
II.З. МИТОЗ — ДЕЛЕНИЕ СОМАТИЧЕСКОЙ КЛЕТКИ




![]()
![]()
Наиболее универсальным способом деления соматических клеток, т.е. клеток тела (от греч. soma — тело), является митоз. Этот вид деления клеток был впервые описан немецким гистологом В.Флемингом в 1882г., который наблюдал возникновение и описал поведение нитчатых структур в ядре в период деления. Отсюда происходит и название процесса деления — митоз(от греч. mitos – нить).
При митотическом делении ядро клетки претерпевает ряд строго упорядоченных последовательных изменений с образованием специфических нитчатых структур. В митозе выделяют несколько фаз: профазу, прометафазу, метафазу, анафазу и телофазу (рис. II.2).
Профаза— первая стадия подготовки к делению. В профазе сетчатая структура ядра постепенно превращается в видимые (хромосомные) нити за счет спирализации, укорочения и утолщения хромосом. В этот период можно наблюдать двойную природу хромосом, т.к. каждая хромосома выглядит продольно удвоенной. Эти половинки хромосом (результат редупликации (удвоения) хромосом в 3-фазе), называемые сестринскими хроматинами, удерживаются вместе одним общим участком — центромерой. Начинается расхождение центриолей к полюсам и образование веретена деления (2n4с).
В прометафазепродолжается спирализация хромосомных нитей, происходит исчезновение ядерной оболочки, смешение кариолим-фы и цитоплазмы с образованием миксоплазмы, которая облегчает движение хромосом к экваториальной плоскости клетки (2n4с).
В метафазевсе хромосомы располагаются в зоне экватора клетки, образуя так называемую «метафазную пластинку». На стадии метафазы хромосомы имеют самую малую длину, поскольку в это время они наиболее сильно спирализованы и конденсированы. Эта стадия наиболее пригодна для подсчета числа хромосом в клетке, изучения и описания их строения, определения размеров и т.д. Расположение хромосом по отношению друг к другу является случайным. Веретено деления полностью сформировано, и нити веретена прикрепляются к центромерам хромосом (2n4с).
| Анафазойназывают следующую фазу митоза, когда делятся центромеры хромосом. Нити веретена деления растаскивают сестринские хроматиды, которые с этого момента можно называть дочерними хромосомами, к различным полюсам клетки. Этим обеспечивается согласованное и точное распределение хромосомного материала в дочерние клетки (2n2с). В телофазедочерние хромосомы деспирализуются и постепенно утрачивают видимую индивидуальность. Образуется оболочка ядра, начинается симметричное разделение тела клетки с формированием двух независимых клеток (2n2с), каждая из которых вступает в период О, интерфазы. И цикл повторяется снова. Биологическое значение митоза состоит в следующем. 1. События, происходящие в процессе митоза, приводят к образованию двух ге — | 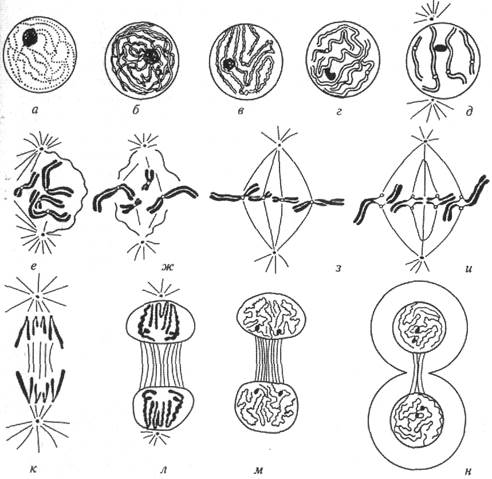 Рис. II.2. Схема митотического деления клетки: а — интерфаза; 6, в, г, д — различные стадии профазы; е, ж — прометафаза; з, и — метафаза; к — анафаза; л, м Рис. II.2. Схема митотического деления клетки: а — интерфаза; 6, в, г, д — различные стадии профазы; е, ж — прометафаза; з, и — метафаза; к — анафаза; л, м телофаза; и — образование двух дочерних клеток |
нетически идентичных дочерних клеток, каждая из которых содержит точные копии генетического материала пред-ковой (материнской) клетки.
