
Вопрос 47. Способы передачи генетической информации
Быстрая реакция на изменения условий окружающей среды и приобретение необходимых генетических признаков (напр. изменение вирулентных свойств, устойчивость к АМП). Обмен генетической информацией (ОГИ)- механизм, обеспечивающий многообразие микробного мира.
ОГИ способствует быстрой адаптации бактерий.
ОГИ приводит к образованию рекомбинантной ДНК.
ОГИ служит важным фактором эволюции бактерий.
3 способа передачи генетической информации:
1. Конъюгация – внедрение ДНК из бактерии – донора в клетку реципиента
2. Трансформация – поглощение свободной ДНК из внешней среды
3. Трансдукция – распространение генов бактерий умеренными фагами
Конъюгация
Конъюгация – однонаправленный перенос генетического материала (хромосомной и плазмидной ДНК) от донора к реципиенту при непосредственном контакте клеток.
Донор – бактерия, содержащая конъюгативную плазмиду или конъюгативный транспозон, способная передавать ДНК реципиенту путем горизонтального переноса генов.
Реципиент – любая бактериальная клетка, способная принимать ДНК от бактерии-донора путем горизонтального переноса генов.
Механизм передачи конъюгативных плазмид между Гр- бактериями:
1. Образование межклеточного контакта — конъюгативных пилей
2. Сайт-специфическое разрезание одной цепочки ДНК
3. Сборка особой структуры — релаксосомы для переноса ДНК
4. Перемещение разделенных цепей ДНК в клетку реципиента
5. Синтез комплементарных цепей ДНК
6. Терминация переноса ДНК и разделение конъюгирующих клеток
Трансформация
Трансформация —поглощение бактерией ДНК из окружающей среды и включение в свой геном, при этом бактерия становится генетически трансформированной.
Впервые этот феномен описан у Streptococcus pneumoniae, когда непатогенные живые мутантные клетки стали патогенными для мышей при одновременной инъекции с убитыми нагреванием вирулентными клетками S.pneumoniae.
Изменение происходило в результате трансформирующего действия ДНК из убитых нагреванием вирулентных клеток S.pneumoniae.
Схема трансформации
1. Адсорбция ДНК на клетке (клеточной стенке)
2. Проникновение внутрь клетки
3. Рекомбинация с ДНК клеточной хромосомы
Трансдукция
Трансдукция — обмен генетическим материалом с участием бактериофагов.
Бактериофаги – вирусы бактерий — молекулярные паразиты, использующие для самовоспроизводства аппарат репликации ДНК и синтеза белка зараженных ими клеток.
Трансдукцию осуществляют бактериофаги, содержащие невирусную генетическую информацию. Подобно плазмидам, бактериофаги могут выступать в качестве векторов переноса ДНК от клеток-хозяев. Перенос фаговой информации может служить механизмом обмена генетическим материалом между бактериями. Бактериофаги E.coli
Виды трансдукции:
1. Специфическая трансдукция – перенос генов клетки-хозяина из строго определенных участков хромосомы бактериальной клетки. (Например, 5 из 11 фагов возбудителя дифтерии Corynebacterium diphtheriae – переносчики генов дифтерийного токсина).
2.Неспецифическая трансдукция (общая) — случайный перенос различных генов клетки-хозяина, расположенных в разных участках генома бактерии..Схема трансдукции 
Вопрос 48. Размножение бактерий (клеточный цикл)
У всех живых организмов рост клеток – это увеличение массы и последующее деление с образованием двух идентичных клеток. Не являются исключением и бактерии.
У бактерий генетический материал — в ковалентно замкнутой кольцевой молекуле ДНК.
Молекула ДНК – хромосома. Хромосома расположена в нуклеоиде.
Деление клетки Neisseria gonorrhoeae
Период от деления до деления называется вегетативным клеточным циклом (ВКЦ), он включает несколько этапов:
Репликация ДНК – удвоение генетического материала
Расхождение двух наборов хромосом
Деление клетки
Особенности клеточного цикла прокариот: ВКЦ прокариот и эукариот во многом сходен. Однако есть отличие: Во время быстрого роста в одной бактериальной клетке может происходить 2 — 3 цикла репликации хромосом одновременно.
