
Образование и дифференцировки мезодермы у птиц.
Образование и диф-ка мезодермы у птиц.
У всех животных, за искл к/п , в связи с гаструляцией (параллельно ей или на след этапе, обусловленном гаструляцией) возникает и третий зародышевый пласт — мезодерма.
Это совокупность клет эл-ов, залегающих м/у экто- и энтодермой те в бластоцеле. Зародыш становится 3х слойным. У высших позвоночных трехслойное строение зародыша возникает уже в процессе гастр ( у низш в процессе гастр образ 2х слойный зар)
2 пути: Телобластический (Protostomia) и энтероцельный (Deuterostomia).
При энтероцельном способе совокупность клеток формирующей меходермы появляется в виде карманоподобных выступов первичного кишечника (выпячивание его стенок внутрь бластоцеля). Выступы, внутрь которых входят участки первичной кишечной полости, обособляются от кишечника и отделяются от него в виде мешочков. Полость мешочков превращается в целом те во вторичную полость тела, целомические мешки могут подразделяться на сегменты.
Особенности у птиц: Яйцевые кл-ки гигантские, с большим кол-вом желтка. Дробится лишь небольшой уч-к яйца, расположенный в районе анимального полюса. Первые бластомеры распластаны на пов-ти желтка. После 2-3 деления бластомеры делятся асинхронно. В центре формирующегося бластодиска появляются борозды, проходящие в плоскости, параллельной пов-ти яйца. Возникают изолированные от желточных масс клетки. Количество клеток бластодиска увеличивается, а под ним появляется подзародышевая полость. Вся средняя часть бластодиска расположена над зародышевой полостью и не соприкасается с желтком — светлая зона. Зона клеток, прилегающих к желтку — темная зона.
В средней части клетки сгущаются, располагаются плотнее — зародышевый щиток. В его задней части, на стороне, обращенной к его полости, начинается обособление путем деламинации внутреннего слоя клеток — первичного гипобласта (внезар энтодерма).
Позднее на заднем конце зародышевого щитка образуется второе поколение клеток гипобласта — вноричный гипобласт, который вытесняет первичный к переферии бластодиска.
Дальше, в области зародышевого щитка образуется первичная полоска(вдоль средней линии зоны). Она удлиняется и занимает 2/3 зоны. На ее переднем конце образуется утолщение — гензеновский узелок, в цетре которого появится ямка. По всей первичной полоске формируется первичная бороздка. До образования узелка первичная полоска — источник формирования кишечной энтодермы зародыша, которая мигрирует под эпибласт, распостраняется латерально. Со дна первичной бороздки мигрируют внутрь клетки мезодермы. Впереди гензеновского узелка в составе эпибласта располагаются клетки, которые погружаются внутрь (около узелка) и оказываются лежащими под эпибластом. (Гензеновский узелок- гомолог дорсальной губы бластопора амфибий, а материал, расположенный спереди — презумптивная хорда. Правые и левые края первичной бороздки — гомологи боковых губ бластопора амфибий).
Дата добавления: 2016-05-05 ; просмотров: 1764 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник
Эмбриональное развитие
От момента образования зиготы и до выхода зародыша из яйцевых оболочек длится эмбриональный период развития.
Дробление зиготы
После того, как произошло оплодотворение — слияние сперматозоида и яйцеклетки, образовавшаяся зигота начинает интенсивно делиться. Ее множественные митотические деления называют дроблением.
Важная особенность дробления в том, что не происходит увеличение в размере зародыша: клетки дробятся (делятся) настолько быстро, что не успевают накопить цитоплазматическую массу. Дробление зиготы человека является полным неравномерным асинхронным.
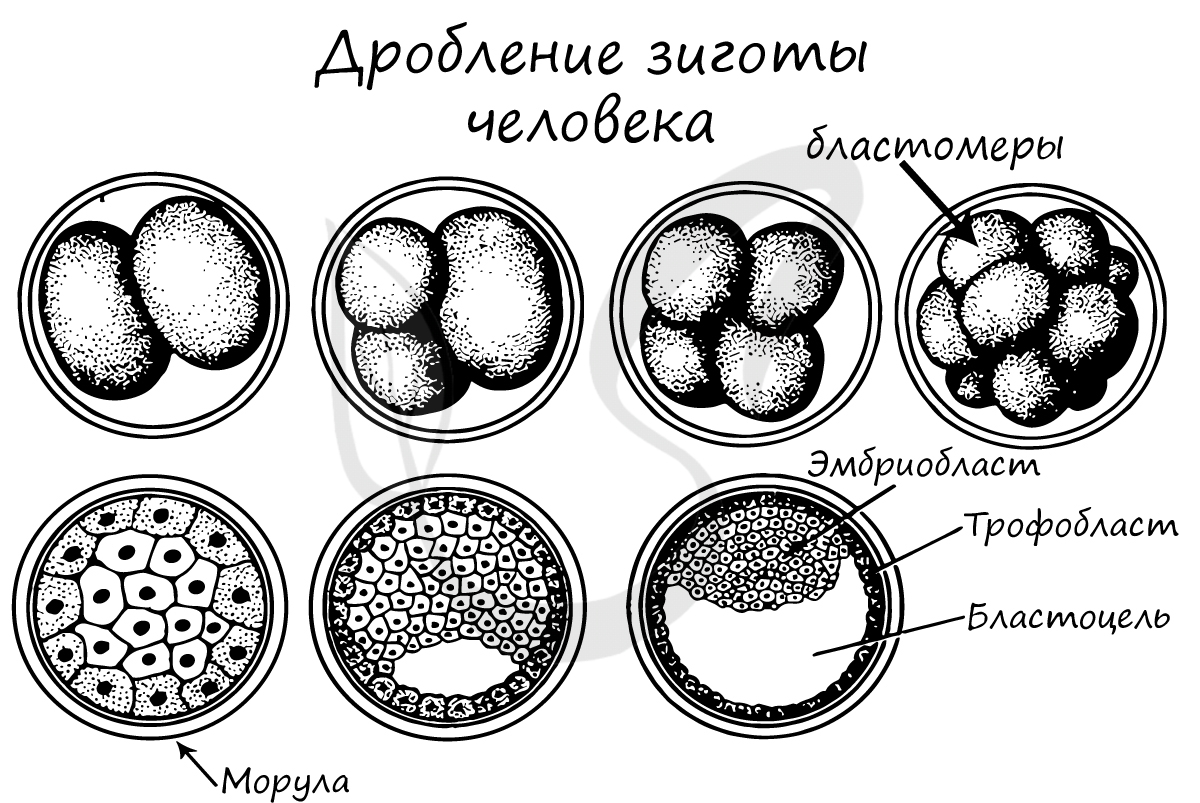
В результате дробления образуется морула. Морула (лат. morum — ягода тутового дерева) — клетка на стадии этапа дробления, когда зародыш представляет собой компактную совокупность клеток (без полости внутри).
Бластуляция
Бластуляция — заключительный период дробления, в который зародыш называется бластулой.
После очередных этапов многократного деления образуется однослойный зародыш с полостью внутри — бластула (греч. blastos — зачаток).
Стенки бластулы состоят из бластомеров, которые окружают центральную полость — бластоцель (греч. koilos — полый). Соединяясь друг с другом, бластомеры образуют бластодерму из одного слоя клеток.

