ДНК-полимераза может только наращивать цепь ДНК, начать же синтез ДНК с нуля она не может, т.е. для ее работы требуется цепь-затравка. ДНК-полимераза способна удлинять цепь только в присутствии цепи, играющей роль матрицы (рис. 45.). Нуклеотиды присоединяются к цепи-затравке в соответствии с принципом комплементарности, напротив аденина всегда будет встроен тимин, а напротив гуанина – цитозин.
Итогом репликации является образование двух дочерних молекул ДНК, являющихся точными копиями материнской. Благодаря этому возможна передача идентичной генетической информации от клетки к клетке.
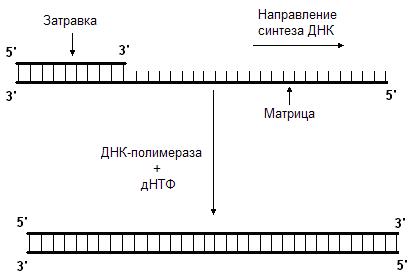
Рис. 45. ДНК-полимераза наращивает цепь в направлении 5’ 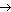 3’.
3’.
Для этого ей нужны матрица и затравка
Транскрипция
В результате транскрипции происходит синтез цепи РНК, нуклеотидная последовательность которой комплементарна одной из цепей ДНК; при этом генетическая информация, содержащаяся в ДНК, передается молекуле РНК. В результате транскрипции образуются три класса РНК: иРНК, тРНК, рРНК. Поскольку в основе транскрипции лежит принцип комплементарности, этот процесс имеет определенное сходство с репликацией. Однако между ними существует и важное различие. Если в процессе репликации копируется вся молекула ДНК, то при транскрипции транскрибируется только ее незначительная часть. Катализирует синтез РНК фермент РНК-полимераза. Транскрибируемый участок ДНК ограничен со стороны 3’-конца промотором, – участком с которым связывается РНК-полимераза, со стороны 5’-конца – терминатором – участком, в котором прерывается синтез РНК. Последовательность ДНК, ограниченная промотором и терминатором, представляет собой единицу транскрипции – транскриптон (рис. 46).
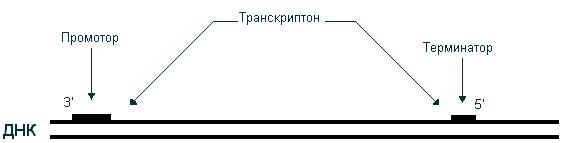
Рис. 46. Транскриптон
РНК-полимераза в качестве субстратов для синтеза РНК использует рибонуклеозид-5’-фосфаты (АТФ, ГТФ, ЦТФ и УТФ). Она удлиняет цепь, присоединяя нуклеотиды к 3’-концу, т.е. так же, как и ДНК-полимераза, наращивает цепь в направлении5’®3’. Этот фермент требует для своей работы в качестве матрицы одну из цепей ДНК. Из двух цепей ДНК транскрибируется только одна. В отличие от ДНК-полимеразы, РНК-полимераза не нуждается в затравке. Удлинение цепи РНК описывается уравнением:

Нуклеотиды присоединяются к цепи в соответствии с принципом комплементарности. Напротив аденина матричной цепи ДНК в растущую цепь РНК всегда будет встроен урацил, напротив гуанина – цитозин, напротив тимина – аденин и напротив цитозина – гуанин соответственно. Т.о., образовавшаяся в результате транскрипции РНК комплементарна матричной цепи ДНК (рис. 47).
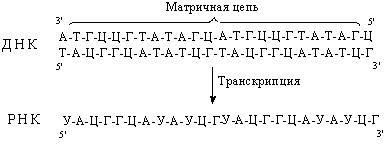
Рис. 47. Образовавшаяся в результате транскрипции РНК комплементарна матричной цепи ДНК
В процессе транскрипции выделяют три стадии: инициацию, элонгацию и терминацию. На стадии инициации РНК-полимераза, взаимодействуя с промотором, вызывает расхождение цепей ДНК и начинает синтез молекулы РНК. В ходе элонгации новосинтезированная цепь РНК образует (за счет спаривания ее оснований с основаниями матричной цепи ДНК) короткие отрезки гибридной двойной спирали ДНК – РНК, которые необходимы для правильного считывания цепи ДНК. Как только РНК-полимераза достигнет терминирующих последовательностей, запускается последняя стадия – терминация. По ее завершении происходит освобождение вновь синтезированной цепи РНК (рис. 48).