Источник
Передача генетической информации у бактерий
Передача генетического материала от клетки-донора в клетку-реципиент путем непосредственною контакта клеток называется конъюгацией.
Передача генетического материала от клетки-донора в клетку-реципиент впервые была обнаружена Дж. Ледербергом и Э. Тейтумом в 1946 г.
Необходимым условием для конъюгации является наличие в клетке-доноре трансмиссивной плазмиды.
Трансмиссивные плазмиды кодируют половые пили, образующие конъюгационную трубочку между клеткой-донором и клеткой-реципиентом, по которому плазмидная ДНК передается в новую клетку. Механизм передачи плазмидной ДНК из клетки в клетку заключается в том, что специальный белок, кодируемый tra-опероном, «узнает» определенную последовательность в ДНК плазмиды (называемую origin — начало, англ., перено-
са), вносит в эту последовательность однопе-почечный разрыв и ковалентно связывается с 5′-концом. Затем цепь ДНК, с которой связан белок, переносится в клетку-реципиент, а неразорванная комплементарная цепь остается в клетке-доноре. Клеточный аппарат синтеза ДНК достраивает одиночные цепи и в доноре и в реципиенте до двухцепочечной структуры. Белок связанный с 5′-концом перенесенной цепи, способствует замыканию плазмиды в реципиентной клетке в кольцо. Этот процесс представлен на рис. 5.4, 1А на примере переноса в реципиентную клетку плазмиды F
клетка-реципиент приобретает донорские свойства (см. рис. 5.4, 1А).
Если F-фактор или другая трансмиссивная плазмида встраиваются в хромосому клетки-донора, то плазмида и хромосома начинают функционировать в виде единого трансмиссивного репликона, что делает возможным перенос бактериальных генов в бесплазмидную клетку-реципиент, т. е. процесс конъюгации. Штаммы, в которых плазмида находится в интегрированном состоянии, переносят свои хромосомные гены бесплазмидным клеткам с высокой частотой и поэтому называются Hfr (от англ. high frequency of recombination — высокая частота рекомбинации)
Процесс переноса хромосомных генов в случае скрещивания: Hfr xF
всегда начинается с расщепления ДНК в одной и той же точке, месте интеграции F-фактора или другой трансмиссивной плазмиды. Одна нить донорской ДНК передается через конъюгационный мостик в реципиентную клетку. Процесс сопровождается достраиванием комплементарной нити до образования двунитчатой структуры. Перенос хромосомных генов при конъюгации всегда имеет одинаковую направленность, противоположную встроенной плазмиде. Сама трансмиссивная плазмида передается последней. Переданная в реципиентную клетку и до-
строенная до двунитчатой структуры нить ДНК донора рекомбинирует с гомологичным участком реципиентной ДНК с образованием стабильной генетической структуры. Вследствие хрупкости конъюгационного мостика половой фактор редко передается в клетку-реципиент, поэтому образовавшиеся рекомбинант донорскими функциями как правило не обладает.
Вследствие направленности передачи генов конъюгация используется для картирования генома бактерий и построения генетической карты.
Трансдукцией называют передачу бактери-альной ДНК посредством бактериофага.
Этот процесс был открыт в 1951 г. Н. Циндером и Дж. Ледербергом. В процессе репликации фага внутри бактерий (см. разд. 3.5) фрагмент бактериальной ДНК проникает в фаговую частицу и переносится в реципиен-тную бактерию во время фаговой инфекции. Существует два типа трансдукции:
общая трансдукция — перенос бактериофагом фрагмента любой части бактериальной хромосомы — происходит вследствие того, что бактериальная ДНК фрагментируется после фаговой инфекции и кусочек бактериальной ДНК того же размера, что и фаговая ДНК, проникает в вирусную, формируя дефектную фаговую частицу с частотой приблизительно 1 на 1000 фаговых частиц (рис. 5.4, 2А). При инфицировании клетки-реципиента дефектной фаговой частицей ДНК клетки-донора «впрыскивается» в нее и рекомбинирует гомологичной рекомбинацией с гомологичным участком хромосомы-реципиента с образованием стабильного рекомбинанта. Этим типом трансдукции обладают Р-фаги;
специфическая трансдукция — наблюдается в том случае, когда фаговая ДНК интегрирует в бактериальную хромосому с образованием профага. В процессе исключения ДНК-фага из бактериальной хромосомы в результате случайного процесса захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы, становясь дефектным фагом (рис. 5.4, 2Б). Так как большинство умеренных бактериофагов
интегрирует в бактериальную хромосому в специфических участках, для таких бактериофагов характерен перенос в клетку-реципиент определенного участка бактериальной ДНК клетки-донора. ДНК дефектного фага рекомбинирует с ДНК клетки-реципиента сайт-специфической рекомбинацией. Рекомбинант становится меродиплоидом по привнесенному гену. В частности, бактери-офагпередает специфической трансдукцией gal-ген у Е. coli.