2. Митоз обеспечивает рост и развитие организма в эмбриональном и постэмбриональном периоде. Организм взрослого человека состоит примерно из 10 14 клеток, для чего требуется приблизительно 47 циклов клеточного деления единственной оплодотворенной спермием яйцеклетки (зиготы).
3. Митоз является универсальным, эволюционно закрепленным механизмом регенерации, т. е. восстановления утраченных или функционально устаревших клеток организма.
Источник
Способ деления соматической клетки это
Деление соматической клетки и ее ядра (митоз) сопровождается сложными многофазными трансформациями хромосом: 1) в процессе митоза происходит удвоение каждой хромосомы на основе комплементарной репликации молекулы ДНК с образованием двух сестринских нитевидных копий (хроматид), соединенных в области центромеры; 2) в последующем сестринские хроматиды разъединяются и эквивалентно распределяются по ядрам дочерних клеток.
В результате в делящихся соматических клетках поддерживается идентичность хромосомного набора и генеетического материала. Отдельно следует сказать о нейронах — высокодифференцированньгх постмитотических клетках, не претерпевающих клеточных делений на протяжении жизни. Компенсаторные возможности нейронов в ответ на действие повреждающих факторов ограничиваются внутриклеточной регенерацией и репарацией ДНК в неделящемся ядре, чем в значительной степени обусловлена специфика нейропатологических процессов наследственной и ненаследственной природы.
Совершенно иной тип деления — мейоз — характерен для половых клехок. Главной особенностью мейозa являются два последовательных деления клетки-предшественника и ее ядра, в то время как хромосомы удваиваются лишь однажды. Схематично механизм мейоза выглядит следующим образом: 1) в первом делении мейоза дочерние клетки получают из каждой хромосомной пары по одной гомологичной хромосоме, состоящей из удвоенных сестринских хроматид (поскольку при этом число хромосом в дочерних клетках уменьшается вдвое, данное деление является редукционным); во втором делении сестринские хроматиды разъединяются и эквивалентно расходятся по образующимся зрелым половым клеткам — гаметам. В результате число хромосом в гаметах оказывается вдвое меньшим по сравнению с исходной родительской клеткой.
После слияния ядер половых клеток при оплодотворении зигота получает стандартный двойной набор хромосом. Данный: механизм обеспечивает постоянство числа хромосом у разных поколений организмов, размножающихся половым путем.
Важнейшей биологической ролью мейоза является обеспечение генетического разнообразия особей в результате «перемешивания» отцовских и материнских генов в гамете. Это достигается двумя путями. Во-первых, в первом делении мейоза распределение отцовских и материнских хромосом по дочерним клеткам происходит случайным образом, в результате чего гаметы несут различные комбинации родительских хромосом.
Второй фундаментальный механизм поддержания генетического разнообразия заслуживает того, чтобы быть разобранным более подробно, поскольку он имеет прямое отношение к теме настоящей монографии -ДНК-диагностике.
В начальной фазе первого деления мейоза гомологичные хромосомы располагаются друг напротив друга и спариваются, образуя одну или несколько зон контакта (хиазм) между отдельными несестринскими хроматидами. Далее пара хроматид, образовавшая хиазму, обменивается участками ДНК — процесс, носящий, название кроссинговер. В результате кроссинговера образуются рекомбинантные хромосомы, состоящие из участков, имеющих происхождение от разных родительских линий. По завершении мейоза рекомбинантные хромосомы разойдутся по разным гаметам.
Таким образом, кроссинговер представляет собой частный случай генетической рекомбинации — процесса перераспределения генетического материала родителей при передаче потомству. Важным следствием кроссинговера становится создание новой комбинации генов у потомков при соединении родительских гамет. Поскольку при рекомбинации происходит обмен генетического материала между отцовской и материнской хромосомами, этот феномен всегда должен приниматься во внимание при анализе наследования хромосом в процессе нроведения косвенной ДНК-диагностики и расчете генетического сцепления.
Источник


Цитогенетические основы размножения. Репродукция клеток
I. Способы размножения организмов, их сущность
Размножение организмов — это воспроизведение себе подобных. Размножение обеспечивает продолжение жизни, преемственность поколений и сохранение вида. Размножение может быть бесполым и половым.
При бесполом размножении происходит деление соматических клеток одной родительской особи. Наследственный материал передаётся через соматические клетки, и потомки абсолютно тождественны родителям.
