
Оплодотворение и дробление у ланцетника
Ланцетник – это типичное хордовое животное, у которого в течение жизни сохраняется скелет в виде хорды, недифференцированные нервная и пищеварительная трубки. Это морское животное, его размножение происходит на морских банках. Оплодотворение внешнее. Половые продукты самца и самки выбрасываются в воду к концу дня. Сперматозоид проникает в яйцеклетку после первого деления созревания. Как мы уже знаем, яйцо ланцетника гомолецитального типа, для него характерно полное равномерное дробление с образованием целобластулы.
Первая борозда дробления проходит меридионально, и образуются два бластомера, вторая тоже меридиональная, но проходит перпендикулярно первой, образуя четыре бластомера; третья борозда – экваториальная – образуется восемь бластомеров; далее идет чередование меридиональных и широтных борозд дробления. Такое равномерное дробление происходит до восьмого деления, после стадии 128 бластомеров равномерность дробления нарушается. Это происходит по причине оседания желтка в вегетативном полюсе. Согласно четвертому правилу дробления, борозды дробления в анимальной части будут проходить быстрее, а в вегетативной – медленнее. Образуются бластомеры неодинакового размера, в анимальной части более мелкие клетки, а в вегетативной несколько крупнее.
В результате такого дробления образуется целобластула, которая принимает форму купола. Ее бластодерма состоит из одного ряда клеток, а внутри находится крупная полость – бластоцель, или первичная полость тела. Бластоцель заполнена жидкостью – продуктом выделения самих клеток. Анимальный полюс бластулы называют крышей, а вегетативный – дном. Мелкие клетки крыши бластулы представляют закладку будущей эктодермы, а крупные клетки дна – будущей энтодермы. Между ними располагается закладка будущей мезодермы. Кверху от энтодермы лежит несколько рядов клеток, из которых ближайшие к дну бластулы будут образовывать хорду, а клетки, расположенные выше материала хорды, будут развиваться в клетки нервной пластинки. Все остальные клетки крыши бластулы в последующем дадут начало эпителию кожи.
Эти различия между клетками, которые обнаруживаются на стадии бластулы, намечаются еще до дробления в цитоплазме оплодотворенного яйца, распределяющейся в клетки будущих зачатков.
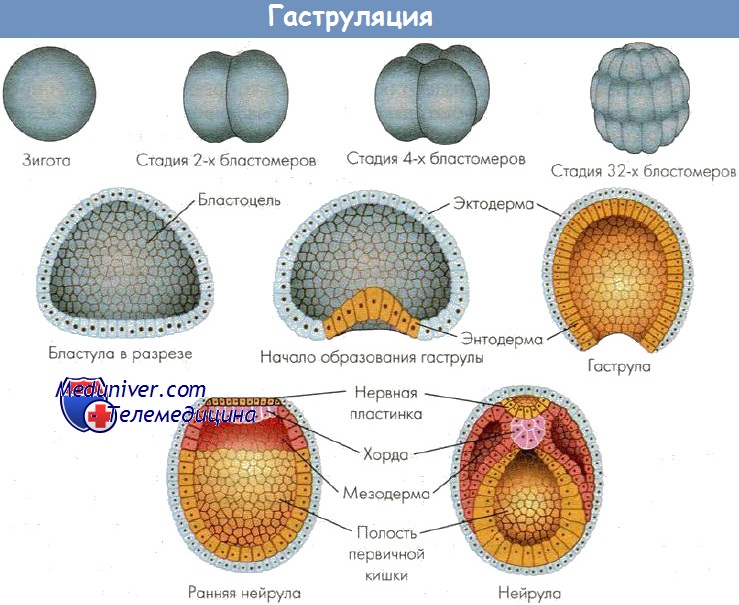
На этом заканчивается цитотипический период в жизни зародыша и начинается органотипический – гаструляция, закладка осевых органов и т.д.
Гаструляция у ланцетника
Гаструляция осуществляется путем впячивания (инвагинации) вегетативной части бластулы в бластоцель. Она начинается в области хордомезодермального участка в виде серповидной бороздки. По мере погружения дна бластулы бороздка увеличивается, закругляется и, наконец, замыкается, ограничивая отверстие бластопора. Клетки дна бластулы постепенно вытесняют бластоцель, и к концу гаструляции бластоцель остается в виде щели в участках, где наружный листок, перегибаясь, переходит во внутренний.