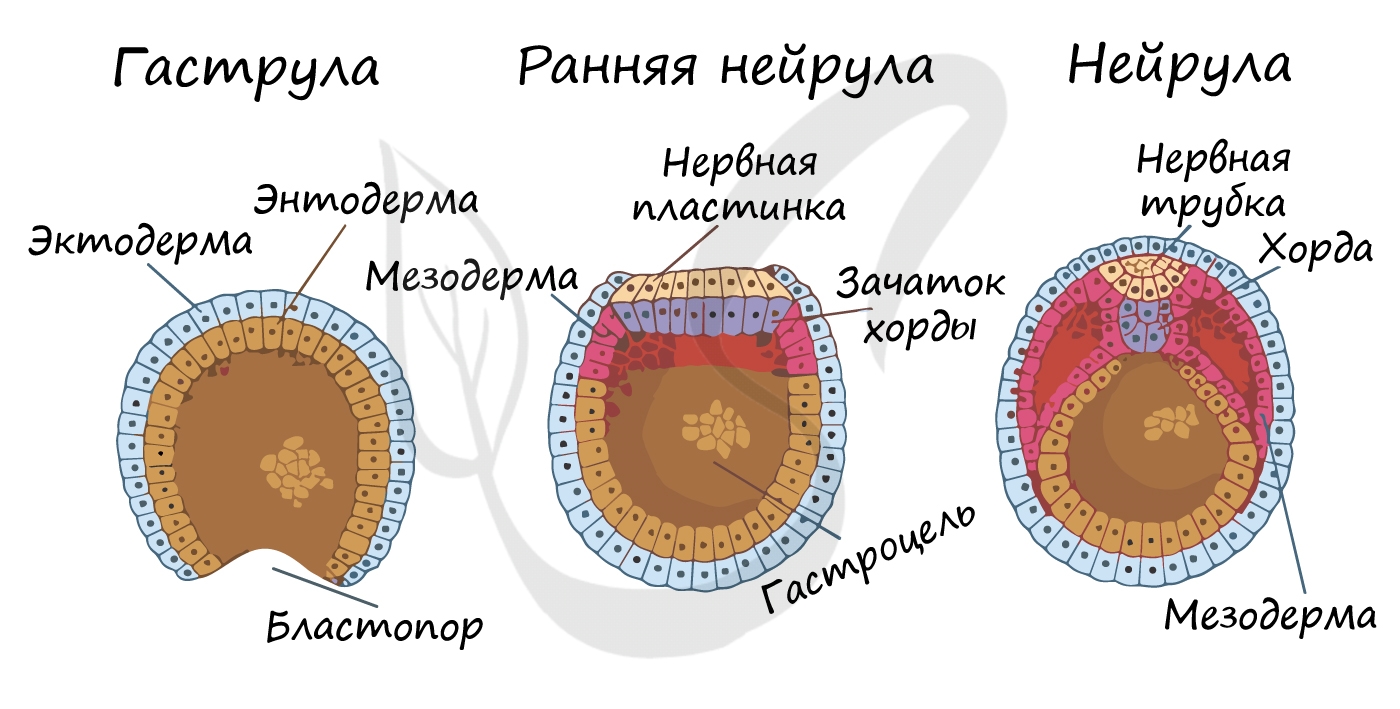
Гаструляция (греч. gaster — желудок, чрево)
Гаструляцией называют стадию эмбрионального развития, в ходе которой клетки, возникшие в результате дробления зиготы, формируют три зародышевых листка: эктодерму, мезодерму и энтодерму.
Стенка бластулы начинается впячиваться внутрь — происходит инвагинация стенки. По итогу такого впячивания зародыш становится двухслойным. Двухслойный зародыш называется — гаструла. Полость гаструлы называется гастроцель (полость первичной кишки), а отверстие, соединяющее гастроцель и внешнюю среду — первичный рот (бластопор).

У первичноротых животных на месте первичного рта (бластопора) образуется ротовое отверстие. К первичноротым относятся: кишечнополостные, плоские, круглые и кольчатые черви, моллюски, членистоногие.
У вторичноротых на месте бластопора формируется анальное отверстие, а ротовое отверстие образуется на противоположном полюсе. К вторичноротым относят хордовых и иглокожих (морских звезд, морских ежей).

При впячивании части бластулы (инвагинации) клетки бластодермы мигрируют внутрь и становятся энтодермой (греч. entós — внутренний). Оставшаяся часть бластодермы снаружи называется эктодермой (греч. ἔκτος — наружный).
Между энто- и эктодермой из группы клеток формируется третий зародышевый листок — мезодерма (греч. μέσος — средний).

Нейрула
Эта стадия следует за гаструлой. Ранняя нейрула представляет собой трехслойный зародыш, состоящий из энто-, экто- и мезодермы. На этапе нейрулы происходит закладка отдельных органов.
Важно отметить, что на стадии нейрулы происходит процесс нейруляции — закладывание нервной трубки. Нервная пластинка, образовавшаяся на ранних этапах, прогибается внутрь, при этом ее края сближаются и, замыкаясь, формируют нервную трубку.

Итак, как уже было сказано, на стадии нейрулы закладываются отдельные органы. Эктодерма образует покровный эпителий и нервную пластинку, мезодерма (из которой в дальнейшем появятся все соединительные ткани), энтодерма — окружает полость первичной кишки (гастроцель), образуя кишечник. От энтодермы отшнуровывается хорда.
Все три зародышевых листка требуют нашего особого внимания, а также понимания того, какие органы и структуры из них образуются.
Эктодерма (греч. ἔκτος — наружный) — наружный зародышевый листок, образует головной и спинной мозг, органы чувств, периферические нервы, эпителий кожи, эмаль зубов, эпителий ротовой полости, эпителий промежуточного и анального отделов прямой кишки, гипофиз, гипоталамус.
Мезодерма (греч. μέσος — средний) — средний зародышевый листок, образует соединительные ткани: кровеносную и лимфатическую системы, костную и хрящевую ткань, мышечные ткани, дентин и цемент зубов, а также выделительную (почки) и половую системы (семенники, яичники).
Энтодерма (греч. entós — «внутренний») — внутренний зародышевый листок, образует эпителий пищевода, желудка, кишечника, трахеи, бронхов, легких, желчного пузыря, мочевого пузыря и мочеиспускательного канала, печень и поджелудочную железу, щитовидную и паращитовидную железы.