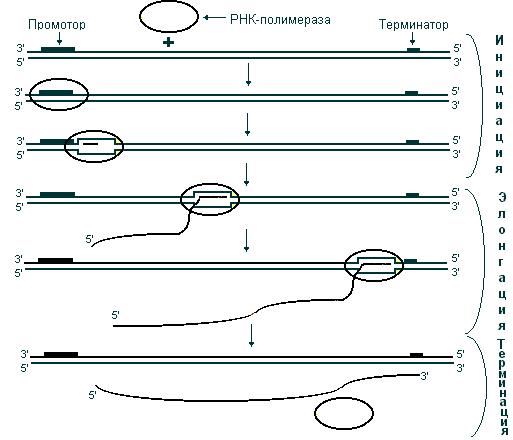
Рис. 48. Стадии транскрипции
Синтезированные молекулы РНК могут подвергаться посттранскрипционным ковалентным модификациям, так называемому процессингу (созреванию). Наиболее ярко процессинг выражен у эукариот.
Процессинг РНК
Рассмотрим процессинг иРНК эукариот.
иРНК синтезируется в виде предшественника пре-иРНК. Модификация пре-иРНК начинается на стадии элонгации, в это время происходит кэпирование 5’-конца. Остаток ГТФ присоединяется своим 5’-концом к 5’-концу пре-иРНК с образованием так называемого кэпа. После завершения транскрипции к 3’-концу присоединяется полиА-последовательность, состоящая из 100 – 200 последовательно соединенных адениновых нуклеотидов. В составе пре-иРНК могут содержаться интроны и экзоны. Интроны при созревании РНК вырезаются, концы экзонов соединяются друг с другом. Этот процесс называется сплайсингом. В результате сплайсинга зрелая молекула иРНК становится приблизительно в 4 раза короче пре-иРНК. Этапы процессинга иРНК представлены на рис. 49. Процессинг для иРНК прокариот не характерен.
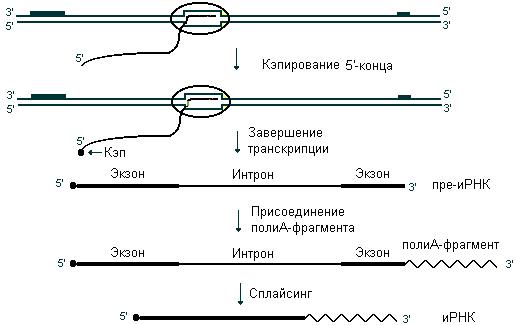
Рис. 49. Процессинг иРНК
рРНК и тРНК также синтезируются в виде более длинных предшественников, которые затем расщепляются и модифицируются. У прокариот рРНК представлены тремя молекулами: 16S-рРНК, 23S-рРНК и 5S-рРНК, у эукариот – четырьмя: 18S-рРНК, 28S-рРНК, 5,8S-рРНК и 5S-рРНК. рРНК как прокариот, так и эукариот образуются из предшественников – пре-рРНК. Предшественник расщепляется, образуя индивидуальные рРНК (рис. 50). рРНК и тРНК не содержат кэпа и полиА-последовательности. У эукариот рРНК и тРНК могут подвергаться сплайсингу. В процессе созревания у некоторых предшественников тРНК наряду с удалением концевых последовательностей может происходить и присоединение нуклеотидных последовательностей, играющих важную роль в их функционировании.
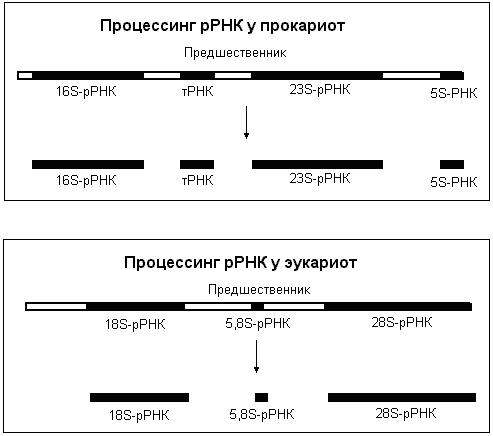
Рис.50. Процессинг рРНК.