В результате активного перемещения материала все клетки вегетативной части бластулы оказываются внутри, и зародыш становится двухслойным. Бластула превращается в гаструлу. Внутри зародыша образуется новая полость – гастроцель, или полость первичной кишки. Гастроцель сообщается с внешней средой с помощью бластопора, или первичного рта. Бластопор ограничен губами. Со стороны расположения нервной пластинки находится спинная губа бластопора, противоположная ей – брюшная, а по бокам – две боковые губы.
Стенка гаструлы состоит из двух зародышевых листков. Наружный – эктодерма, которая в своем составе содержит группу высоких цилиндрических клеток, представляющих собой медуллярную, или нервную, пластинку. Остальная эктодерма представлена мелкими клетками и является зачатком покрова животного. Стенка первичной кишки образована внутренним зародышевым листком – энтодермой. Энтодерма неоднородна и помимо материала кишечника содержит в своем составе материал хорды и мезодермы. Материал хорды составляет верхнюю часть первичной кишки и располагается под медуллярной пластинкой. Таким образом, внутри зародыша образуется единый зачаток хорды, мезодермы и энтодермы.
В процессе гаструляции материалы будущих внутренних органов, которые в бластуле находились снаружи, перемещаются внутрь зародыша и располагаются на тех местах, где из них будут развиваться соответствующие органы. Только нервная пластинка остается еще на поверхности. Она опустится внутрь зародыша на следующей стадии развития.
В конце гаструляции зародыш поворачивается на 90 0 , и бластопор оказывается сбоку. Зародыш начинает быстро расти и удлиняться.
Источник
Способ дробления у ланцетника
Гаструляция — результат активного деления клеток, роста и направленных перемещений (миграций) клеточных потоков с формированием многослойного зародыша, или гаструлы, (возникновением послойно расположенных, отделенных друг от друга отчетливой щелью, зародышевых листков: наружного — эктодермы, среднего — мезодермы, внутреннего — энтодермы). Перемещение клеток происходит в строго определенной области зародыша — в области серого серпа. Последний был описан В. Ру в 1888 г. В оплодотворенном яйце амфибии серый серп выявляется как окрашенная область на стороне, противоположной проникновению спермия. В этом месте, как полагают, локализуются факторы, необходимые для гаструляции.
У разных представителей позвоночных гаструляция совершается несколькими основными способами: путем инвагинации (впячивания), иммиграции (перемещения части клеток внутрь зародыша), эпиболии (обрастания), деламинации (расщепления). Способы гаструляции зависят от типа яйцеклетки. При любом способе гаструляции в качестве ведущих сил выступают неравномерная пролиферация клеток в разных частях зародыша, уровень обменных процессов в клетках, расположенных в разных частях зародыша, активность амебоидных движений клеток, а также индуктивные факторы (белки, нуклеопротеиды, стероиды и др.). В результате гаструляции обособляются основные зачатки органов и тканей.
Следующим периодом эмбриогенеза является гисто- и органогенез — дифференцировка различных тканей и органов организма из материала зародышевых листков и эмбриональных зачатков.
В результате гаструляции возникает многослойный зародыш. Несмотря на различные способы гаструляции после выделения материала зародышевых листков по оси зародыша находится материал хорды, который подстилает нервную пластинку, слева и справа от хорды располагается материал мезодермы. Все это характеризует осевой комплекс зачатков. В дальнейшем происходит формирование зачатков органов, представляющих собой пространственно локализованные группы стволовых клеток — источников развития тканей. Закономерности дифференцировки клеточного материала зачатков можно проследить в эмбриогенезе наиболее изученных животных.
Ланцетник. Развитие ланцетника.
Классическим объектом эмбриологических исследований ланцентник, подробно изученный А.О. Ковалевским. Ланцетник является представителем класса хордовых животных подтипа бесчерепных, размером до 8 см и обитает на песчаном дне в теплых морях. Свое название получил из-за формы, напоминающей ланцет (хирургический инструмент с обоюдоострым лезвием, современный скальпель).
Яйцеклетка ланцетника олиго- и изолецитальная, размером 110 мкм, ядро располагается ближе к анимальному полюсу. Оплодотворение наружное. Дробление зиготы полное, почти равномерное, синхронное и заканчивается образованием бластулы. В результате чередования меридианальных и широтных борозд дробления формируется однослойная бластула с полостью, заполненной жидкостью — бластоцелем. Бластула сохраняет полярность, ее дно представляет собой вегетативную, а крыша — анимальную части; между ними находится краевая зона.
