
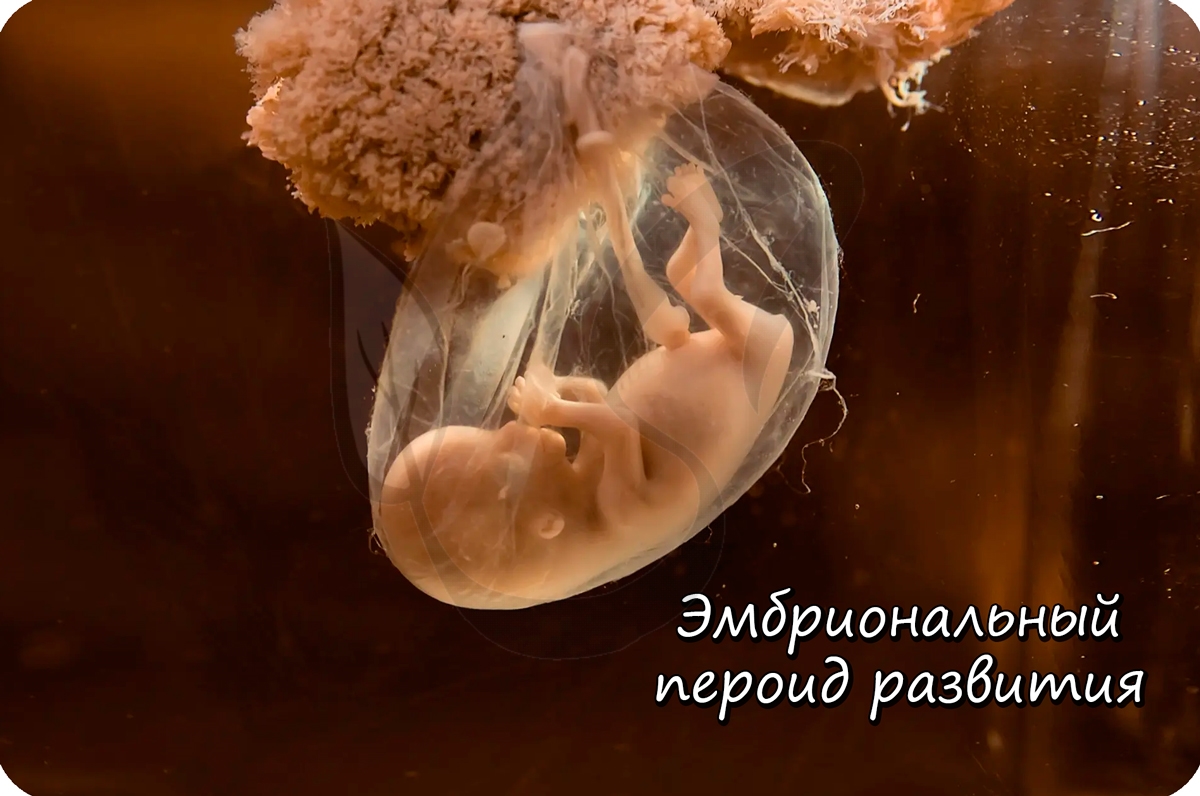
Эмбриональное развитие
От момента образования зиготы и до выхода зародыша из яйцевых оболочек длится эмбриональный период развития.
Дробление зиготы
После того, как произошло оплодотворение — слияние сперматозоида и яйцеклетки, образовавшаяся зигота начинает интенсивно делиться. Ее множественные митотические деления называют дроблением.
Важная особенность дробления в том, что не происходит увеличение в размере зародыша: клетки дробятся (делятся) настолько быстро, что не успевают накопить цитоплазматическую массу. Дробление зиготы человека является полным неравномерным асинхронным.