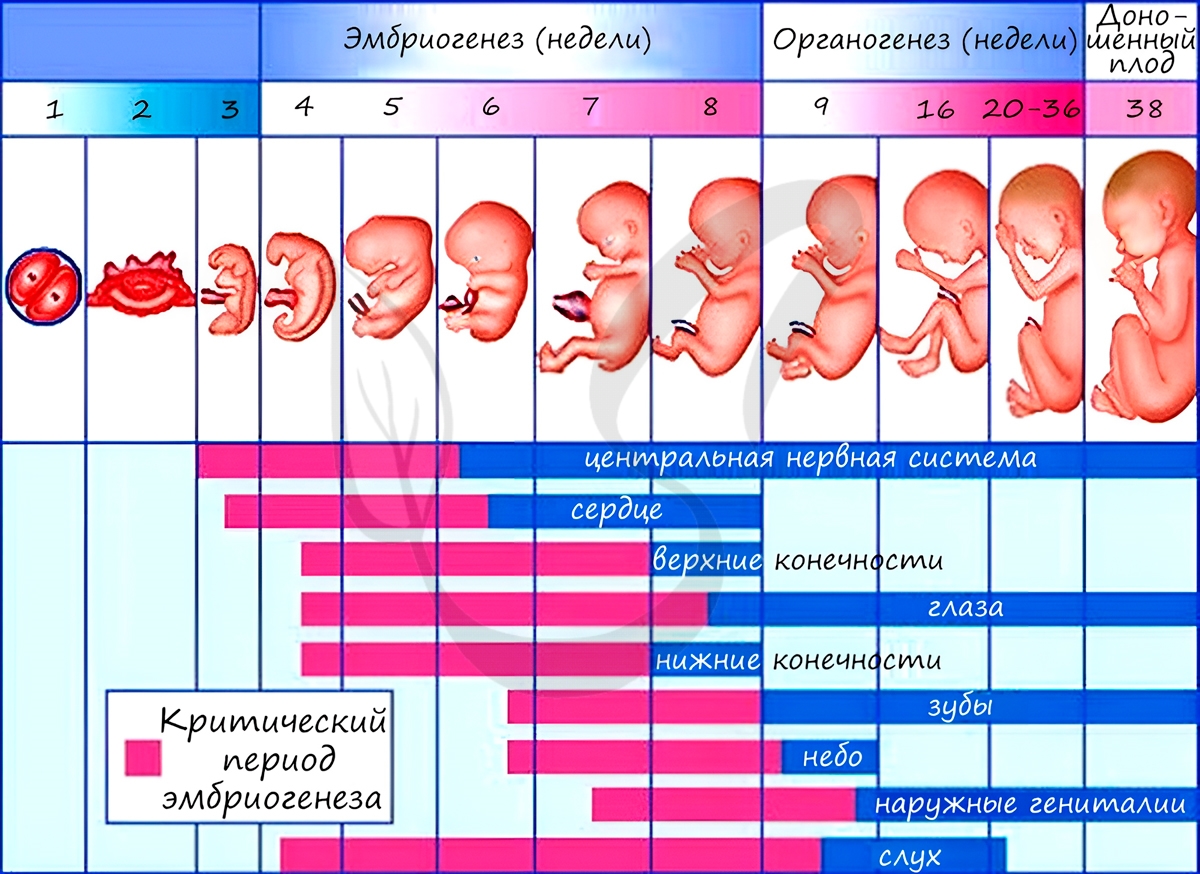
Из зародышевых листков образуются ткани, органы и системы органов. Такой процесс называется органогенезом. В период закладки органов важное значение имеет воздержание матери от вредных привычек (алкоголь, курение), которые могут нарушить процесс дифференцировки клеток и привести к тяжелейшим аномалиям, уродствам плода.
Некоторые лекарства также могут оказывать на плод тератогенный эффект (греч. τέρας — чудовище, урод), приводя к развитию уродств. Периоды закладки органов и система органов вследствие их большой важности носят название критических периодов эмбриогенеза.
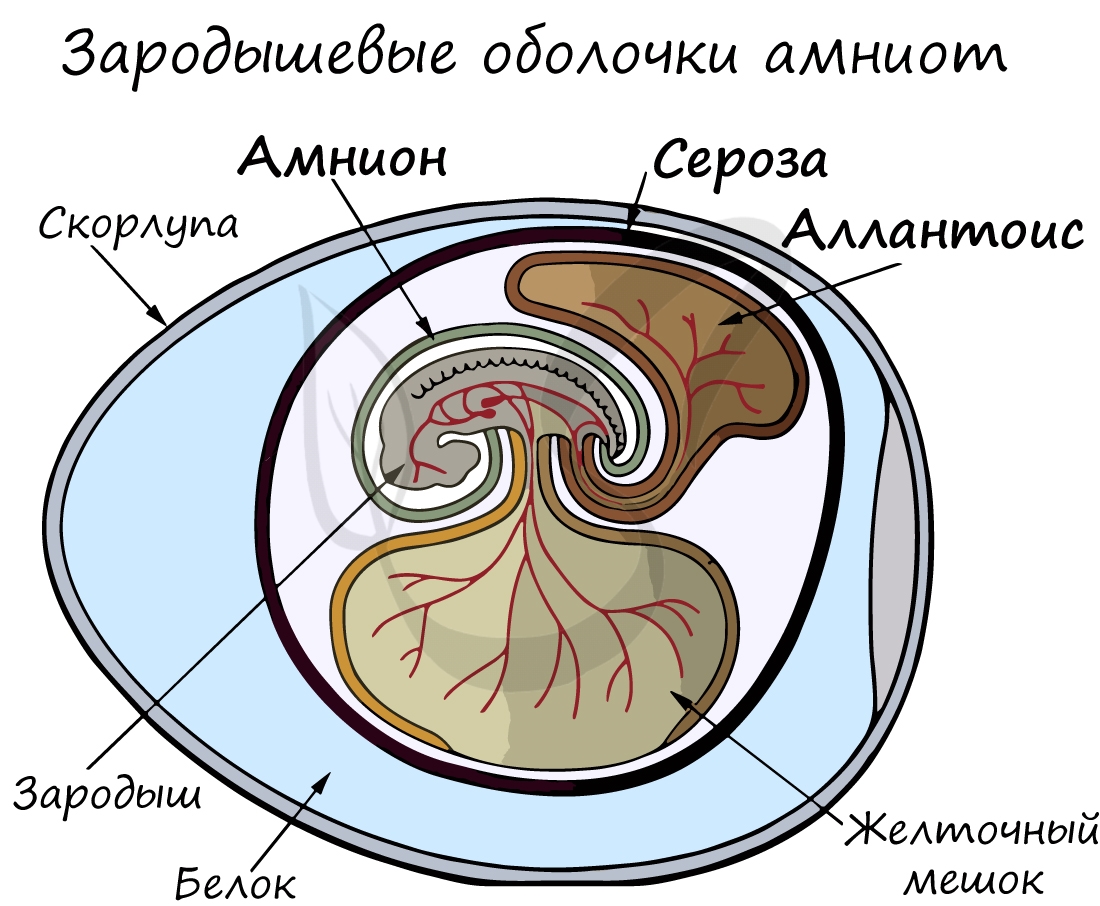
Анамнии и амниоты
Анамнии, или низшие позвоночные — группа животных, не имеющая зародышевых оболочек (зародышевого органа — аллантоиса и амниона). Анамнии проводят большую часть жизни в воде, без которой невозможно их размножение.
К анамниям относятся рыбы, земноводные.

Амниоты — группа высших позвоночных, характеризующаяся наличием зародышевых оболочек. К амниотам относятся пресмыкающиеся, птицы и млекопитающие.
Зародышевый орган, аллантоис, является органом дыхания и выделения.
За счет особых оболочек, развивающихся в ходе эмбрионального развития, амниона и серозы, у амниот формируется амниотическая полость. В ней находится зародыш, окруженный околоплодными водами. Благодаря такому гениальному устройству, амниотам для размножения и развития более не нужно постоянное нахождение в водоеме, они «обрели независимость» от него.