При гаструляции происходит впячивание вегетативной части бластулы в анимальную. Впячивание постепенно углубляется и, наконец, формируется двустенная чаша с широко зияющим отверстием, ведущим в новообразованную полость зародыша. Такой способ гаструляции называется инвагинацией. Так бластула превращается в гаструлу. В ней материал зародыша оказывается дифференцированным на наружный листок — эктодерму, и внутренний — энтодерму. Полость чаши именуется гастроцелем, или полостью первичной кишки, которая сообщается с внешней средой посредством бластопора, что соответствует заднепроходному отверстию. В бластопоре различают дорсальную, вентральную и две латеральных губы. В результате инвагинации центр тяжести зародыша смещается, и зародыш поворачивается бластопором кверху. Постепенно происходит смыкание краев бластопора и удлинение зародыша. Топография клеток в составе губ бластопора определяет развитие разных частей зародыша. При гаструляции из внутреннего листка гаструлы обособляются хорда и мезодерма, которые располагаются между экто- и энтодермой. Завершается гаструляция образованием осевого комплекса зачатков и дальше — обособлением зачатков органов. Хорда индуцирует развитие нервной трубки из материала дорсальной эктодермы. Эта часть эктодермы утолщается, образуется нервная пластинка (нейроэктодерма), которая прогибается по средней линии и превращается в желобок.
Края желобка постепенно смыкаются в нервную трубку. Оставшаяся часть эктодермы — кожная, срастается над нервной трубкой. Однако на самом переднем и заднем концах зародыша нервная трубка некоторое время сообщается с внешней средой с помощью двух отверстий — нейропоров. В последующем происходит разделение мезодермы на спинные сегменты — сомиты, количество которых нарастает от 15 пар до 60-65 пар у взрослого ланцетника. Часть латерально расположенной мезодермы не сегментируется и расщепляется на наружный (париетальный) и внутренний (висцеральный) листки спланхнотома. Эти листки растут между экто- и энтодермой и, достигнув середины на вентральной стороне зародыша под кишечной трубкой, срастаются, формируя единую вторичную полость — целом. На переднем конце зародыша возникает углубление (ротовая бухта), растущее по направлению к переднему отделу кишечной трубки. При контакте эктодермы ротовой бухты и слепого конца кишечной трубки происходит апоптоз клеток и возникает сообщение кишки с внешней средой. Аналогичный процесс протекает на заднем конце зародыша. По бокам головного отдела зародыша также возникает контакт кожной эктодермы и кишечной энтодермы. В месте этого контакта происходит прорыв. Так полость передней кишки сообщается с внешней средой (формируется жаберный аппарат). После этого зародыш выходит из яйцевой оболочки во внешнюю среду в виде личинки.
Методы маркирования для изучения процессов миграции бластомеров позволили выделить определенные области зародыша на ранних стадиях развития (зиготы — бластулы), которые позднее развиваются в зародышевые листки и эмбриональные зачатки органов и тканей. Эти области были названы презумптивными (предполагаемыми) участками, или зачатками.
Таким образом, на примере развития ланцетника можно наблюдать закономерное перемещение клеток и формирование зародышевых листков, осевого комплекса зачатков и обособление зачатков органов и тканей.
Источник
Дробление у ланцетника
Малое количество желтка объясняет простоту дробления и гаструляции. Дробление полное, почти равномерное, радиального типа, в результате образуется целобластула.

Дробление яйца ланцетника (по Алмазову, Сутулову, 1978):
А – зигота; Б, В, Г – образование бластомеров
(показано расположение веретена деления)
Анимальный полюс приблизительно соответствует будущему переднему концу тела личинки. Оплодотворенная яйцеклетка (зигота) целиком дробится на бластомеры в правильной геометрической прогрессии. Бластомеры почти одинаковой величины, анимальные лишь несколько мельче вегетативных. Первая борозда дробления – меридинальная, проходит через анимальный и вегетативный полюс. Она разделяет шаровидное яйцо на две совершенно симметричные половины, но бластомеры округляются. Они шаровидные, имеют малую площадь соприкосновения. Вторая борозда дробления также меридиональная, пер-
пендикулярна первой, а третья – широтная.