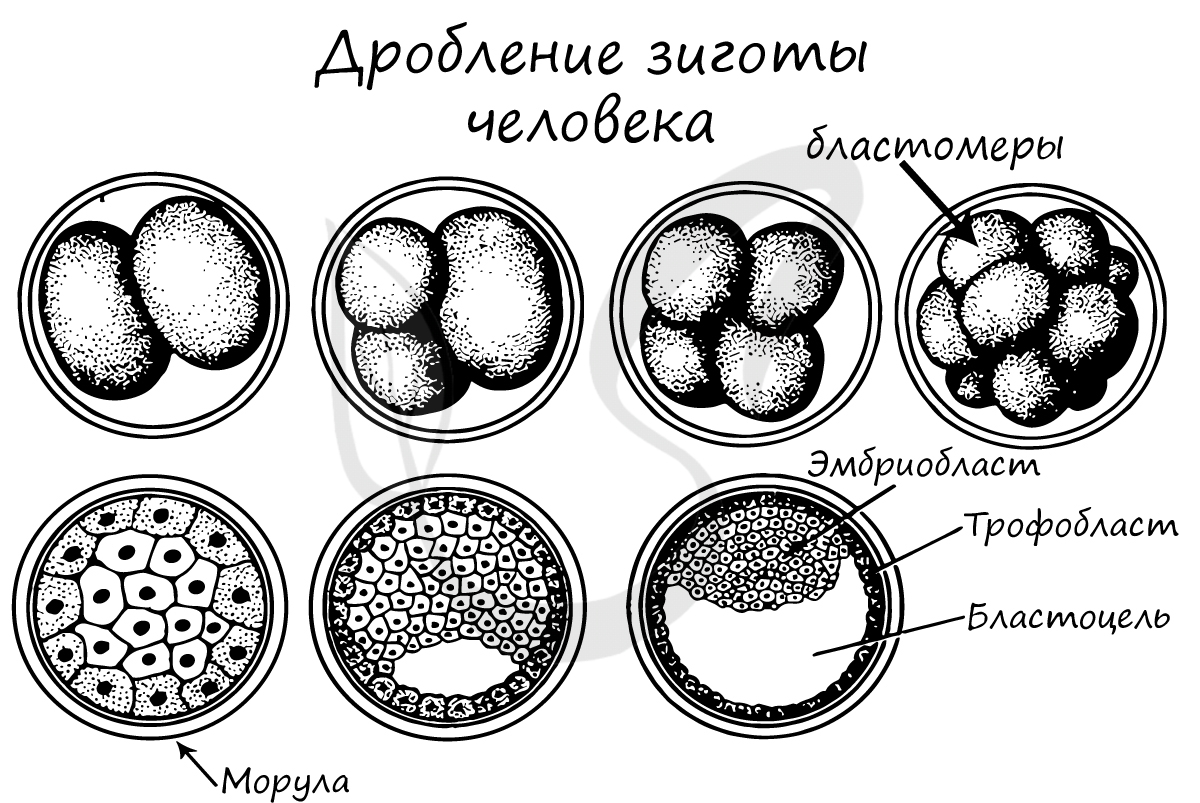
В результате дробления образуется морула. Морула (лат. morum — ягода тутового дерева) — клетка на стадии этапа дробления, когда зародыш представляет собой компактную совокупность клеток (без полости внутри).
Бластуляция
Бластуляция — заключительный период дробления, в который зародыш называется бластулой.
После очередных этапов многократного деления образуется однослойный зародыш с полостью внутри — бластула (греч. blastos — зачаток).
Стенки бластулы состоят из бластомеров, которые окружают центральную полость — бластоцель (греч. koilos — полый). Соединяясь друг с другом, бластомеры образуют бластодерму из одного слоя клеток.

Гаструляция (греч. gaster — желудок, чрево)
Гаструляцией называют стадию эмбрионального развития, в ходе которой клетки, возникшие в результате дробления зиготы, формируют три зародышевых листка: эктодерму, мезодерму и энтодерму.
Стенка бластулы начинается впячиваться внутрь — происходит инвагинация стенки. По итогу такого впячивания зародыш становится двухслойным. Двухслойный зародыш называется — гаструла. Полость гаструлы называется гастроцель (полость первичной кишки), а отверстие, соединяющее гастроцель и внешнюю среду — первичный рот (бластопор).

У первичноротых животных на месте первичного рта (бластопора) образуется ротовое отверстие. К первичноротым относятся: кишечнополостные, плоские, круглые и кольчатые черви, моллюски, членистоногие.
У вторичноротых на месте бластопора формируется анальное отверстие, а ротовое отверстие образуется на противоположном полюсе. К вторичноротым относят хордовых и иглокожих (морских звезд, морских ежей).

При впячивании части бластулы (инвагинации) клетки бластодермы мигрируют внутрь и становятся энтодермой (греч. entós — внутренний). Оставшаяся часть бластодермы снаружи называется эктодермой (греч. ἔκτος — наружный).
Между энто- и эктодермой из группы клеток формируется третий зародышевый листок — мезодерма (греч. μέσος — средний).

Нейрула
Эта стадия следует за гаструлой. Ранняя нейрула представляет собой трехслойный зародыш, состоящий из энто-, экто- и мезодермы. На этапе нейрулы происходит закладка отдельных органов.
Важно отметить, что на стадии нейрулы происходит процесс нейруляции — закладывание нервной трубки. Нервная пластинка, образовавшаяся на ранних этапах, прогибается внутрь, при этом ее края сближаются и, замыкаясь, формируют нервную трубку.

Итак, как уже было сказано, на стадии нейрулы закладываются отдельные органы. Эктодерма образует покровный эпителий и нервную пластинку, мезодерма (из которой в дальнейшем появятся все соединительные ткани), энтодерма — окружает полость первичной кишки (гастроцель), образуя кишечник. От энтодермы отшнуровывается хорда.