При половом размножении новый организм развивается из особой клетки (зиготы), которая образуется при слиянии половых клеток родителей. При образовании половых клеток (гамет) происходит неоднократная перекомбинация наследственного материала (мейоз) и поэтому гаметы генетически неоднородны. При оплодотворении в зиготе комбинируется наследственный материал отца и матери, поэтому потомки не абсолютно тождественны родителям, они имеют индивидуальные особенности.
Особые виды полового размножения: партеногенез и гиногенез. При партеногенезе яйцеклетки способны развиваться в целый организм без оплодотворения (насекомые, плоские черви). При гиногенезе — спермии должны присутствовать в среде для активации яйцеклеток, причём иногда это могут быть спермии другого вида (некоторые рыбы, земноводные и др.).
В развитии одного вида могут чередоваться разные способы размножения. Метагенез — чередование полового и бесполого размножения (кишечнополостные, простейшие). Гетерогенез — чередование полового размножения с оплодотворением и партеногенеза (сосальщики).
Формы полового и бесполого размножения разнообразны у одноклеточных и многоклеточных организмов.
Особый вид бесполого размножения, который встречается и у человека — полиэмбриония. Обычно из зиготы развивается один организм, но иногда, после одного или нескольких (реже) митотических делений, образовавшиеся клетки (бластомеры) могут развиваться в самостоятельные организмы. Так развиваются однояйцевые близнецы — генетически идентичные организмы.
В основе любого размножения, роста и развития организма лежат процессы репродукции клеток (пролиферация клеток). Процессы репродукции, скорость пролиферации, частота делений клеток в разных органах находятся под регулирующим контролем генетического аппарата. Если этот контроль нарушается, то начинается нерегулируемое размножение клеток — развиваются новообразования (опухоли). В течение жизни организма клетки стареют и погибают, поэтому всегда происходят процессы самообновления, образование новых клеток (физиологическая регенерация). В молодом организме репродукция клеток преобладает, за счёт чего увеличивается количество клеток, увеличиваются размеры органов и организм растёт. У взрослых — репродукция клеток обеспечивает только замещение погибших, а не увеличение их количества.
II. Клеточные популяции
Жизненный цикл клетки (ЖЦ) или клеточный цикл — период с момента возникновения клетки в результате деления материнской до ее гибели или исчезновения в процессе собственного деления.
Митотический цикл (МЦ) — период подготовки клетки к делению и само деление.
При подготовке к делению клетка проходит особые периоды интерфазы: пресинтетический (G1), синтетический (S) и постсинтетический (G2).
Таким образом, митотический цикл (МЦ) = G1 + S + G2 + митоз. Часть интерфазы, не имеющая отношения к МЦ, характеризует активное функциональное состояние (состояние покоя у камбиальных клеток) и обозначается — G0.
Жизненный цикл клеток разных популяций неодинаков (рис 13).

По продолжительности жизни и по отношению к делению различают три популяции клеток: стабильная, растущая и обновляющаяся.
Стабильная популяция. Клетки этой популяции имеют наибольшую продолжительность жизни, высокодифференцированы и не способны делиться. К такой популяции относятся нервные клетки, клетки миокарда.
Растущая популяция. Клетки этой популяции высокодиф-ференцированные, имеют большую продолжительность жизни. Они составляют основную массу клеток внутренних органов (печень, поджелудочная железа, и т. д.). Эти клетки не утратили способности к делению, они могут перестраивать свой метаболизм, снижать уровень дифференцировки и делиться.
Обновляющаяся популяция. Среди клеток этой популяции есть два типа: высокодифференцированные и недифференцированные (стволовые или камбиальные клетки).Высокодифференцировнные клетки живут недолго (часы, сутки, месяцы), неспособны к делению,
постоянно отмирают. Например, клетки поверхностного слоя эпидермиса, клетки крови, клетки слизистой оболочки кишечника.
Недифференцированные (стволовые) клетки этой популяции постоянно делятся, дифференцируются и замещают погибшие. Так, стволовые клетки эпидермиса находятся в самом нижнем (мальпигиевом) слое; стволовые клетки слизистой оболочки кишечника — в глубоких отделах кишечных крипт, стволовые клетки крови — в красном костном мозге.