По мере увеличения количества бластомеров они все больше расходятся от центра зародыша, образуя посередине большую полость. В конце концов зародыш принимает форму типичной целобластулы – пузырька со стенкой, образованной одним слоем клеток – бластодермой и с по-
лостью, заполненной жидкостью – бластоцелем.
Клетки бластулы, вначале округлые и потому не плотно сомкнутые, затем приобретают форму призм и плотно смыкаются. Поэтому позднюю бластулу, в противоположность ранней, называют эпителиальной. Стадия поздней бластулы завершает период дробления. К концу этого периода размеры клеток достигают минимума, а общая масса зародыша не увеличивается по сравнению с массой оплодотворенной яйцеклетки.
Дробление у амфибий.
Дробление у амфибий голобластическое (полное), неравномерное и асинхронное. Первая борозда дробления меридиональная, проходит отанимального полюса к вегетативному. Поскольку у амфибий вегетативный полюс перегружен желтком, то чем ближе к нему, тем скорость врезания борозды становится меньшей. Она еще не успевает дойти до вегетативного полюса, как начинает врезаться 2-я борозда.

Рис. 3. Дробление (А – Е) зиготы и поперечный разрез бластулы (Ж) лягушки
(по Гилберту, 1993)
Вторая борозда тоже меридиональная, проходит перпендикулярно первой. После прохождения первой борозды образуется два, после второй – четыре бластомера. На стадии 4-х бластомеров два анимальных бластомера получают по ½ материала серого серпа, а в 2-х вегетативных бластомерах материала серого серпа нет.
Третья борозда дробления широтная, проходит ближе к анимальному полюсу. В результате ее прохождения образуются 8 бластомеров: 4 анимальных и 4 вегетативных, существенно различающихся своими размерами. Далее следуют две меридиональные борозды. Из-за разницы в размерах бластомеров и количества желтка вначале делятся 4 анимальных (кратковременная стадия 12 бластомеров), а затем 4 вегетативных бластомера (стадия 16 бластомеров).
Следующие две одновременно возникающие борозды — широтные. Более быстрое прохождение верхней широтной борозды обусловливает 6 кратковременную стадию 24 бластомеров. Завершение прохождения нижней широтной борозды приводит к образованию 32 бластомеров. После стадии 64 бластомеров несмотря на то, что последовательность борозд сохраняется, геометрическая последовательность ряда (2 — 4 — 8 — 12 — 16 — 24 — 32 — 64) нарушается. Параллельно с прохождением меридиональных и широтных борозд дробления образуется и тангенциальная борозда, в результате чего зародыш приобретает многослойность.
Зародыш амфибий, содержащий от 16 до 64 клеток, обычно называют морулой из-за отдаленного внешнего сходства с тутовой ягодой (лат. morum). На 128-клеточной стадии появляется хорошо различимый бластоцель и принято считать, что в это время зародыш достигает стадии бластулы, хотя образование бластоцеля прослеживается с самого первого деления дробления.( Голиченков В.А.)
Дробление у рыб
 Яйца полилецитальные,
Яйца полилецитальные,
полиспермия. Полагают, что
ядра спермиев, не слившихся с
женским пронуклеусом, долго
сохраняются в желточном слое
яйца, граничащем с бластодермой,
и даже участвуют в переработке
желтка. Дробление дискоидальное.
Пятое деление дробления
отделяет ряд бластомеров от
желтка. Краевые бластомеры
и бластомеры, лежащие в основании бластодермы, сохраняют связь с желтком. Шесть делений дробления проходят более или менее синхронно, после чего синхронность нарушается. В итоге в основании бластодермы формируется перибласт или слой мероцитов. В результате дробления образуется дискобластула, снаружи ограниченная слоем плотно соединенных покровных клеток. Со временем щелевидная полость бластулы увеличивается, причем в большей степени у одного из краев бластодиска. Полагают, что различия в размере бластоцеля (или, иными словами, плотность упаковки клеток в различных частях бластодермы) определяет оси и билатеральную симметрию будущего зародыша — область расширенного бластоцеля соответствует каудальному концу зародыша.