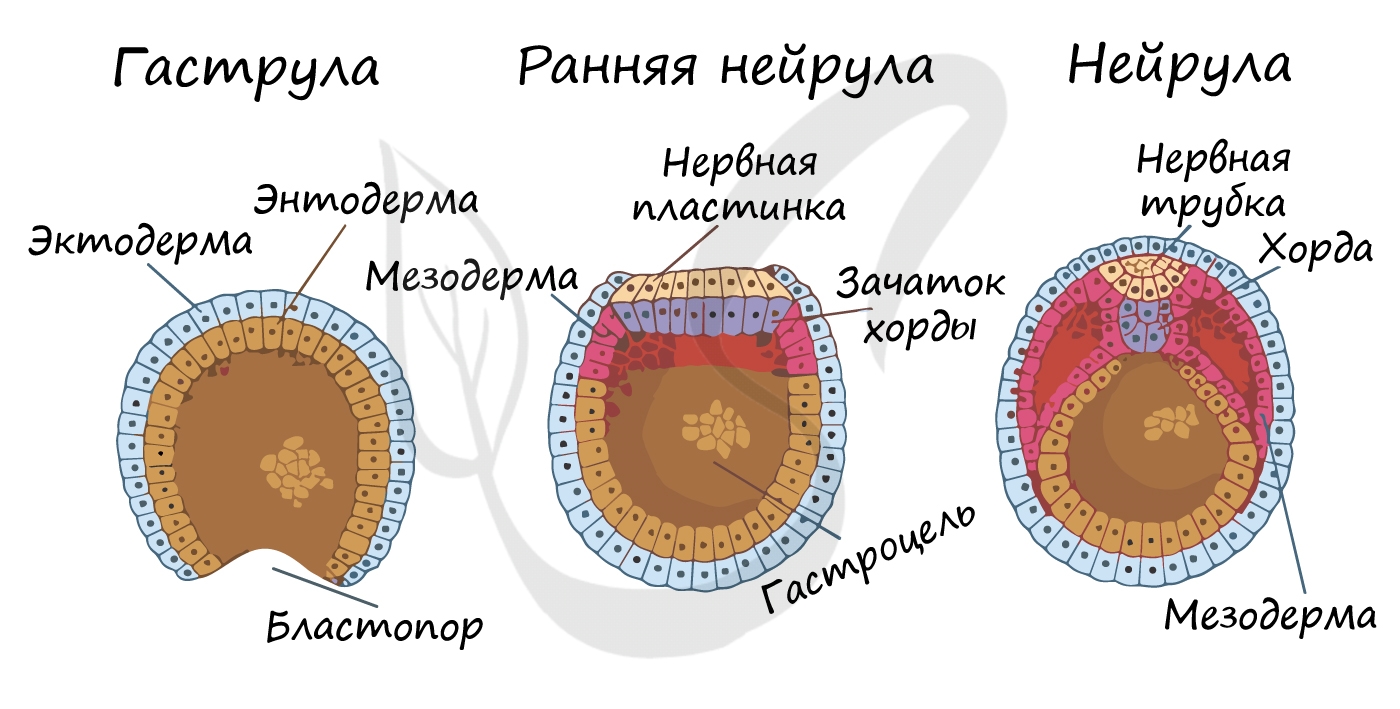
Все три зародышевых листка требуют нашего особого внимания, а также понимания того, какие органы и структуры из них образуются.
Эктодерма (греч. ἔκτος — наружный) — наружный зародышевый листок, образует головной и спинной мозг, органы чувств, периферические нервы, эпителий кожи, эмаль зубов, эпителий ротовой полости, эпителий промежуточного и анального отделов прямой кишки, гипофиз, гипоталамус.
Мезодерма (греч. μέσος — средний) — средний зародышевый листок, образует соединительные ткани: кровеносную и лимфатическую системы, костную и хрящевую ткань, мышечные ткани, дентин и цемент зубов, а также выделительную (почки) и половую системы (семенники, яичники).
Энтодерма (греч. entós — «внутренний») — внутренний зародышевый листок, образует эпителий пищевода, желудка, кишечника, трахеи, бронхов, легких, желчного пузыря, мочевого пузыря и мочеиспускательного канала, печень и поджелудочную железу, щитовидную и паращитовидную железы.

Из зародышевых листков образуются ткани, органы и системы органов. Такой процесс называется органогенезом. В период закладки органов важное значение имеет воздержание матери от вредных привычек (алкоголь, курение), которые могут нарушить процесс дифференцировки клеток и привести к тяжелейшим аномалиям, уродствам плода.
Некоторые лекарства также могут оказывать на плод тератогенный эффект (греч. τέρας — чудовище, урод), приводя к развитию уродств. Периоды закладки органов и система органов вследствие их большой важности носят название критических периодов эмбриогенеза.