Таким образом, ЖЦ клеток стабильной популяции и дифференцированных клеток обновляющейся популяции равен G0, в их жизненном цикле нет митотического цикла.
ЖЦ стволовых клеток обновляющейся популяции состоит из подготовки к делению и деления, т.е. равен МЦ (периодом G0 в данном случае можно пренебречь, т.к. клетки функционально не активны, находятся в состоянии покоя). Такой ЖЦ также имеют клетки злокачественных опухолей, т. к. они не дифференцируются в нормальные клетки, а снова и снова вступают в деление.
ЖЦ клеток растущей популяции состоит из G0+(G1+S+G2+M)
III. Регуляция временной организации клетки
В разные периоды жизненного цикла строение клетки и ее функции различны.
В интерфазе в клетке идут разнообразные метаболические процессы, происходит реализация генетической информации, клетка выполняет соответствующие функции, то есть она функционально активна (G0). При переходе клетки в МЦ перестраивается характер метаболических процессов; синтезируются новые белки (структурные и ферменты), которые будут обеспечивать нормальный процесс деления. В этот период клетка утрачивает признаки специализации (дифференцировки) и те функции, которые свойственны клеткам данного типа.
Изучены некоторые факторы и механизмы, которые изменяют временную организацию клетки и либо способствуют делению, либо тормозят деление:
1.Ядерно-плазматическое отношение, сфера влияния ядра (отношение объема ядра к объему цитоплазмы). Если это отношение уменьшается, то клетка начинает подготовку к делению.
2.Повышение концентрации в клетке предшественников ДНК (в особенности тимидина) способствует переходу клетки к МЦ. 3.При подавлении синтеза белка и РНК процессы подготовки к делению тормозятся.
4.Наличие в клетке белков-кейлонов тормозит переход клетки из одного периода в другой на любой стадии.
5. Некоторые вещества провоцируют деление клеток, которые в нормальных условиях не делятся. Например, если лейкоциты периферической крови (высокодифференцированные клетки обновляющейся популяции) поместить в среду с фитогемагглютинином (ФГА), то они начинают делиться. Это используют для изучения кариотипа.
6. Выявлен ген клеточного деления (ген CDC), который кодирует белок протеинкиназу, способствующий вступлению клетки в МЦ.
IV. Понятие о кариотипе
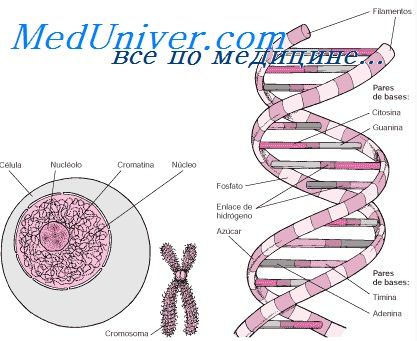
Генетический аппарат эукариотической клетки представлен хромосомами. Кариотип — это совокупность хромосом соматической клетки,
признак постоянный и специфичный для вида. Кариотипы разных видов различаются количеством, величиной и строением хромосом.
Количество (набор) хромосом принято обозначать как » n «. Для разных видов организмов коэффициент » n » — различный.
Каждая соматическая клетка содержит двойной (парный) набор хромосом » 2n «. Такой набор называется диплоидным. Одинаковые хромосомы называются гомологичными (парными). Половые клетки содержат одинарный набор хромосом » n «. Такой набор называется гаплоидным. У человека соматические клетки содержат 46 хромосом (23х2), из них 44 хромосомы (22х2) — аутосомы, одинаковые для мужского и женского пола и 2 хромосомы половые (гетеросомы): у женщин — ХХ, у мужчин — ХУ. По химическому строению хромосомы представляют собой дезоксирибонуклеопротеид (ДНП). В состав одной хромосомы входит одна молекула ДНК. В различные периоды жизненного цикла клетки хромосомы имеют разную морфофункциональную организацию. Хромосомы в виде плотных структур выявляются во время деления клетки, а в интерфазе они деспирализованы и вся совокупность наследственного материала носит название — хроматин. Основная биологическая роль интерфазных хромосом — передача генетической информации.
Основная биологическая роль хромосом при делении — равномерное распределение их между дочерними клетками.