Дробление у пресмыкающихся
Дробление яйцеклетки у пресмыкающихся неполное, дискоидальное. Так как оплодотворение яйцеклетки происходит в верхней трети яйцевода, то дробление начинается во время движения яйцеклетки по яйцеводу и при откладывании яйца зародыш находится либо на стадии дискобластулы, либо ранней гаструлы. У ряда пресмыкающихся, в связи с длительным нахождением яйца в яйцеводе происходит формирование нового организма, который рождается, или выходит из яйца сразу же после его откладывания (живородящая ящерица, гадюки).

Дискоидальное дробление приводит к возникновению дискобластулы. Крыша бластулы состоит из мелких клеток бластодермы, совокупность которых называется бластодиском, а дно бластулы образовано нераздробленной массой желтка. Часть желтка под зародышевым диском резорбируется и образуется подзародышевая полость в виде щели между внешним и внутренним слоем бластодиска.
Дробление у птиц
Дробление и формирование бластулы у птиц проходят при движении яйца вниз по яйцеводу, т.е. одновременно с окружением его третичными оболочками. Период дробления у куриного зародыша составляет 22 часа.
Дробление зиготы у птиц неполное, дискоидального типа. Дроблению подвергается только зародышевый диск, расположенный на анимальном полюсе, имеющий ничтожные поверхность и объем по сравнению с недробящейся массой желтка. Первые три борозды дробления радиальные, соответстветствующие меридиональным бороздам ланцетника и амфибий, затем возникают широтные и тангенциальные борозды (рис. 2).

Рис. 2. Дискоидальное дробление у курицы (по Дондуа, 2005):
А – 2 бластомера; Б – 4 бластомера; В – 8 бластомеров; Г – 16 бластомеров
При дискоидальном дроблении на краю бластодиска и под ним все время остаются неполностью отделенные от желтка клетки (перибласт), от которых обособлены центральные бластомеры. Дробление у птиц резко неравномерное и неправильное. Начиная с прохождения третьей и четвертой борозд образуются клетки разной величины, без какой-либо правильности и постоянства в их расположении.
К моменту откладки яйцо проходит около 14 делений зиготы, в результате бластодерма только что отложенного яйца насчитывает около 60 тыс. клеток. В итоге образуется многослойный бластодиск, состоящий из клеток неправильной формы, вплотную прилежащих друг к другу и налегающих на нераздробившийся желток (рис. 3).

Рис. 3. Формирование зародышевого диска курицы (по Дондуа, 2005):
А – поздняя стадия дробления; Б – перед началом гаструляции:
1 – бластомеры; 2 – эпибласт; 3 – подзародышевая полость; 4 – желток
Под влиянием клеток бластодиска и мероцитов часть желтка под зародышевым диском разжижается, образуется небольшое пространство,
заполненное жидкостью – подзародышевая полость. На этой стадии четко различимы две области бластодиска: светлое поле в центре (area pellucida) и темное по периферии (area opaca). (Голиченков, В. А. Практикум по)
Появление светлого поля связано с тем, что в центре бластодиска
зародышем раньше всего используется желток, вследствие чего образуется подзародышевая щель и вся эта область выглядит прозрачной. Клетки периферической зоны по краю бластодиска налегают на желток, поэтому этот участок зародыша выглядит темным.( Голиченков, В. А)
Дробление умлекопитающих.
Вскоре после образования зиготы начинается ряд митотических делений, строго контролируемых геномом и называемых дроблением; этот процесс начинается спустя несколько часов после оплодотворения, еще в яйцеводах. Каждое деление продолжается от 12 до 24 часов. Зигота делится на клетки-бластомеры в определенной последовательности. Первое деление происходит в плоскости, проходящей через оба полюса яйца, таким образом, шаровидная зигота делится на два полушаровидных бластомера. Возникающая борозда деления располагается не случайным образом, а в зависимости от места проникновения спермия и последующего перераспределения цитоплазмы. Вторая борозда дробления проходит перпендикулярно предыдущей. Получаются четыре бластомера, группирующихся по принципу радиальной симметрии, затем они снова делятся пополам с образованием восьми бластомеров. Продолжающиеся деления бластомеров характеризуются чередованием плоскостей деления – один из двух бластомеров делится в экваториальной плоскости, другой – в меридиональной. Такой тип деления, типичный для млекопитающих, называется чередующимся. Причем бластомеры делятся не одновременно, и в результате этого не происходит четкого нарастания числа клеток от 2 к 4, а далее к 8. В определенные моменты времени зародыши содержат нечетное число бластомеров.