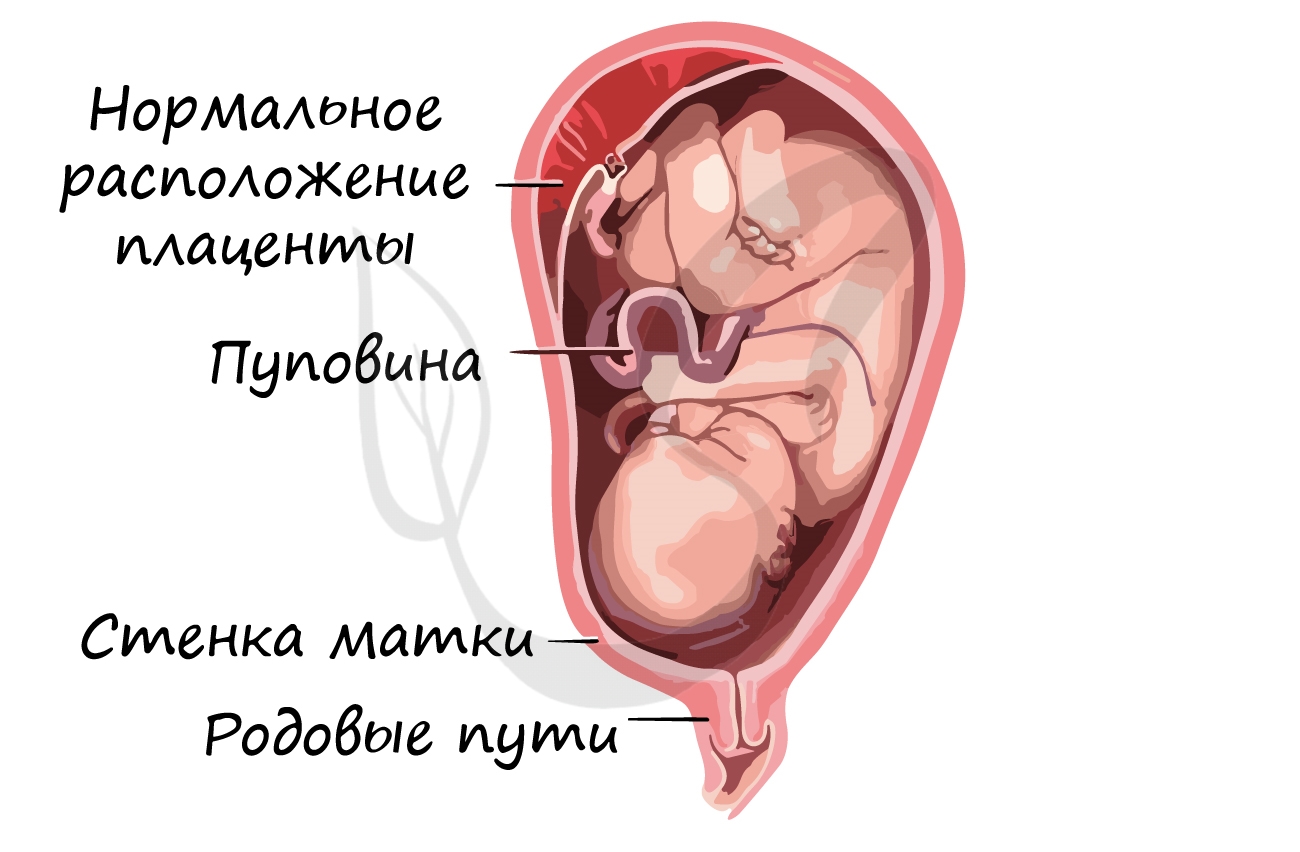
Развитие плода происходит в мышечном органе — матке, которая, сокращаясь во время родов, стимулирует изгнание плода через родовые пути. Питание осуществляется через плаценту — «детское место» — орган, который с одной стороны омывается кровью матери, а с другой — кровью плода. Через плаценту происходит транспорт питательных веществ и газообмен.
Соединяет плаценту и плод особый орган — пуповина, внутри которой проходят артерии, вены.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
МЕЗОДЕ́РМА
В книжной версии
Том 19. Москва, 2011, стр. 582
Скопировать библиографическую ссылку:








МЕЗОДЕ́РМА (от мезо . и греч. δέρμα – кожа) (мезобласт), средний зародышевый листок у всех многоклеточных животных (кроме губок, кишечнополостных) и человека. Располагается между наружным зародышевым листком (эктодермой) и внутренним – энтодермой. У первичноротых животных (большинство беспозвоночных) исходным является телобластич. способ закладки М. из крупных клеток (телобластов), лежащих между эктодермой и энтодермой в первичной полости тела. Телобласты отпочковывают от себя цепочки мелких клеток, формирующих мезодермальные сегменты (сомиты), внутри которых появляется вторичная полость тела – целом. У большинства вторичноротых животных (иглокожих, плеченогих, щетинкочелюстных, бесчерепных, круглоротых, рыб, земноводных) М. образуется энтероцельным способом: из отделяющихся участков стенки первичной кишки (enteron). У др. вторичноротых (пресмыкающихся, птиц и млекопитающих) зачаток М. сначала входит в состав поверхностного слоя зародыша и затем обособляется от него в виде отдельных клеток.
Источник
Гаметогенез и Эмбриогенез 2
1. Дробление. Типы дробления.
2. Бластула. Типы бластул. Влияние факторов внешней среды на процессы дробления.
4. Гистогенез и органогенез.
5. Теория зародышевых листков.
Рекомендуемые файлы
6. Производные зародышевых листков.
ИСПОЛЬЗОВАННАЯ И РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
Антипчук, Ю.П. Гистология с основами эмбриологии / Ю.П. Антипчук. – М.: Просвещение, 1983. – 240 с.
Алмазов, И.В., Сутулов Л.С. Атлас по гистологии и эмбриологии / И.В. Алмазов, Л.С. Сутулов. – М.: Медицина, 1978. – 148 с.
Гистология / под ред. Ю.И. Афанасьева. – М: Медицина, 1989. – 361 с.
Рябов, К.П. Гистология с основами эмбриологии / К.П. Рябов. – Мн.: Высш. шк., 1991. – 289 с.
Биологический энциклопкдический словарь / под ред. М.С. Гилярова. – М.: Сов. Энцикл., 1989. – 864 с.
Практикум по гистологии, цитологии и эмбриологии / под ред. Н.А. Юриной, А.И. Радостиной. – М.: Высш. шк., 1989. – 154 с.
Хэм А., Кормик Д. Гистология / А. Хэм, Д. Кормик. – М.: Мир, 1983. – 192
После оплодотворения яйцеклетка начинает делиться. Про цесс деления яйцеклетки, приводящий к увеличению числа клеток без их роста, называют дроблением. Характер дробления определяется количеством желтка в яйцеклетке и его распределением. В зависимости от этого дробление может быть полным и частичным.. Дробление, при котором яйцеклетка делится полностью, называют голобластическими , а не полностью, частично – меробластическими.
Полное дробление яйцеклетки может быть равномерным и неравномерным.
Полное равномерное дробление свойственно яйцеклеткам ланцетника, яйца которого имеют мало желтка. При этом виде дробления образующиеся клетки, или бласто меры, имеют примерно одинаковые размеры.
Полное неравно мерное дробление свойственно яйцеклеткам хрящевых рыб, земноводных, яйца которых имеют боль ше желтка. При этом виде дробления образуются неодинаковые по величине бластомеры, причем первые два дробления дают равные бластомеры, а затем бластомеры, расположенные на анимальном полюсе, делятся быстрее, становятся мельче, нежели на вегетативном.