Большая часть интерфазного хроматина находится в деконденсированном состоянии, на разных его участках идут процессы транскрипции — это активный хроматин — эухроматин. Но есть участки конденсированного (плотного) хроматина — это неактивный гетерохроматин.
Различают структурный (конститутивный) гетерохроматин — участки постоянно конденсированные, неинформативные. Второй вид гетерохроматина — факультативный, это те участки, которые могут деконденсироваться и переходить в активное состояние. При подготовке к делению происходит постепенная общая конденсация хроматина и весь генетический материал представляет собой факультативный гетерохроматин; в световом микроскопе он обнаруживается в виде плотных структур — хромосом.
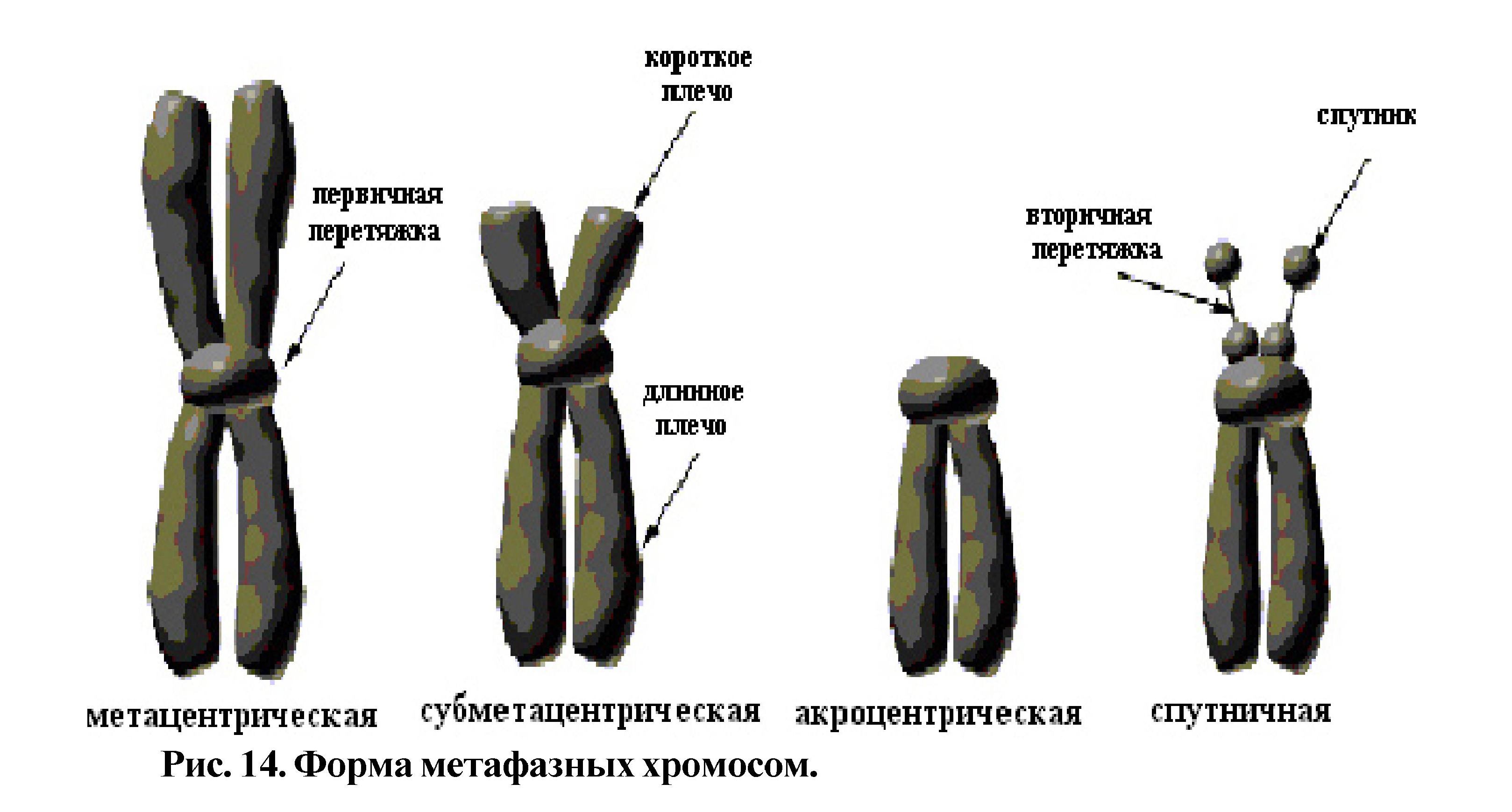
Процесс конденсации хроматина очень важен для регуляции активности генетического материала и для свободного распределения хромосом в цитоплазме клетки во время деления. По мере конденсации активность хроматина снижается. В результате усиления компактизации хроматина и процесса спирализации метафазные хромосомы уменьшаются по длине в несколько тысяч раз и свободно размещаются в цитоплазме клетки, а затем расходятся к полюсам. Форма метафазных хромосом зависит от расположения центромеры.