Каждая из клеток, образовавшихся при дроблении, меньше материнской примерно в 2 раза. В период между делениями дробления не происходит интерфазного роста клеток, и поэтому общая масса всех клеток остается примерно равной массе яйцеклетки.

Рис. 1. Ранние стадии эмбрионального развития у млекопитающих:
I – начало дробления;
А – оплодотворения; В, С, D – бластомеры;
II – образование морулы — постепенное обрастание темных клеток светлыми;
III – образование бластоцисты:
А – начало образования полости;
В – обособление зародышевого узелка от трофобласта;
C – превращение зародышевого узелка в зародышевый щиток (дискобластула);
1 – трофобласт; 2 – эмбриобласт.
В результате делений формируется многоклеточный зародыш, напоминающий по внешнему виду ягоду малины и называемый морулой. На стадии 8-клеточной морулы, что характерно только для млекопитающих, происходит значительное сближение 6ластомеров. При этом между клетками возникают плотные контакты, позволяющие некрупным молекулам и ионам переходить из клетки в клетку. Это явление носит название компактизации и способствует дальнейшей дифференцировке зародыша и отделению трофобласта, берущего на себя питательную функцию, и внутренней клеточной массы, дающей начало собственно эмбриону. В процессе компактизации отдельные части плазматической мембраны клеток зародыша начинают двигаться в разных, строго генетически детерминированных направлениях. В этих процессах участвуют белки, входящие в состав клеточных мембран, и с возникновением микроворсинок, прикрепляющих бластомеры друг к другу, изменяется их цитоскелет.
На стадии 32-клеточной морулы зародыш собаки попадает в матку. Это происходит на 7-8-й день, что значительно позже, чем у других млекопитающих. За это время прекращается развитие и происходит гибель зародышей с нарушениями деления, вызванными как наследственными, так и средовыми факторами. Таким образом, наследственные и средовые факторы являются действующими факторами естественного отбора.
Бластомеры, получающиеся в процессе деления, располагаются по периферии, и постепенно в середине плотной клеточной массы морулы формируется полость (бластощель) и отделяется собственно зародышевое скопление клеток – эмбриобласт, иначе называемый зародышевым узлом, и окружающий его слой питающих клеток – трофобласт. Из эмбриобласта позднее формируется тело зародыша. Трофобласт служит питающим листком для раннего зародыша. Эта стадия развития зародыша называется бластоцистой.
Бластоциста на этом этапе состоит из наружного слоя клеток трофобласта и внутренней клеточной массы, шаровидного скопления клеток, прикрепленного изнутри к одному из полюсов трофобласта и представляющего собой материал для построения зародыша. Будущее расположение клетки в зародыше или трофобласте определяется в процессе компактизации, когда клетки оказываются или на поверхности, или внутри зародыша.
Заключение
Список литературы
1. Алмазов И. В. Атлас по гистологии и эмбриологии./ Алмазов И. В., Сутулов Л. С М.: Медицина, 1978.
2. Белоусов Л.В. Основы общей эмбриологии. — Москва: Издательство Московского университета: Наука, 2005.
3. Голиченков, В. А. Практикум по эмбриологии / В. А. Голиченков,
М. Л. Семенова. М. : Academa, 2004.
4. Гилберт С. Биология развития. Т. 1. М.: Мир, 1993.
5. Голиченков, В. А. Эмбриология / В. А. Голиченков, Е. А. Иванов,
Е. Н. Никерясова. М. : Academa, 2004.
6. Дондуа А.К. Биология развития. Начала сравнительной эмбриологии /
С.-Пб.: Издательство СПбГУ, 2005. Т. 1.
7. Карлсон Б. М. Основы эмбриологии по Пэттену. Т. 1. М.: Мир, 1983
8. Кноррэ А. Г. Краткий очерк эмбриологии человека. М.: Медицина, 1967
9. Токин Б.П. Общая эмбриология: Учеб. для биол. спец. ун-тов. — 4-е изд., перераб. и доп. — М.: Высш. шк., 1987. — 480 с.
10. Сотская М.Н. Племенное разведение собак./ Сотская М.Н., Московкина Н.Н. — М.: ООО «Аквариум-Принт», 2006. — С. 95-109.
11. Sze, S. M., Physics of Semiconductor Devices, John Wiley and Sons, N.Y., 1981.
Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Студалл.Орг (0.013 сек.)
Источник