Феномен трансформации впервые был описан в 1928 г. Ф. Гриффитсом, обнаружившим превращение бескапсульного R-штамма пневмококков (Streptococcus pneumoniae) в штамм, образующий капсулу S-формы. Гриффите ввел мышам одновременно небольшое количество авирулентных R-клеток и убитых нагреванием S-клеток. R-клетки были получены от штамма, капсульное вещество которого принадлежало к типу S II, а убитые нагреванием S-штаммы принадлежали к типу SIII. Из крови погибших мышей были выделены вирулентные пневмококки с капсулой S III.
Природу трансформирующего фактора в 1944 г. установили О. Эвери, К. Мак-Леод, М. Мак-Карти, которые показали, что ДНК, экстрагированная из инкапсулированных пневмококков, может трансформировать не-капсулированные пневмококки в инкапсулированную форму. Таким образом, было доказано, что именно ДНК является носителем генетической информации.
Процесс трансформации может самопроизвольно происходить в природе у некоторых видов бактерий, чаще у грамположительных, когда ДНК, выделенная из погибших клеток, захватывается реципиентными клетками.
Процесс трансформации зависит от компетентности клетки-реципиента и состояния донорской трансформирующей ДНК. Компетентность — это способность бактериальной клетки поглощать ДНК. Она зависит от присутствия особых белков в клеточной мембране, обладающих специфическим аффинитетом к ДНК. Состояние компетентности у грамположительных бактерий связано с определенными фазами кривой роста.
Трансформирующей активностью обладает только двунитчатая высокоспирализо-ванная молекула ДНК.
Это связано с тем, что в клетку-реципиент проникает только одна нить ДНК, тогда как другая — на клеточной мембране — подвергается деградации с освобождением энергии, которая необходима для проникновения в клетку сохранившейся нити. Высокий молекулярный вес трансформирующей ДНК увеличивает шанс рекомбинации, так как внутри клетки трансформирующая нить ДНК подвергается воздействию эндонуклеаз. Интеграция с хромосомой требует наличия гомологичных с ней участков у трансформирующей ДНК. Рекомбинация происходит на одной нити, в результате чего образуется гетеродуплексная молекула, одна нить которой имеет генотип реципиента, а другая — рекомбинантный генотип. Рекомбинантные трансформанты формируются только после цикла репликации (рис. 5.4, 3).
В настоящее время этот метод является основным методом генной инженерии, используемым при конструировании рекомбинант-ных штаммов с заданным геномом.
Дата добавления: 2016-02-04 ; просмотров: 2520 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник
Общая характеристика способов обмена генетической информацией у бактерий
У бактерий, как и у высших организмов, существуют механизмы, которые обеспечивают перекомбинацию генетического материала между родственными, но генетически неидентичными клетками. В результате такого обмена возникает потомство с новой комбинацией наследственных признаков – рекомбинанты. Однако способы обмена генетической информацией и процессы формирования рекомбинантов у бактерий отличаются от существующих у эукариот, что вносит своеобразие в рекомбинационный анализ генетической организации бактериальной хромосомы.
Известны три механизма передачи генетического материала у бактерий: конъюгация, трансформация и трансдукция.
Конъюгация – это направленный перенос генетического материала из клетки-донора в клетку-реципиент при непосредственном физическом контакте между ними.
Трансформация – способ передачи генетической информации от клеток-доноров к клеткам-реципиентам с помощью химически чистой ДНК.