Обратная транскрипция
Синтез ДНК при использовании в качестве матрицы РНК носит название обратной транскрипции. Данный процесс катализирует фермент обратная транскриптаза или ревертаза. Существование обратных транскриптаз в составе РНК-содержащих вирусов было показано Г.Темином и Д. Балтимором. Обнаружение обратной транскриптазы позволило ответить на вопрос: как генетическая информация РНК-содержащих вирусов может включиться в ДНК клетки-хозяина. Процесс обратной транскрипции, катализируемый ревертазой, и последующая интеграция генетического материала в геном клетки хозяина представлены на рис. 51. В процессе обратной транскрипции вначале образуется дуплекс РНК – ДНК, затем РНК в составе этого дуплекса разрушается, синтезированная цепь ДНК далее служит матрицей для синтеза второй цепи ДНК. На заключительной стадии происходит интеграция ДНК, синтезированной в результате обратной транскрипции, в ДНК клетки-хозяина. Обратные транскриптазы способны синтезировать ДНК, комплементарную самым различным РНК. Благодаря этой особенности ревертаза нашла широкое применение в научных исследованиях. С помощью обратной транскриптазы можно получить, например, искусственный ген, используя в качестве матрицы иРНК.
Источник
Биология. 10 класс
Генетическая информация в клетке
Хранение, передача и реализация наследственной информации в клетке. Ген. Геном. Реакции матричного синтеза
Необходимо запомнить
На Земле живёт около 7 млрд людей. Если не считать 25–30 млн пар однояйцовых близнецов, то генетически все люди разные: каждый уникален, обладает неповторимыми наследственными особенностями, свойствами характера, способностями, темпераментом.
Такие различия объясняются различиями в генотипах – наборах генов организма; у каждого он уникален. Генетические признаки конкретного организма воплощаются в белках – следовательно, и строение белка одного человека отличается, хотя и совсем немного, от белка другого человека.
Это не означает, что у людей не встречается совершенно одинаковых белков. Белки, выполняющие одни и те же функции, могут быть одинаковыми или совсем незначительно отличаться одной-двумя аминокислотами друг от друга. Но не существует на Земле людей (за исключением однояйцовых близнецов), у которых все белки были бы одинаковы.
Информация о первичной структуре белка закодирована в виде последовательности нуклеотидов в участке молекулы ДНК – гене – единице наследственной информации организма. Каждая молекула ДНК содержит множество генов. Совокупность всех генов организма составляет его генотип.
Кодирование наследственной информации происходит с помощью генетического кода, который универсален для всех организмов. Каждая аминокислота кодируется тремя нуклеотидами (триплетом) ДНК, комбинирующихся в разной последовательности (ААТ, ГЦА, АЦГ, ТГЦ и т.д. Аминокислот, входящих в состав белков – 20, а возможностей для комбинаций четырёх нуклеотидов в группы по три – 64, поэтому одна аминокислота может кодироваться несколькими триплетами. Часть триплетов вовсе не кодирует аминокислоты, а запускает или останавливает биосинтез белка.
ДНК непосредственно не принимает участия в биосинтезе белка. Информация с ДНК сначала копируется на иРНК (транскрипция), а затем на рибосомах переводится в последовательность аминокислот в молекулах синтезируемого белка (процесс трансляции).
В состав и-РНК входят нуклеотиды АЦГУ, триплеты которых называются кодонами: кодоны иРНК комплементарны триплетам ДНК: триплет на ДНК ЦГТ на и-РНК станет триплетом ГЦА, а триплет ДНК ААГ станет триплетом УУЦ.
Таким образом, генетический код – единая система записи наследственной информации в молекулах нуклеиновых кислот в виде последовательности нуклеотидов. Генетический код основан на использовании алфавита, состоящего всего из четырёх букв-нуклеотидов, отличающихся азотистыми основаниями: А, Т (У), Г, Ц.
Основные свойства генетического кода:
1. Генетический код триплетён. Триплет (кодон) – последовательность трёх нуклеотидов, кодирующая одну аминокислоту. Поскольку в состав белков входит 20 аминокислот, то очевидно, что каждая из них не может кодироваться одним нуклеотидом. Двух нуклеотидов для кодирования аминокислот также не хватает, поскольку в этом случае могут быть закодированы только 16 аминокислот. Значит, наименьшее число нуклеотидов, кодирующих одну аминокислоту, оказывается равным трём. (В этом случае число возможных триплетов нуклеотидов составляет 43 = 64).
2. Избыточность (вырожденность) кода является следствием его триплетности и означает то, что одна аминокислота может кодироваться несколькими триплетами (поскольку аминокислот 20, а триплетов – 64), за исключением метионина и триптофана, которые кодируются только одним триплетом. Кроме того, некоторые триплеты выполняют специфические функции: в молекуле иРНК триплеты УАА, УАГ, УГА – являются терминирующими кодонами, т. е. стоп-сигналами, прекращающими синтез полипептидной цепи. Триплет, соответствующий метионину (АУГ), стоящий в начале цепи ДНК, не кодирует аминокислоту, а выполняет функцию инициирования (начала) синтеза.