Неполное дробление яйцеклеток может быть поверхностным и дискоидальным.
При неполном поверхностном дроблении, ко торое присуще многим членистоногим, дробится лишь поверх ностная часть яйцеклетки, а центральная ее часть, богатая желтком, не делится.
При неполном дискоидальном дробле нии , которое свойственно, например, пресмы кающимся, птицам, дробится лишь небольшой участок поверх ности яйцеклетки, где мало желтка, а остальная ее часть, бо гатая желтком, не делится. В связи с тем что при этом виде дробления поверхностный участок дробящегося яйца имеет форму диска, то и дробление получило наименование дискои дального (рис. 1).
 |
1 – полное равномерное; 2 – полное неравномерное; 3 – неполное поверхностное; 4 – неполное дискондальное.
Характер дробления определяется не только количеством желтка и распределением его в яйцеклетке, но и взаимным расположением клеток, которые образуются в результате дробления.
По направлению дробления различают радиальное, спиральное, двусимметричное, или билатеральное, дробление.
При радиальном дроблении, которое свойственно кишечнополост ным, иглокожим и многим низшим хордовым, верхний ряд бла стомеров располагается точно над нижним.
При спиральном дроблении, которое наблюдается у большинства червей и мол люсков, верхний ряд бластомеров располагается между бластомерами нижнего ряда.
При билатеральном дроблении, которое свойственно круглым червям, асцидиям, бластомеры располага ются симметрично по бокам исходного бластомера. Следует за метить, что в процессе дробления яйцеклетки зачастую имеет место сочетание различных типов дробления.
2. БЛАСТУЛА, ИЛИ ОДНОСЛОЙНЫЙ ЗАРОДЫШ
В процессе дробления возникает многоклеточный однослой ный зародыш, который внешне напоминает шар, во многих слу чаях с полостью в середине. Такой зародыш называют бласту лой . Слой клеток, образующий стенку бластулы, называют бла стодермой ( derma – кожа), а полость бластулы – бластоцелем ( koilon – полость) или первичной полостью тела. Когда же при дроблении возникает шаровидный зародыш без полости внутри, похожий на ягоду шелковицы, то его называют мору лой ( morula ягода шелковицы). Такая бластула свойствен на, например, плацентарным млекопитающим.
В зависимости от типа дробления яйцеклетки различают, кроме морулы, еще 5 типов бластул: целобластулу, амфи бластулу, стерробластулу, перибластулу и дискобластулу (рис. 2).
Бластулу, бластодерма которой образована од ним слоем клеток, а бластоцель занимает центральное положе ние, называют типичной бластулой или целобластулой. Такая бластула свойственна, например, ланцетнику.
Амфибластула отличается от типичной бластулы тем, что ее бластоцель сме щен к анимальному полюсу – зоне мелких бластомеров, а бла стодерма представлена несколькими рядами клеток. Такая бластула свойственна земноводным, от которых и получила свое наименование.
Стерробластула ( sterros – плотный) состоит из крупных клеток бластодермы, которые настолько глубоко за ходят в полость бластоцеля, что от последнего ничего не оста ется. Такую бластулу имеют некоторые членистоногие.
В перибластуле, возникающей при по верхностном дроблении яйца, клетки бластодермы располагают ся по периферии недробящегося желтка и бластоцеля нет. Такой тип бластулы присущ некоторым насекомым.
Диско идальное дробление ведет к формированию дискобластулы, бластоцель которой имеет вид узкой щели, расположенной под клетками бластодермы, именуемой в данном случае зародыше вым диском. Такой тип бластулы наблюдается у костистых рыб, пресмыкающихся, птиц.

Рис. 2. Типы бластул (ориг.):
/ – морула; 2 – цслобластула; 3 – амфнбластула; 4 – дискобластула; 5 – стерробла стула; 6 – перибластула; 7 – плакула.
Рассмотренное многообразие бластул показывает, что на этой стадии развития организмов имеет место дифференцировка клеточного материала зародыша, связанная с количеством и расположением желтка, с содержанием и синтезом ДНК, с раз делением веществ в цитоплазме клеток.
На процессы дробления и ранние стадии развития организ мов влияют условия внешней среды, поскольку развитие организма происходит в среде, в которой проходило его формирование в историческом развитии. Незначитель ные отклонения условий среды от типичных сказываются в ос новном лишь на темпах дробления.
К факторам внешней среды , которые влияют на дробление яйцеклетки и ранние стадии развития организма, следует в первую очередь отнести наличие влаги, химический состав и реакцию среды (рН), световой поток, температуру, наличие питательных веществ, кислорода и др.
Гаструляция – это процесс образова ния двуслойного зародыша.
В более широком представлении гаструляция – сложный процесс химических и морфологических изменений, сопровождающийся размножением, ростом, направленным перемещением и дифференцировкой клеток, в результате чего образуются зародышевые листки, источники зачатков тканей и органов, комплексы осевых органов.
Гаструляция у большинства животных происходит в два этапа. На первом этапе образуются 2 зародышевых листка (эктодерма и энтодерма), а на втором формируется третий 3 зародышевый листок (мезодерма) и происходят дальнейшее формирование эмбриона.
Различают четыре типа, или способа, образования двуслойного зародыша: вселение, или иммиграция; впячивание, или инвагинация; обрастание, или эпиболия; расслоение, или деляминация .
Исходной в процессе филогенетического развития и наибо лее примитивной формой гаструляции является вселение, или иммиграция . Такой способ образования двуслойного зародыша состоит в том, что отдельные клетки или группы клеток активно перемещаются из бластодермы в бластоцель. Эти клетки, возникающие вследствие деления кле ток бластодермы, оседают на ее внутренней поверхности, обра зуя внутренний слой клеток, или энтодерму, а клетки бласто дермы, расположенные снаружи, именуются уже эктодермой. Если вселение клеток в бластоцель имеет место лишь со сто роны одного вегетативного полюса бластулы, то такая имми грация носит название униполярной или однополюсной, а ког да из различных участков бластодермы, – мультиполярной или многополюсной. Униполярная иммиграция свойственна не которым гидроидным полипам, медузам и почти всем гидроме дузам. Мультиполярная иммиграция является редким явлением и наблюдается у некоторых медуз. При иммиграции гастраль ная полость, или гастроцель (полость первичной кишки), обра зуется либо одновременно с возникновением двуслойного за родыша, как это имеет место, например, у гидромедуз, либо значительно позже, как у многих видов кишечнополостных. У последних наблюдается столь массовое вселение клеток, что бластоцель ими полностью заполняется, полость исчезает и лишь впоследствии, с обособлением энтодермы, воз никает гастроцель, естественно, не имеющий связи с внешней средой. Этот способ гаструляции особенно распространен сре ди двуслойных животных – кишечнополостных. Все иные спо собы гаструляции являются производными иммиграции.
Если гаструляция осуществляется путем впячивания, или инвагинации ( in — проникновение, vagina – оболочка, влага лище), то однослойная стенка бластулы – бластодерма – про гибается внутрь бластоцеля и достигает противоположной стен ки. Впячивание начинается с вегетативного полюса, который распознается по более крупным клеткам, и идет в направлении анимального полюса. По окончании инвагинации зародыш ста новится двуслойным, поскольку бластодерма образует два слоя клеток: наружный – эктодерму и внутренний – энтодерму. После впячивания формируется полость, которая окружена клетками энтодермы. Эту полость называют гастральной по лостью. Гастроцель сообщается с внешней средой отверстием, которое называют первичным ртом или бластопором ). Края этого отверстия именуются губами.