Различают метацентрические, субметацентрические, акроцентрические хромосомы и хромосомы с вторичной перетяжкой (спутничные) (рис. 14).
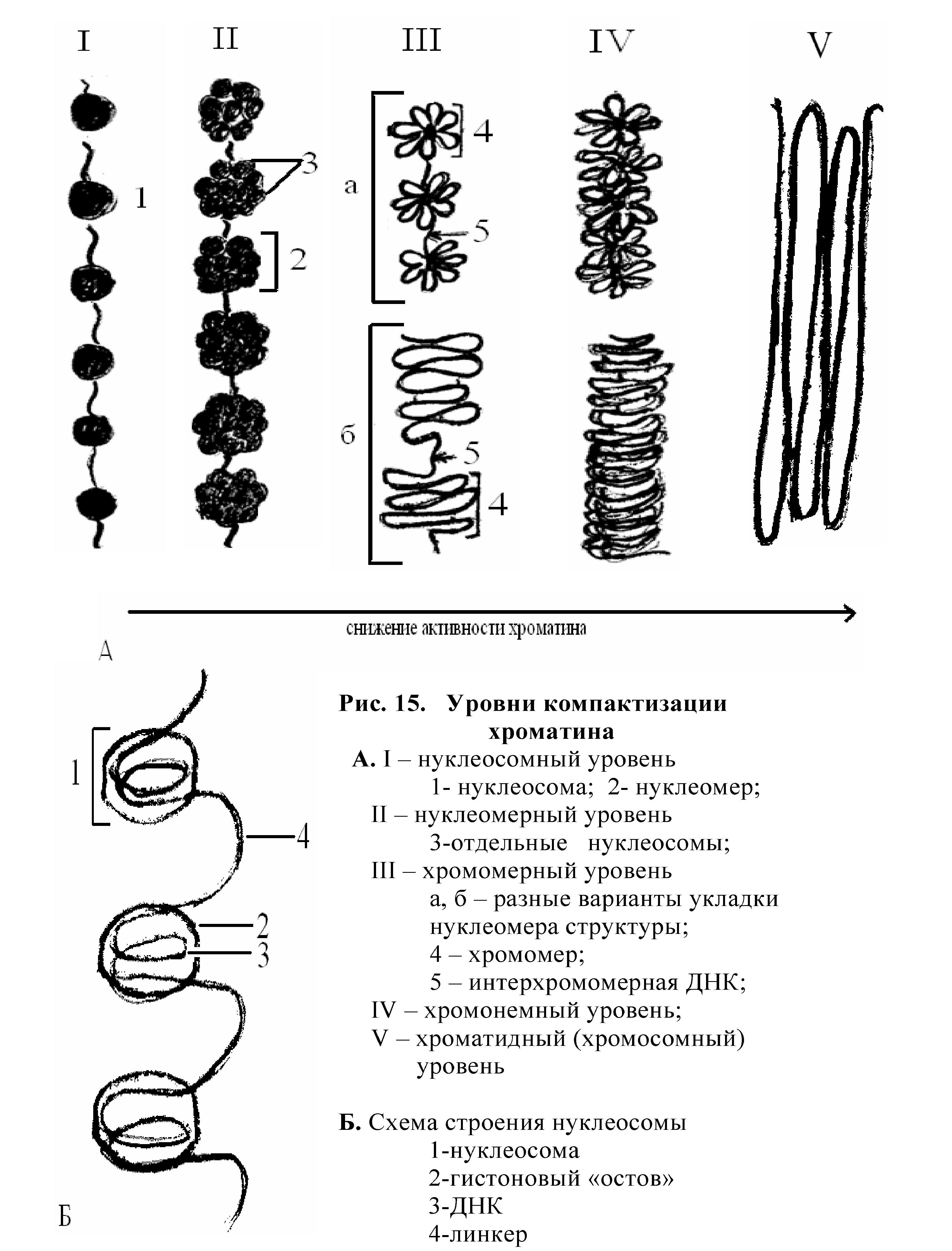
Различают несколько уровней организации хроматина (рис.15):
1. Расправленные нити. Эта структура состоит из 1 молекулы ДНК и молекул гистонов, расположенных параллельно. Неактивный хроматин.