Трансдукция – это перенос генетического материала из клеток-доноров в клетки-реципиенты с помощью бактериофагов.
Основная отличительная черта всех трех механизмов – это однонаправленный перенос от одной клетки (донора) к другой (реципиенту) части генетического материала (рис.5). Вследствие такого обмена возникает не полноценная диплоидная зигота, а частично диплоидная (меродиплоидная) зигота, или, так называемая, мерозигота. В мерозиготе генетический материал реципиентной клетки представлен целой хромосомой, а материал донорной клетки только фрагментом хромосомы или частью генома.
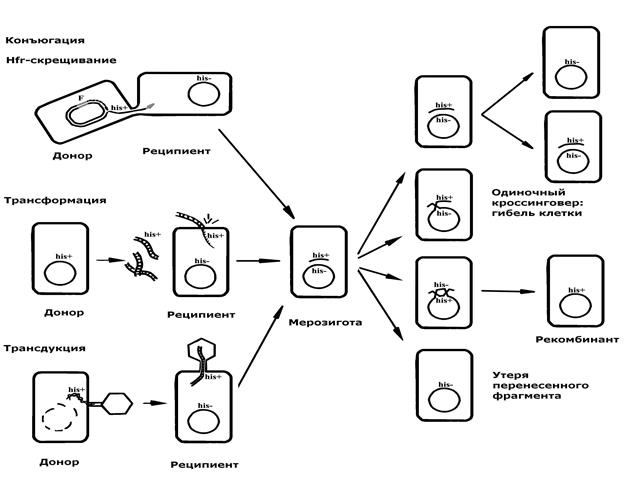 |
Рис. 5. Механизмы передачи генетического материала у бактерий и судьба перенесенного фрагмента
Судьба перенесенного фрагмента донорной ДНК в мерозиготе неоднозначна и зависит от многих факторов. Обычно он не способен к самостоятельной редупликации в клетке реципиента, так как не является репликоном, но некоторое время может в ней сохраняться и даже функционировать (транскрибироваться). В этом случае фрагмент ДНК донора при каждом делении мерозиготы будет передаваться лишь одной из дочерних клеток (однолинейная передача). Однако такое состояние не стабильно и может привести либо к утере фрагмента, либо к включению его в хромосому реципиентной клетки за счет рекомбинации (кроссинговера или конверсии) с ее гомологичной последовательностью.
Важно отметить ту особенность рекомбинации у бактерий, что только четное число обменов между хромосомой реципиента и переданным фрагментом донора дает полноценную хромосому. Единичный или нечетный кроссинговер приводит к размыканию хромосомы и дупликации концевых фрагментов (рис.5). Следовательно, элементарным регистрируемым актом рекомбинации в мерозиготе является двойной кроссинговер. Результатом двойного (или четного) обмена является реципрокная интеграция донорного фрагмента в реципиентную хромосому и участка реципиентной хромосомы в донорный фрагмент. Так как донорный фрагмент обычно не реплицируется и в дальнейшем теряется, регистрируется только интеграция в хромосому. Включенный фрагмент (или его участок) будет реплицироваться совместно с хромосомой реципиента и передаваться потомству. В результате в популяции реципиентных клеток появятся стабильные рекомбинанты, получившие некоторые признаки донора. Отмеченная особенность рекомбинации приводит к нереципрокности и уменьшенному вдвое выходу рекомбинантов.
Следует также напомнить, что бактериальные клетки в различных условиях могут содержать несколько копий геномных молекул ДНК. Поэтому рекомбинанты появляются не сразу, а после 3-4 последовательных делений, когда клетки становятся гаплоидными (несут одинаковые копии).
Размер перенесенного фрагмента хромосомы донора зависит от способа передачи генетического материала. Например, при трансдукции он ограничен объемом головки бактериофага, в которую может быть ошибочно упакована ДНК клетки-хозяина. При трансформации длина переносимой молекулы очень мала и зависит от способа приготовления препарата ДНК и специфики конкретной системы. Только при конъюгации размер переносимого генетического материала может значительно колебаться и в редких случаях достигать размера полного генома.
Дата добавления: 2015-02-07 ; просмотров: 2402 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник