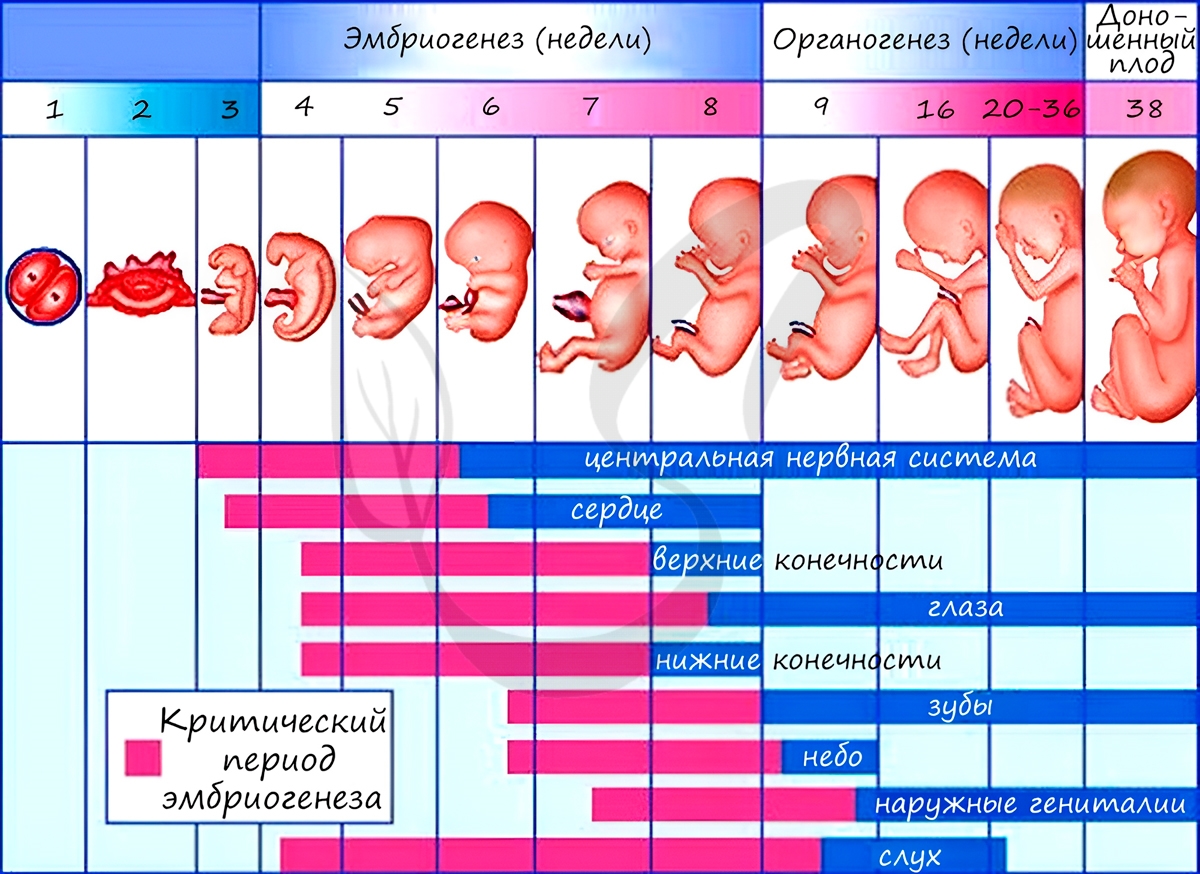
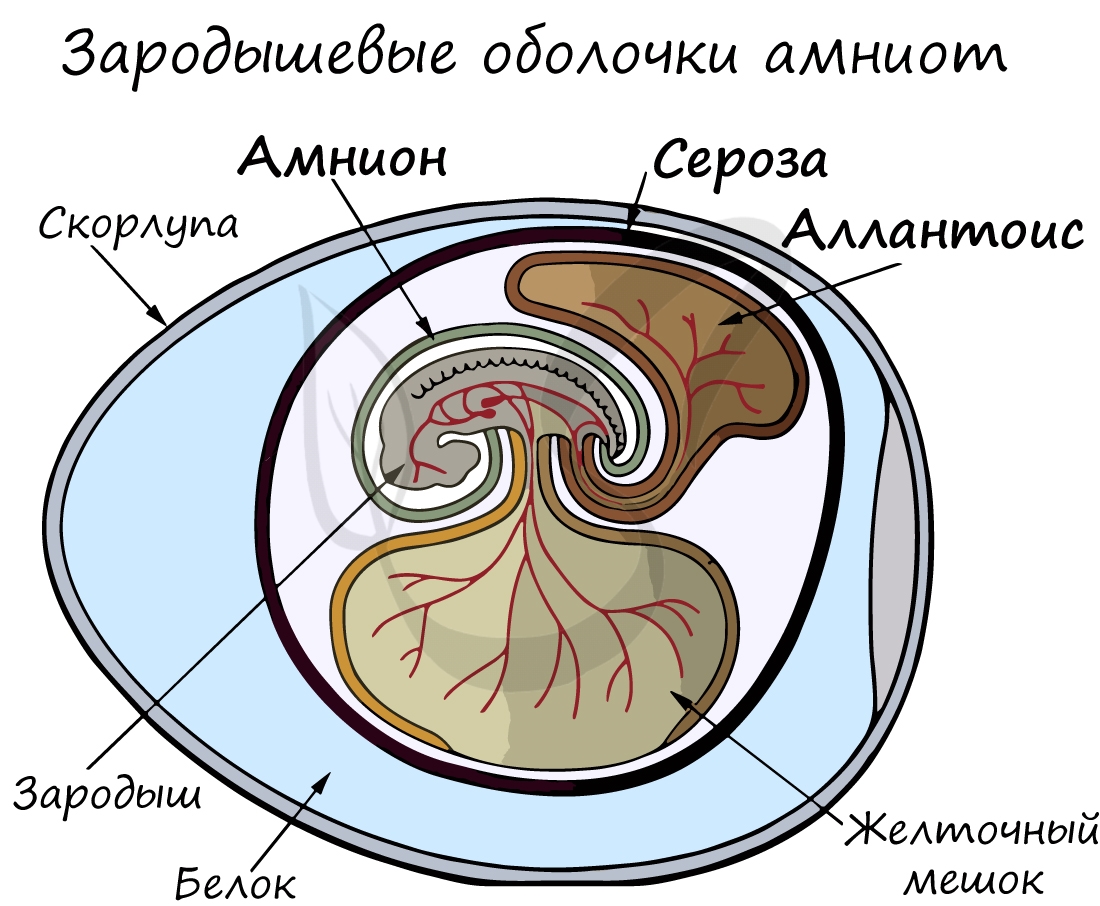
Анамнии и амниоты
Анамнии, или низшие позвоночные — группа животных, не имеющая зародышевых оболочек (зародышевого органа — аллантоиса и амниона). Анамнии проводят большую часть жизни в воде, без которой невозможно их размножение.
К анамниям относятся рыбы, земноводные.

Амниоты — группа высших позвоночных, характеризующаяся наличием зародышевых оболочек. К амниотам относятся пресмыкающиеся, птицы и млекопитающие.
Зародышевый орган, аллантоис, является органом дыхания и выделения.
За счет особых оболочек, развивающихся в ходе эмбрионального развития, амниона и серозы, у амниот формируется амниотическая полость. В ней находится зародыш, окруженный околоплодными водами. Благодаря такому гениальному устройству, амниотам для размножения и развития более не нужно постоянное нахождение в водоеме, они «обрели независимость» от него.
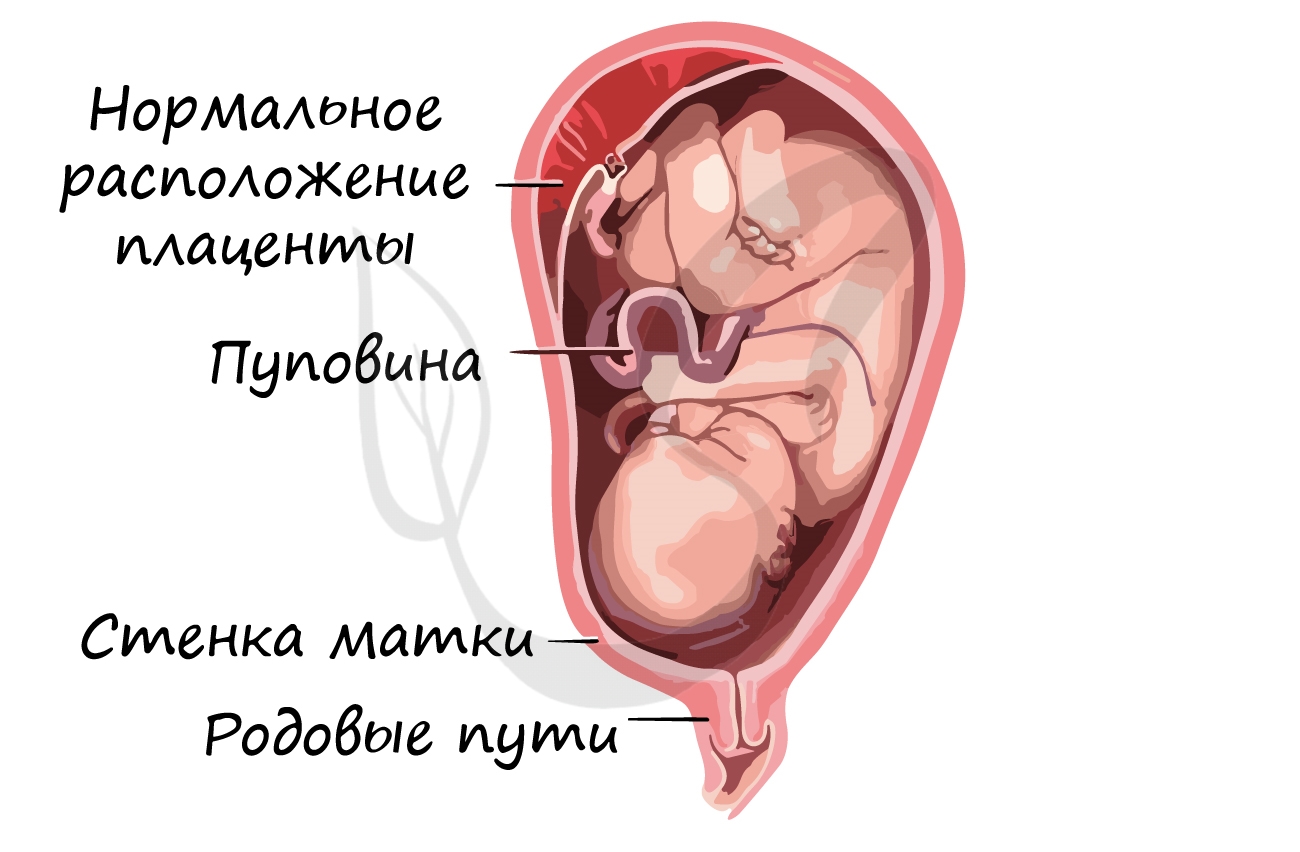
Развитие плода происходит в мышечном органе — матке, которая, сокращаясь во время родов, стимулирует изгнание плода через родовые пути. Питание осуществляется через плаценту — «детское место» — орган, который с одной стороны омывается кровью матери, а с другой — кровью плода. Через плаценту происходит транспорт питательных веществ и газообмен.