2. Нуклеосомный уровень. Формируются компактные структуры из 8 молекул гистонов и участка молекулы ДНК (около 200 пар нуклеотидов) — нуклеосомы. Хроматиновая нить укорачивается в 7 раз. Наиболее активный хроматин.
3. Нуклеомерный. Объединяются 8-10 нуклеосом, образуется нуклеомер. Укорочение нити в 20 раз.
4. Хромомерный. Нуклеомерная нить образует петли, соединённые белками. Укорочение в 200 раз.
5. Хромонемный уровень образуется в результате сближения хромомеров по длине.
6. Хроматидный. Хромонема складывается в несколько раз, образуя тело хроматиды. Хроматиду можно назвать нереплицированной хромосомой. После репликации ДНК хромосома содержит 2 хроматиды — это реплицированная хромосома.
V. Способы репродукции клеток
Различают несколько способов деления клеток: митоз, амитоз, мейоз.
Митоз — универсальный способ деления клеток. Это непрямое сложное деление, характерное для соматических клеток. Биологическое значение митоза — увеличение количества генетически идентичных клеток.
Амитоз — это простое, прямое деление ядра на две или более частей. Не формируется аппарат деления, способствующий строго равномерному распределению генетического материала между дочерними ядрами. Дочерние ядра могут содержать разный объём генетического материала. Таким образом, амитоз нельзя считать полноценным делением. Деление цитоплазмы часто не происходит, и тогда образуются двуядерные (многоядерные) клетки. Такие клетки теряют способность в дальнейшем вступать в полноценное митотическое деление. Различают три вида амитоза: реактивный, дегенеративный и генеративный.
Мейоз — сложное деление, в результате которого образуются половые клетки (гаметы). Состоит из двух последовательных делений. Особенно сложным является первое деление мейоза (профаза I). При мейозе происходит перекомбинация генетического материала (кроссинговер, независимое расхождение целых хромосом в анафазе I и независимое расхождение хроматид в анафазе II). В результате мейоза образуются гаплоидные клетки («nc») и возникает комбинативная изменчивость. Биологическое значение мейоза состоит в поддержании постоянства кариотипа и возникновении генетически неидентичных гамет, что определяет формирование организмов с индивидуальными особенностями. Мейоз происходит в процессе гаметогенеза (образование половых клеток) в половых железах (гонадах).
VI. Эндорепродукция
Эндорепродукция — явление, связанное не с увеличением количества клеток, а с увеличением (репродукцией) генетического материала в клетке. Различают два вида эндорепродукции: эндомитоз и политению.
Эндомитоз происходит при нарушении нормального хода митоза (сохранение ядерной оболочки в профазе, разрушение митотического аппарата в начале анафазы) и приводит к увеличению плоидности клетки, кратное » n «. Если в эндомитоз вступила клетка, содержавшая 2n, то образуется клетка — 4 n и т. д. Таким образом, результат эндомитоза — полиплоидия.
Политения — образование гигантских политенных (многонитчатых) хромосом. В периоде S одна репликация ДНК следует за другой десятки и сотни раз, поэтому образуются хромосомы, содержащие сотни молекул ДНК. Важным является то, что это — интерфазные хромосомы, на которых идут транскрипционные процессы (области пуффов), и это можно наблюдать в световой микроскоп. Сравнивая локализацию пуффов и синтез определённых белков, можно составить цитологические карты хромосом, то есть ориентировочно определить локализацию отдельных генов на хромосоме (рис.16). Биологическое значение политении — увеличение числа идентичных генов, и как следствие, резкая интенсификация синтеза определённых белков.
Источник