3. Одновременно с избыточностью коду присуще свойство однозначности: каждому кодону соответствует только одна определённая аминокислота.
4. Код коллинеарен, т. е. последовательность нуклеотидов в гене точно соответствует последовательности аминокислот в белке.
5. Генетический код неперекрываем. Это значит, что процесс считывания не допускает возможности перекрывания кодонов (триплетов).
6. Генетический код универсален, т. е. одинаковые для всех живых организмов вне зависимости от уровня организации и систематического положения этих организмов.
7. Генетический код содержит «знаки препинания» – стоп-кодоны. Начавшись на определённом кодоне, считывание идёт непрерывно триплет за триплетом вплоть до стоп-сигналов (терминирующих кодонов).
Источник
Реализация генетической информации
Центральная догма молекулярной биологии — обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Правило было сформулировано Френсисом Криком в 1958 году [1] и приведено в соответствие с накопившимися к тому времени данными в 1970 году [2] . Переход генетической информации от ДНК к РНК и от РНК к белку является универсальным для всех без исключения клеточных организмов, лежит в основе биосинтеза макромолекул. Репликации генома соответствует информационный переход ДНК->ДНК. В природе встречаются также переходы РНК->РНК и РНК->ДНК (например у некоторых вирусов), а также изменение конформации белков, передаваемое от молекулы к молекуле.
Содержание
Информация, содержащаяся в биологических последовательностях
Биополимеры — это синтезируемые живыми существами (биологические) полимеры. ДНК, РНК и белки относятся к линейным полимерам, то есть каждый входящий в их состав мономер соединяется с минимум двумя другими мономерами. Последовательность мономеров кодирует информацию, правила передачи которой описываются центральной догмой. Информация передаётся с высокой точностью, детерминистически и один биополимер используется как шаблон для сборки другого полимера с последовательностью, которая полностью определяется последовательностью первого полимера.
Универсальные способы передачи биологической информации
| Общие | Специальные | Неизвестные |
|---|---|---|
| ДНК → ДНК | РНК → ДНК | белок → ДНК |
| ДНК → РНК | РНК → РНК | белок → РНК |
| РНК → белок | ДНК → белок | белок → белок |
В живых организмах встречаются три вида гетерогенных, то есть состоящих из разных мономеров полимера — ДНК, РНК и белок. Передача информации между ними может осуществляться 3 х 3 = 9 способами. Центральная догма разделяет эти 9 типов передачи информации на три группы:
- Общий — встречающиеся у большинства живых организмов;
- Специальный — встречающиеся в виде исключения, у вирусов и у мобильных элементов генома или в условиях биологического эксперимента;
- Неизвестные — не обнаружены.
Репликация ДНК (ДНК → ДНК)
ДНК — основной способ передачи информации между поколениями живых организмов, поэтому точное удвоение (репликация) ДНК очень важна. Репликация осуществляется комплексом белков, которые расплетают хроматин, затем двойную спираль. После этого ДНК полимераза и ассоциированные с ней белки, строят на каждой из двух цепочек идентичную копию.
Транскрипция (ДНК → РНК)
Транскрипция — биологический процесс, в результате которого информация, содержащаяся в участке ДНК, копируется на синтезируемую молекулу информационной РНК. Транскрипцию осуществляют факторы транскрипции и РНК-полимераза. В эукариотической клетке первичный транскрипт (пре-иРНК) часто редактируется. Этот процесс называется сплайсингом.
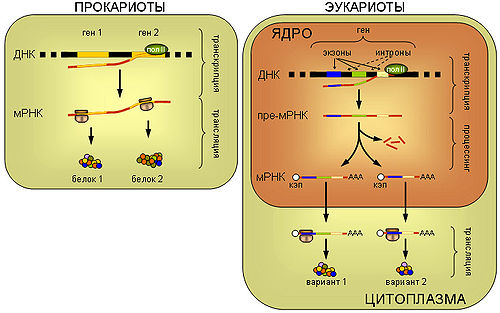
Трансляция (РНК → белок)
Зрелая иРНК считывается рибосомами в процессе трансляции. В прокариотических клетках процесс транскрипции и трансляции не разделён пространственно, и эти процессы сопряжены. В эукариотических клетках место транскрипции клеточное ядро отделено от места трансляции (цитоплазмы) ядерной мембраной, поэтому иРНК транспортируется из ядра в цитоплазму. иРНК считывается рибосомой в виде трёхнуклеотидных «слов». Комплексы факторов инициации и факторов элонгации доставляют аминоацилированные транспортные РНК к комплексу иРНК-рибосома.