В процессе эволюционного развития представителей живот ного царства формирование бластопора при гаструляции по служило основанием для разделения животных на две большие группы по такому признаку, как образование первичного рта. У червей, моллюсков, членистоногих первичный рот, развива ясь и дифференцируясь, превращается в постоянный, или дефи нитивный, рот взрослой особи. Таких животных называют пер вичноротыми. У иглокожих, ки шечнодышащих, хордовых бластопор превращается в задне проходное, или анальное, отверстие, или в нервно-кишечный канал, который находится на заднем конце эмбриона, а рото вое отверстие возникает заново на переднем конце брюшной по верхности зародыша. Таких животных именуют вторичнороты ми . Гаструляция путем впячивания очень распространена у низших вторичноротых и хордовых.
Судьба бластоцеля при инвагинации может быть двоякой: он может сохраниться в том случае, если клеточные слои эктодермы и энтодермы не соприкасаются между собой и тогда у зародыша наблюдаются одновременно две полости – бласто цель и гастроцель. Если же клеточные слои эктодермы и энто дермы при впячивании соприкасаются на всем протяжении, то бластоцель исчезает и у зародыша наблюдается лишь одна полость – полость первичной кишки, или гастроцель.
У некоторых позвоночных животных в связи с увеличением количества желтка в яйцеклетке и незначительными размера ми бластоцеля, оттесняемого к анимальному полюсу, гастру ляция лишь по типу инвагинации становится невозможной. Тог да у некоторых круглоротых и земноводных к впячиванию при соединяется обрастание, или эпиболия . Гаструляция путем эпиболии состоит в том, что клетки бластодермы анимального полюса бластулы делятся значитель но быстрее, чем клетки вегетативного полюса. В связи с этим мелкие клетки анимального полюса постепенно снаружи на ползают на крупные клетки вегетативного полюса, обрастая их и образуя эктодерму, а клетки вегетативного полюса формиру ют энтодерму. При гаструляции путем эпиболии в чистом виде, как это имеет место у некоторых беспозвоночных, вначале нет ни бластопора, ни гастроцеля, которые возникают значительно позже. В тех же случаях, когда эпиболия сопутствует инваги нации, как это наблюдается у некоторых костистых рыб, земно водных, то возникает бластопор, а также полость первичной кишки, которые существуют наряду с бластоцелем.
У костистых рыб, пресмыкающихся, птиц и яйцекладущих млекопитающих, яйцеклетки которых имеют очень много желт ка, в первой фазе гаструляции двуслойный зародыш образуется путем расслоения, или деляминации, клеточного слоя бластодермы. Этот тип образования двуслойного зародыша наблюда ется у кишечнополостных, а также у высших плацентарных млекопитающих, яйцеклетки которых утратили большие запа сы желтка. Гаструляция путем деляминации состоит в делении клеток бластодермы, причем возникающие при этом дочерние клетки оттесняются в бластоцель и образуют внутренний слой клеток, или энтодерму, а наружный слой клеток, или прежняя бластодерма, преобразуется в эктодерму. Когда происходит оттеснение дочерних клеток в бластоцель, мы имеем дело с так называемой первичной деляминацией, а при вторичной делями нации клетки будущей эктодермы, наоборот, оттесняются кна ружи. При этом способе гаструляции бластопор не образуется и гастроцель, естественно, не сообщается с внешней средой.
Следует отметить, что, несмотря на своеобразность отдель ных способов гаструляции, в процессе образования многослой ного зародыша зачастую наблюдается комбинация различных типов гаструляции – двух и даже трех типов.
Образование третьего слоя клеток, или третьего зародыше вого листка – мезодермы, в процессе эмбрионального развития животных осуществляется четырьмя способами: телобластиче ским, энтероцельным, эктодермальным и смешанным, или пере ходным .
У первичноротых животных образование мезодермы осуще ствляется за счет нескольких крупных клеток, располагающих ся во время гаструляции на границе между эктодермой и энтодермой по бокам бластопора. Происхождение этих клеток не выяснено, равно как и отношение их к какому-либо из зароды шевых листков, поскольку они обособляются уже на стадии дробления. Дочерние клетки указанных клеток располагаются между эктодермой и энтодермой и формируют третий слой клеток – мезодерму. Поскольку исходные клетки, образующие мезодерму, в течение всего времени располагаются на заднем конце зародыша, то соответственно своей локализации они по лучили название телобластов, а сам способ образования мезо дермы – телобластического. Вторичная полость тела зароды ша, или целом, образуется клетками мезодермы вследствие их расслоения.
У вторичноротых, включая низших хордовых, мезодерма образуется из клеток энтодермы. При этом способе образова ния мезодермы клетки стенки первичной кишки, или энтодер мы, интенсивно размножаясь, образуют по бокам первичной кишки мешковидные выпячивания в бластоцель. Эти выпячи вания, врастая в бластоцель, располагаются между эктодер мой и энтодермой. В дальнейшем энтодермальные выпячива ния отшнуровываются от энтодермы, а целостность стенки пер вичной кишки и энтодермальных выростов восстанавливается путем размножения клеток. После обособления от первичной кишки клеточный материал энтодермального происхождения именуется третьим зародышевым листком или мезодермой. Раз растаясь, мезодерма заполняет всю полость бластоцеля, а по лость мешкоподобных выростов, являясь по происхождению по лостью гастроцеля, именуется вторичной полостью тела. Этот способ образования мезодермы получил название энтероцельного , поскольку мезодерма происходит из энтодермы при одновременном образовании це лома.
У пресмыкающихся, птиц, млекопитающих животных и че ловека третий зародышевый листок образуется из эктодермы во время второй фазы гаструляции. Во время второй фазы гаструляции наблюдается впячивание клеток экто дермы в бластоцель при одновременной иммиграции клеток эк тодермы в пространство между эктодермой и энтодермой. Клетки, выселяющиеся из эктодермы и размещающиеся между по следней и слоем клеток энтодермы, являются клетками мезо дермы. Этот способ образования мезодермы называется экто дермальным.
У хрящекостных рыб и земноводных наблюдается смешан ный, или переходный, способ образования мезодермы, который является отражением эволюционных преобразований эмбрио нального развития от низших хордовых к высшим. У этих жи вотных мезодерма формируется в процессе гаструляции одно временно с эктодермой и энтодермой.
4. ГИСТОГЕНЕЗ И ОРГАНОГЕНЕЗ
Процесс гаструляции завершается образованием 3 зародышевых листков: наружный – эктодерма; внутренний – энтодерма и средний – мезодерма. В дальнейшем у зародыша происходит формирование нервной трубки, закладка осевых органов (хорда и сомиты), и обособление других эмбриональных зачатков.
Эмбриональные зачатки – источники развития тканей и органов в онтогенезе, представленные группами более или менее многочисленных малодифференцированных (неспециализированных) клеток; межклеточного вещества зачатки не имеют.
К эмбриональным зачаткам относятся:
1) Спинная струна, или хорда ( chorda – хорда);
2) Сегментированная мезодерма, или сомиты;
3) Сегментированные ножки, или нефротом ( nephros – почка);
4) Несегментированная мезодерма (боковые пластинки) или спланхнотом.
Рассмотрим, как происходит формирование нервной трубки, закладка осевых органов и обособление эмбриональных зачатков на примере птиц (рис. 3).

Рис. 3. Закладка осевых органов и эмбриональных зачатков у птиц (ориг.):
1 — эктодерма- 2 — нервная трубка; 3 – сомит; 4 – нефротом; 5 – париетальный ли сток спланхнотома; 6 – целом; 7 – висцеральный листок спланхнотома; 8 – кровяной островок; 9 – энтодерма; 10 – хорда.
Нервная трубка образуется из клеток эктодермы. Процесс образования нервной трубки у зародышей хордовых животных и чело века называется нейруляцией. После образования скопления клеток мезодермы, соот ветствующего будущей хорде, клеточный материал эктодермы, расположенный над хордой, образует медуллярную ( medulla – мозг), или нервную, пластинку. Этот обширный пласт клеток эктодермы, покрывающий хорду и сомиты, слегка прогибается, а по краям его образуются два нервных, или медуллярных, ва лика, представляющие собой края нервной пластинки. Прогибаясь, нервная пластинка образует нервный жело бок, а нервные валики своими гребнями смыкаются, формируя нервную труб ку. У бесчерепных нервная трубка на всем протяжении одина кова, она превращается в спинной мозг. У позвоночных жи вотных и человека передний отдел нервной трубки образуется позднее за счет смыкания более высоких нервных валиков, вследствие чего головной отдел нервной трубки оказывается расширенным и из него впоследствии развиваются мозговые пузыри, а затем головной мозг,
Хорда располагается между энтодер мой и нервной трубкой и имеет вид нерасчлененного сплошного клеточного тяжа. У большинства хордовых животных и челове ка она существует лишь у зародышей. На протяжении всей жизни хорда сохраняется у ланцетника, и некоторых рыб – цельноголовых, осетровых и двоякодышащих.
Скопление клеток мезодермы, располагающихся симметрич но по бокам от хорды, представляет собой сегментированную мезодерму, или сомиты. Этому эмбриональному зачатку свой ственна метамерия, т. е. расчленение спинной части мезодермы на одинаковые части, или сегменты, которые располагаются один за другим вдоль главной оси тела. Такое строение сег ментированной мезодермы является характерным признаком, присущим всем хордовым.
Несегментированная мезодерма, или спланхнотом, расщеп ляется на два слоя клеток, или два листка, – наружный, или пристеночный, или париетальный, который примыкает к эктодерме, и внутренний, или висцеральный , который примыкает к энтодерме. Прост ранство, образующееся между наружным и внутренним лист ками спланхнотома, представляет собой вторичную полость тела. В течение некоторого времени сомиты связаны со спланх нотомом сегментными ножками, или нефротомами, но скоро связь между сегментированной и несегментированной мезодер мой прерывается.
Формирование тканевых зачатков идет на основе процессов детерминации и коммитирования.
Детерминация – генетически запрограммированный путь развития клеток и тканей. В основе ее лежат стойкие изменения репрессии (блокирование) и дерепрессии (деблокирование) генов, определяющих специфику синтеза и-РНК и белков.
Коммитирование – ограничение возможных путей развития клеток.
Одновременно в первичных зачатках зародышевых и внезародышевых органов продолжаются процессы дифференцировки.
Дифференцировка – это изменения в структуре клеток, связанные с их функциональной специализацией, обусловленные активностью определенных генов.
В ходе эмбрионального развития выделяют следующие этапы дифференцировки:
I . Оотипическая дифференцировка – материал будущих зачатков представлен отдельными участками цитоплазмы яйцеклетки или зиготы.
II . Бластомерная дифференцировка – различие в клеточном материале устанавливается в бластомерах.
III . Зачатковая дифференцировка – стадия ранней гаструляции.
IV . Гистогеническая дифференцировка зачатков тканей, когда в пределах одного зародышевого листка появляются зачатки разных тканей.
Эмбриональный гистогенез – процесс возникновения специализированных тканей из малодифференцированного клеточного материала эмбриональных зачатков, происходящий в течение эмбрионального развития органов.
Гистогенез сопровождается размножением и ростом клеток, их перемещением – миграцией, дифференцировкой клеток и их производных, межклеточными и межтканевыми взаимодействиями – корреляциями, отмиранием клеток.
В процессе гистогенетической дифференцировки происходит специализация тканевых зачатков и формирование различных видов тканей. При дифференцировке клеток из исходной стволовой клетки образуются диффероны – последовательные ряды клеток. Количество дифферонов в каждом виде тканей различно.
Результатом гистогенетических процессов является формирование основных групп тканей. Их формирование начинается в эмбриональном периоде и заканчивается после рождения.
В процессе, образования многослойного зародыша последо вательно формируются три полости: бластоцель, гастроцель и целом. После образования гастроцеля и целома в процессе гаструляции и нейруляции бластоцель либо сливается с гаст роцелем, как, например, у земноводных, либо настолько умень шается, что приобретает вид узких щелей, которые преобразу ются в полости кровеносной системы. Гастроцель в процессе развития превращается в полость средней кишки организма, а целом образует вторичную полость тела.
5. ТЕОРИЯ ЗАРОДЫШЕВЫХ ЛИСТКОВ
Первым, кто обратил внимание на возникновение органов из зародышевых листков, или пластов, был К. Ф. Вольф (1759). Изучая развитие цыпленка, он показал, что из «неорганизован ной, бесструктурной» массы яйца возникают зародышевые лист ки, дающие затем начало отдельным органам. К. Ф. Вольф различал нервный и кишечный листки, из которых развиваются соответствующие органы. Впоследствии X . Пандер (1817), по следователь К. Ф. Вольфа, также описал наличие у куриного эмбриона зародышевых листков. К. М. Бэр (1828) обнаружил наличие зародышевых листков и у других животных, в связи с чем распространил понятие о зародышевых листках на всех позвоночных. Так, К. М. Бэр различал первичные зародышевые листки, называя их анимальным и вегетативным, из которых впоследствии, в процессе эмбрионального развития, возникают вторичные зародышевые листки, дающие начало определенным органам.
Описание зародышевых листков значительно облегчило изучение особенностей эмбрионального развития организмов и дало возможность установить филогенетические связи между жи вотными, казалось весьма отдаленными в систематическом отношении. Это было блестяще продемонстрировано А. О. Ковалевским (1865, 1871), который по праву считается основателем современной теории зародышевых листков. А. О. Ковалевский на основании широких сравнительно-эмбриологических сопо ставлений показал, что двуслойную стадию развития проходят почти все многоклеточные организмы. Он доказал сходство зародышевых листков у различных животных не только по происхождению, но и по производным зародышевых листков. Под влиянием учения о зародышевых листках Э. Геккель (1866, 1871) создал теорию гастреи, согласно которой все многокле точные животные происходят от одного общего предка, гипотетического организма гастреи — двуслойного животного, напо минающего зародышей некоторых современных животных на стадии гаструлы либо современных кишечнополостных. Таким образом, зародышевые листки рассматривались как производ ные исходной двуслойной организации предков современных многоклеточных животных. В этом и состоит суть теории за родышевых листков.