Специальные способы передачи информации
Обратная транскрипция (РНК — ДНК)
Обратная транскрипция — перенос информации с РНК на ДНК, процесс, обратный нормальной транскрипции, осуществляемый ферментом обратной транскриптазой . Встречается у ретровирусов, например, ВИЧ и в случае ретротранспозонов.
Репликация РНК (РНК — РНК)
Репликация РНК — копирование цепи РНК на комплемлементарную ей цепь РНК с помощью фермента РНК-зависимой РНК-полимеразы. Вирусы, содержащие одноцепочечную (например, пикорнавирусы, к которым относится вирус ящура) или двуцепочечную РНК реплицируются подобным способом.
Прямая трансляция белка на матрице ДНК (ДНК — белок)
Прямая трансляция была продемонстрирована в клеточных экстрактах кишечной палочки, которые содержали рибосомы, но не иРНК. Такие экстракты синтезировали белки с введённых в систему ДНК, и антибиотик неомицин усиливал этот эффект. [3] [4]
Эпигенетические изменения
Эпигенетические изменения — это изменения в проявлении генов, не обусловленные изменением генетической информации (мутациями). Эпигенетические изменения происходят в результате модификации уровня экспрессии генов, то есть их транскрипции и/или трансляции. Наиболее изученным видом эпигенетической регуляции является метилирование ДНК с помощью белков ДНК-метилтрансфераз, что приводит к временной, зависящей от условий жизни организма инактивации метилированного гена. Однако поскольку первичная структура молекулы при этом ДНК не изменяется, это исключение нельзя считать истинным примером передачи информации от белка к ДНК.
Прионы
Прионы — белки, которые существуют в двух формах. Одна из форм (конформаций) белка является функциональной, обычно растворимой в воде. Вторая форма образует нерастворимые в воде агрегаты, часто в виде молекулярных трубочек-полимеров. Мономер — молекула белка — в этой конформации способен просоединяться к другим сходным молекулам белка, переводя их во вторую, прионоподобную, конформацию. У грибов такие молекулы могут передаваться по наследству. Но, как и в случае метилирования ДНК, первичная структура белка в данном случае остаётся прежней, и переноса же информации на нуклеиновые кислоты не происходит.
История возникновения термина «догма»
Хорас Джадсон (англ. Horace Judson ) написал в книге «Восьмой день творения»:
«Я считал, что догма — это идея, которая не подтверждена фактами. Видите?». И Крик воскликнул с удовольствием: «Я просто не знал, что означает „догма“. Я мог бы с тем же успехом назвать это „Центральной Гипотезой“ или — чем-нибудь еще. Догма была просто слоганом.» [5]
My mind was, that a dogma was an idea for which there was no reasonable evidence. You see?!» And Crick gave a roar of delight. «I just didn’t know what dogma meant. And I could just as well have called it the ‘Central Hypothesis,’ or — you know. Which is what I meant to say. Dogma was just a catch phrase
Кроме того, в автобиографической книге «Что за сумасшедший поиск» («What mad pursuit») Крик писал о выборе слова «догма» и о вызванных этим выбором проблемах:
«Я назвал эту идею центральной догмой, я подозреваю, по двум причинам. Я уже использовал слово гипотеза в гипотезе о последовательности, кроме того, я хотел предположить, что это новое допущение более центральное и сильное… Как оказалось, использование термина догма вызвало больше неприятностей, чем оно того стоило… Через много лет Жак Моно сказал мне, что по-видимому я не понимал, что подразумевается под словом догма, которая означает часть веры, не подлежащая сомнению. Я смутно опасался подобного значения слова, но поскольку я считал, что все религиозные убеждения не имеют основания, я использовал слово так, как понимал его я, а не большинство других людей, применив его к грандиозной гипотезе, которая, несмотря на внушаемое ею доверие, была основана на небольшом количестве прямых экспериментальных данных.»
I called this idea the central dogma, for two reasons, I suspect. I had already used the obvious word hypothesis in the sequence hypothesis, and in addition I wanted to suggest that this new assumption was more central and more powerful. . As it turned out, the use of the word dogma caused almost more trouble than it was worth. Many years later Jacques Monod pointed out to me that I did not appear to understand the correct use of the word dogma, which is a belief that cannot be doubted. I did apprehend this in a vague sort of way but since I thought that all religious beliefs were without foundation, I used the word the way I myself thought about it, not as most of the world does, and simply applied it to a grand hypothesis that, however plausible, had little direct experimental support.
См. также
Примечания
- ↑ Crick, F.H.C. (1958): On Protein Synthesis. Symp. Soc. Exp. Biol. XII, 139—163. (pdf, early draft of original article)
- ↑ Crick, F. (1970): Central Dogma of Molecular Biology. Nature 227, 561—563. PMID 4913914
- ↑ B. J. McCarthy and J. J. Holland (September 15, 1965). «Denatured DNA as a Direct Template for in vitro Protein Synthesis». Proceedings of the National Academy of Sciences of the United States54: 880–886. DOI:10.1073/pnas.54.3.880. PMID 4955657.
- ↑ . T. Uzawa, A. Yamagishi, T. Oshima (2002-04-09). «Polypeptide Synthesis Directed by DNA as a Messenger in Cell-Free Polypeptide Synthesis by Extreme Thermophiles, Thermus thermophilus HB27 and Sulfolobus tokodaii Strain 7″. The Journal of Biochemistry131: 849–853. PMID 12038981.
- ↑ The Eighth Day of Creation: Makers of the Revolution in Biology (25th anniversary edition). — 1996.
Ссылки
- B. J. McCarthy, J. J. Holland Denatured DNA as a Direct Template for in vitro Protein Synthesis // PNAS. — 1965. — Т. 54. — С. 880—886.
- Werner, E. Genome Semantics, In Silico Multicellular Systems and the Central Dogma. // FEBS Letters. — 2005. — В. 579. — С. 1779—1782. PMID 15763551
- Horace Freeland Judson Chapter 6: My mind was, that a dogma was an idea for which there was no reasonable evidence. You see?! // The Eighth Day of Creation: Makers of the Revolution in Biology (25th anniversary edition).. — 1996.
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое «Реализация генетической информации» в других словарях:
Генетика — I Генетика (греч. genētikos относящийся к происхождению) наука о наследственности и изменчивости организмов. Термин предложен в 1906 г. английским биологом Бейтсоном (W. Bateson). Способность организма обеспечивать в ряду поколений… … Медицинская энциклопедия
Молекулярная генетика — раздел генетики (См. Генетика) и молекулярной биологии (См. Молекулярная биология), ставящий целью познание материальных основ наследственности (См. Наследственность) и изменчивости (См. Изменчивость) живых существ путём исследования… … Большая советская энциклопедия
ГЕНЕТИЧЕСКИЙ КОД — свойственная живым организмам единая система записи наследств, информации в молекулах нуклеиновых к т в виде последовательности нуклеотидов; определяет последовательность включения аминокислот в синтезирующуюся полипептидную цепь в соответствии с … Биологический энциклопедический словарь
Ген — I (греч. genos род, происхождение) структурно функциональная единица генетического материала, наследственный фактор, который можно условно представить как отрезок молекулы ДНК (у некоторых вирусов молекулы РНК), включающий нуклеотидную… … Медицинская энциклопедия
Генетическая информация — информация о строении белков, закодированная с помощью последовательности нуклеотидов генетического кода в генах (особых функциональных участках молекул ДНК или РНК). Генетическая информация определяет морфологическое строение, рост, развитие,… … Википедия
ЯДРО (в биологии) — ЯДРО (клеточное ядро), в биологии обязательная часть клетки у многих одноклеточных и всех многоклеточных организмов. Размеры от 1 мкм (у некоторых простейших) до 1 мм (в яйцах некоторых рыб и земноводных). Все организмы нашей биосферы как… … Энциклопедический словарь
Ген — У этого термина существуют и другие значения, см. Ген (значения). Схема транскрипции ДНК. Ген (др. греч … Википедия
Атабеков, Иосиф Григорьевич — Иосиф Григорьевич Атабеков … Википедия
Биология развития — онтогенетика, раздел биологии, всесторонне изучающий процессы и движущие силы индивидуального, или онтогенетического, развития организма. Б. р. преемник таких ранее возникших отраслей исследования Онтогенеза, как Механика развития, или… … Большая советская энциклопедия
Генетический код — Генетический код свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов. В ДНК используется четыре азотистых основания аденин (А), гуанин (G), цитозин… … Википедия
Источник