Однако в теории зародышевых листков есть ряд исключе нии. Согласно этой теории хорда развивается из энтодермы, нервная система – из эктодермы, а мышечная ткань – из мезо дермы. Однако у пресмыкающихся, птиц, млекопитающих хорда развивается из мезодермы, возникающей из эктодермы. У асци дий определенные группы бластомеров дают одновременно и хорду, и нервную систему, т. е. органы, происходящие по тео рии зародышевых листков из различных зародышевых лист ков. Гладкая мышечная ткань радужной оболочки глаза, мышц волосяных сумок кожи млекопитающих развивается не из мезодермы, как того требует теория зародышевых листков, а из эктодермы. Подобные примеры можно было бы продолжить.
Исключения из теории зародышевых листков объяснимы с точки зрения теории филэмбриогенеза А. Н. Северцова (1939). Их следует рассматривать как результат изменения ранних стадий развития, при которых не только зародышевые листки, но и сам онтогенез способны измениться до неузнаваемости в ходе эволюции под влиянием воздействий внешней среды.
Таким образом, теория зародышевых листков является круп нейшим морфологическим обобщением за всю историю эмбрио логии. Благодаря ей возникло новое направление в эмбриоло гии, а именно эволюционная эмбриология, которая показала, что зародышевые листки, имеющиеся у подавляющего большин ства животных, являются одним из свидетельств общности про исхождения и единства всего животного мира.
6. ПРОИЗВОДНЫЕ ЗАРОДЫШЕВЫХ ЛИСТКОВ
С момента возникновения зародышевых листков их клеточ ный материал специализируется в направлении образования определенных эмбриональных зачатков, а также широкого кру га тканей и органов. Уже на стадии образования зародышевых листков наблюдаются различия в их клеточном составе. Так, клетки эктодермы всегда меньше по размерам, более правиль ной формы и делятся быстрее, нежели клетки энтодермы. Воз никающие в процессе эмбрионального развития в первично однородном материале, а также между клетками зародышевых листков различия называют дифференцировкой. Это заключительный этап эмбриогенеза.
Наружный зародышевый листок, или эктодерма , в процессе развития дает такие эмбриональные зачатки, как нервную труб ку, ганглиозную пластинку, плакоды (локальные утолщения эк тодермы), эктодерму кожи и внезародышевую эктодерму. Из этих эмбриональных зачатков возникают следующие ткани и органы. Нервная трубка дает нейроны и макроглию головного и спинного мозга, хвостовую мускулатуру зародышей амфибий, а также сетчатку глаза. Из ганглиозной пластинки возника ют нейроны и макроглия ганглиев соматической и вегетатив ной нервной системы, макроглия нервов и нервных окончаний, хроматофоры низших позвоночных, птиц и млекопитающих, хромаффинные клетки, мозговой слой надпочечников, скелет ные закладки челюстной, подъязычной, жаберных дуг, хрящей гортани, а также эктомезенхима. Из плакод развиваются ней роны и макроглия некоторых ганглиев, или нервных узлов, го ловы, а также органы равновесия, слуха и хрусталик глаза. Кожная эктодерма дает начало эпидермису кожи и его производным – железам кожи, волосяному покрову, ногтям и пр., эпителию слизистой оболочки преддверия ротовой полости, вла галища, прямой кишки и их железам, а также зубной эмали. Кроме того, из кожной эктодермы развиваются мышечные во локна волосяных сумок кожи и радужная оболочка глаза. Из внезародышевой эктодермы возникает эпителий амниона, хориона и пупочного канатика, а у зародышей пресмыкающих ся и птиц – Эпителий серозной оболочки.
Внутренний зародышевый листок, или энтодерма , в развития образует такие эмбриональные зачатки, как ки шечную и желточную энтодерму. Из этих эмбриональных зачатков развиваются следующие ткани и органы. Кишечная эн тодерма является исходной для образования эпителия желу дочно-кишечного тракта и желез – железистой части печени, поджелудочной железы, слюнных желез, а также эпителия ор ганов дыхания и их желез. Желточная энтодерма дифференци руется в эпителий желточного мешка. Внезародышевая энтодерма развивается в соответствующую оболочку желточного мешка.
Средний зародышевый листок, или мезодерма , в процессе развития дает такие эмбриональные зачатки, как хордальный зачаток, сомиты и их производные в виде дерматома, миотома и склеротома ( scleros – твердый), а также эмбриональную со единительную ткань, или мезенхиму. Кроме того, мезодерма образует нефротом, мезонефрические, или вольфовы, каналы; мюллеровы, или парамезонефрические, каналы; спланхнотом; мезенхиму, выселяющуюся из спланхнотома; внезародышевую мезодерму. Из хордального зачатка у аппендикулярий, бесче репных, круглоротых, цельноголовых, осетровых и двоякодыша щих развивается хорда, которая у перечисленных групп жи вотных сохраняется на всю жизнь, а у позвоночных заменяется скелетогенными тканями. Дерматом дает соединительнотканную основу кожи, миотом – поперечнополосатую мышечную ткань скелетного типа, а склеротом образует скелетные ткани – хря щевую и костную. Нефротомы дают начало эпителию почки, мочевыводящих путей, а вольфовы каналы – эпителию семявы носящих путей. Мюллеровы каналы формируют эпителий яйце вода, матки и первичный эпителиальный покров влагалища. Из спланхнотома развивается целомический эпителий, или ме зотелий, корковый слой надпочечников, мышечная ткань серд ца и фолликулярный эпителий половых желез. Мезенхема, ко торая выселяется из спланхнотома, дифференцируется в клетки крови, соединительную ткань, сосуды, гладкую мышечную ткань полых внутренних органов и сосудов. Внезародышевая мезо дерма дает начало соединительнотканной основе хориона, ам ниона, желточного мешка, а также экзоцеломическому эпи телию.
Образование некоторых эмбриональных зачатков происхо дит раньше формирования зародышевых листков. К ним отно сятся трофобласт и гонобласт. Трофобласт у плацентарных млекопитающих животных и человека образуется еще при дроб лении. Половой зачаток, или гонобласт, также возникает до образования трех зародышевых листков и поэтому не может быть производным какого-либо из них.
1. Назовите и охарактеризуйте основные этапы эмбрионального развития, присущие всем видам животных.
2. Что называется дроблением, его типы. Чем определяется характер дробления?
3. Что называется бластулой? Какие части выделяют в бластуле зародыша? Назовите и охарактеризуйте виды бластул.
4. Какие факторы внешней среды влияют на процессы дробления?
5.Что представляет собой гаструляция и каковы особенности этого этапа эмбрионального развития у животных и человека?
6. Какие этапы выделяют в ходе гаструляции, что образуется на каждом этапе? Что образуется в процессе гаструляции?
7. Перечислите основные типы образования двухслойного и трехслойного зародыша.
8. Что послужило причиной деления животных на первичноротых и вторичноротых?
9. Что называется нейруляцией?
10. Назовите осевые органы, образующиеся в процессе нейруляции.
11. Перечислите эмбриональные зачатки, образующиеся в эмбриогенезе.
12. Назовите основные признаки, присущие всем хордовым.
13. Какие зародышевые листки образуются в процессе эмбриогенеза?
14. Кто является автором теории зародышевых листков?
Люди также интересуются этой лекцией: Лекция 5.
15. Что называется детерминацией, коммитированием и дифференцировкой, охарактеризуйте их.
16. Перечислите основные этапы дифференцировки, выделяющиеся в ходе эмбрионального развития.
Источник