Соединяет плаценту и плод особый орган — пуповина, внутри которой проходят артерии, вены.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Какой способ деления клеток наблюдается при дроблении зиготы?
Какой способ деления клеток характерен для дробления зиготы:
А) репликация;
Б) мейоз;
В) амитоз;
Г) митоз.
![]()
Деление клеток нужно для развития организмов.
Существует как прямое деление ядра и клетки, называемое амитозом, так и
митоз — это непрямое дробление клетки, зиготы.
Ответом на вопрос будет вариант Г, митоз.
![]()
Зигота — клетка с полным, характерным для данного организма, набором хромосом, получающаяся в результате слияния двух половых клеток — яйцеклетки и сперматозоида.
Деление зиготы называется митозом. Митоз — это деление дроблением. Исходные клетки после каждого такого деления/дробления уменьшаются вдвое, а общее количество клеток, получающихся при делении, пропорционально увеличивается. Роста клеток, их увеличения между делениями, не происходит. Возникающие при дроблении зиготы клетки носят название бластомеров.
С помощью митоза делятся все бесполые клетки организма.
![]()
В результате слияния яйцеклетки и сперматозоида образуется оплодотворенная клетка с диплоидным, то есть удвоенным, набором хромосом. Эта оплодотворенная клетка и называется зиготой.
Через некоторое время после оплодотворения происходит деление зиготы. Способ деления, характерный для дробления зиготы, называется митоз.
![]()
Правильный ответ Г)МИТОЗ-это деление путем дробления.
Зигота-является клеткой с полным набором хромосом,который характерен для данного организма и получается в результате слияния яйцеклетки и сперматозоида.
Также с помощью Митоза делятся все бесполые клетки организма.
![]()
Зигота — клетка, которая образуется в результате слияния сперматозоида и яйцеклетки, то есть после оплодотворения. Самое первое дробление такой клетки называется дроблением (микозом). Название такое не случайно — зигота действительно дробится, каждая новая часть становится в 2 раза меньше по размеру и содержанию.
![]()
Митоз без сомнения подходит для ответа на этот вопрос, ведь зигота (исходная клетка новой особи) должна дробиться каждый раз пополам, чтобы постепенно вырасти в полноценное существо определенного вида животного или человека.
Источник
Онтогенез

![]()
Список вопросов теста
Вопрос 1
Варианты ответов
- бластоцель
- гастроцель
- миксоцель
- целлом
Вопрос 2
Какой способ деления клеток характерен для дробления зиготы
Варианты ответов
- амитоз
- мейоз
- митоз
- репликация
Вопрос 3
В результате дробления в эмбриогенезе образуется
Варианты ответов
- нейрула
- бластула
- гаструла
- зигота
Вопрос 4
Из какого зародышевого листка образуется нервная система и кожа животных
Варианты ответов
- мезодермы
- эктодермы
- энтодермы
- бластомеров
Вопрос 5
Установите последовательность процессов эмбриогенеза у ланцетника.
Варианты ответов
- дробление зиготы
- формирование бластулы
- образование гаструлы
- формирование трёх зародышевых листков
Вопрос 6
Установите соответствие между стадией онтогенеза и происходящих в них процессах
Варианты ответов
- двухслойный зародыш
- формирование многослойного зародыша
- первичный рот
- наличие мезодермы и хорды
- наличие бластоцели
- формирование нервной трубки
Вопрос 7
Благодаря непрямому развитию у животных ослабляется конкуренция между
Варианты ответов
- особями разных видов
- популяциями разных видов
- личинками и взрослыми формами
- взрослыми особями вида
Вопрос 8
Развитие комнатной мухи идёт по этапам:
Варианты ответов
- взрослая муха – яйцо – личинка – куколка
- взрослая муха – яйцо – куколка
- яйцо – личинка – куколка – муха
- яйцо – личинка – муха
Вопрос 9
Выберите три признака внутриутробного развития
Варианты ответов
- зародыш развивается в организме матери
- яйцеклетка развивается в воде
- эмбрион прикреплён к плаценте
- личинка питается другой пищей по сравнению со взрослым организмом
- характерен для птиц
- характерен для млекопитающих
Вопрос 10
Из какого зародышевого листка формируется сердечно-сосудистая система организма
Варианты ответов
- Эктодерма
- Энтодерма
- Мезодерма
- Бластомера
Получите комплекты видеоуроков + онлайн версии








Комментарии 0
Чтобы добавить комментарий зарегистрируйтесь или войдите на сайт
Возможности тестов
Источник
Лекция 7 Дробление

ЛЕКЦИЯ 7 Дробление
Общая характеристика процесса дробления.
1.Особенности митотических циклов в процессе дробления.
2. Правила дробления Гертвига-Сакса.
3. Полное (голобластическое), частичное (меробластическое) дробление.
4. Типы голобластического дробления.
5. Типы меробластического дробления.
6. Типы бластул животных с разным способом дробления, их строение и особенности.
Общая характеристика процесса дробления. Особенности митотических циклов в процессе дробления. Молекулярная природа факторов, определяющих укороченный клеточный цикл. Пространственная организация дробления. Правила Гертвига-Сакса. Полное (голобластическое), частичное (меробластическое) дробление. Основные типы голобластического дробления (радиальное, спиральное, билатеральное, анархическое) и их особенности. Основные типы меробластического дробления (дискоидальное, поверхностное) и их особенности.
Основные типы бластул у животных с разным способом дробления, их строение и особенности.
Биологическое значение и определения
Оплодотворение играет чрезвычайно важную роль в развитии, но оно лишь только первая его ступень. Зигота с ее новым генетическим потенциалом и новым распределением цитоплазмы приступает к созданию многоклеточного организма. У всех известных животных это начинается с процесса дробления.
Дробление — серия митотических делений, в результате которых огромный объем цитоплазмы яйца разделяется на многочисленные содержащие ядро клетки меньшего размера. Такие клетки называются бластомерами.
После объединения хромосомных наборов, являющегося следствием оплодотворения, без всякого перерыва начинается митотическое деление зиготы. За этим первым делением следует серия следующих делений ядер и цитоплазмы, общие свойства которых таковы:
- разделившиеся клетки зародыша не растут, т. е. в промежутке между делениями масса их цитоплазмы не увеличивается — в результате суммарный объем и масса всех возникших клеток не превышает объема и массы яйцеклетки во время оплодотворения; количество ДНК в ядре удваивается после каждого деления, как и при обычном митозе, так что все клетки сохраняют диплоидность.
Перетяжки, разделяющие дробящуюся яйцеклетку на все более мелкие клетки (бластомеры), называются бороздами дробления.
Дробление — это многократные митотические деления зиготы, в результате которых зародыш становится многоклеточным, не меняя при этом существенно своего объема.
Образование многоклеточности — первая и основная биологическая роль дробления. Вторая роль состоит в восстановлении ядерно-плазматического отношения, которое падает в ходе стадии большого роста ооцита.
Видовые особенности процесса дробления определяются двумя основными параметрами:
- количеством и распределением желточных белков в цитоплазме (желток подавляет дробление);
• присутствием в цитоплазме факторов, которые влияют на ориентацию митотического веретена и время его образования.
Дробление начинается вскоре после оплодотворения и заканчивается, когда у зародыша достигается новое равновесие между ядром и цитоплазмой. Дробление — строго координированный процесс, находящийся под генетическим контролем.
Отличие дробления от деления соматических клеток
В большинстве других случаев клеточной пролиферации в период между митозами происходит рост клеток. Клетка увеличивается в объеме почти вдвое и затем делится. Такой рост приводит к увеличению общего объема клеток при сохранении относительно постоянного отношения объема ядра к объему цитоплазмы.
В период дробления зиготы объем цитоплазмы не возрастает: огромная масса цитоплазмы зиготы разделяется на все более мелкие клетки. Это деление цитоплазмы яйца, не сопровождающееся ростом, осуществляется путем выпадения G1-периода в интерфазе, тогда как митозы следуют друг за другом с большой скоростью.
Темпы увеличения числа клеток в период дробления намного выше, чем на стадии гаструляции. Одним из последствий высокой интенсивности делений в процессе дробления является постепенное уменьшение отношения объема цитоплазмы к объему ядра. Изменение скорости, с которой происходит снижение отношения объема цитоплазмы к объему ядра, у многих типов зародышей является решающим фактором, определяющим время активации некоторых генов.
От митотического деления соматических клеток дробление отличается тем, что полученные в результате дробления клетки не растут, а поэтому с каждым следующим делением становятся всё более мелкими, при этом увеличивается только их количество, а зародыш в целом не растет. Получающиеся при дроблении клетки малодифференцированны и сравнительно однородны.
Период синхронных делений дробления характеризуется укороченными клеточными циклами, из которых фактически выпадает пресинтетический, или G1-период, а также постсинтетический, или G2-период (рис. 19).
Глубокие изменения в клеточном метаболизме частично связаны с циклическим чередованием окисленной и восстановленной конформации белков, обусловленной S-S и S-H группами. Свободные сульфгидрильные группы в наибольшем количестве имеются в делящихся бластомерах и в наименьшем в то время, когда клетки не делятся.

Рис. 19. Изменение клеточного цикла при дроблении у амфибий: А — нормальный клеточный цикл; Б—клеточный цикл при дроблении
Микрофиламенты (сократимой кольцо)

Рис. 20. Схема расположения микротрубочек и микрофиламентов при клеточном делении (по Гилберту, 1993)
Высокая скорость делений яйцеклеток объясняется следующим:
- в яйцеклетках заранее запасены (в период оогенеза) непосредственные предшественники ДНК (цитидин, тимитидин-3-фосфаты, а также ядерные белки гистоны) и мРНК, а в других клетках таких запасов нет; ДНК синхронно делящихся бластомеров имеет значительно больше точек инициации репликации, нежели у других клеток эукариот.
Дробление является результатом двух координированных процессов — кариокинеза (митотическое деление ядра) и цитокинеза (деление клетки). Механическим аппаратом кариокинеза является митотическое веретено с его микротрубочками, состоящими из тубулина, а цитокинеза — сократимое кольцо микрофиламентов, состоящих из актина. Микротрубочки распределяют хромосомы по центриолям, тогда как в результате сокращения микрофиламентов происходит перешнуровывание цитоплазмы (рис. 20).
Обычно кариокинез и цитокинез координированы между собой. Локализация борозд дробления определяется положением звезд митотического веретена, а число борозд зависит от числа последних. Дробление протекает нормально, если яйцо содержит две звезды.
При дроблении зародыша происходит образование новых клеточных мембран посредством двух механизмов:
- синтез мембран de novo; растягивание плазматической мембраны ооцита.
За невероятным многообразием типов дробления скрывается общность функций и механизмов. Во всех случаях кариокинез и цитокинез должны быть скоординированы и яйцо разделено на клеточные области. В итоге восстанавливается характерное для соматических клеток ядерно-плазменное отношение и важная для развития информация распределяется между разными клеточными областями.
Клеточный цикл соматических клеток делится на четыре стадии (фазы). За митозом (М) следует пресинтетический период (G1), после которого происходит синтез ДНК (S). Затем наступает премитотическая фаза (G2), за которой опять следует митоз.
Высокая пролиферативная активность во время дробления обусловлена прежде всего укороченными клеточными циклами. В период синхронных делений дробления они отличаются отсутствием G1 периода, составляющего у обычных делящихся клеток значительную часть клеточного цикла.
Фаза синтеза ДНК занимает непродолжительный отрезок времени, а удельная доля митоза составляет, как правило, около 50 % (в отдельных случаях до 70-75 %) генерационного времени. Таким образом, главные синтетические процессы, протекающие в синхронно делящихся бластомерах, — синтез ДНК и гистонов.
Укорочение клеточных циклов при дроблении происходит потому, что в G1 фазе дробящихся эмбрионов отсутствует экспрессия генов, т. е. геном зародыша полностью неактивен. Все синтетические процессы, в том числе и синтез гистоновых белков, идут за счет материнских матричных РНК, накопленных еще в оогенезе. Исключение из этого правила составляют животные с асинхронным дроблением. Так, у млекопитающих некоторые гены эмбриона начинают экспрессироваться уже на стадии двух бластомеров.
Клеточный цикл бластомеров на ранних стадиях дробления в отличие от нормального клеточного цикла может быть гораздо более простым — двухфазным.
Остановка развития на стадии метафазы

Рис. 21. Модель регуляции клеточного цикла бластомеров (по Гилберту, 1993)
Факторы, регулирующие этот цикл, локализованы в цитоплазме. Это те же самые факторы, которые регулировали деления созревания при оогенезе: фактор, стимулирующий созревание (maturation promoting factor, MPF), цитостатический фактор (сytostatic factor, CSF) и ионы кальция.
Показано, что в делящихся клетках уровень активности MPF претерпевает циклические изменения. Активность MPF в бластомерах лягушки на ранних стадиях дробления наивысшая в М-фазе и не выявляется в S-фазе.
Действие MPF осуществляется путем изменения структуры ядерной оболочки. Цитостатический фактор стабилизирует фактор созревания, задерживая клетки в состоянии митоза, а кальций инактивирует цитостатический фактор, стимулируя переход к S-фазе за счет инактивации MPF. При добавлении CSF прекращаются также циклические сокращения кортикального слоя цитоплазмы, а последующая инъекция ионов кальция их стимулирует (рис. 21).
На ранних стадиях развития цитоплазма определяет скорость клеточных делений и продолжительность S — и M-фаз. В период асинхронных делений дробления появляется фаза G1, удлиняется продолжительность всех остальных фаз цикла. Начинается синтез различных видов РНК на матрицах ДНК, т. е. пробуждается транскрипционная активность генома зародыша.
Считается, что потеря синхронности дробления связана именно с активацией генома зародыша. В развитии всех животных наступает момент, начиная с которого темпы клеточной репродукции замедляются и происходит десинхронизация делений дробления. Изменение структуры клеточного цикла сопровождается увеличением подвижности бластомеров во время удлинившейся интерфазы и началом транскрипции собственных генов зародыша. Этот процесс контролируется ядерно-цитоплазматическим отношением, которое постепенно увеличивается при дроблении.
Гены, внесенные в геном зародыша со сперматозоидом, проявляют свое действие именно в этот период и, во всяком случае, не раньше окончания периода синхронного дробления. Именно в это время зародыш берет свою судьбу в собственные руки и перестает быть генетической копией матери. Поскольку период асинхронности начинается после разного числа делений дробления, то и пробуждение транскрипционной активности начинается при соответственно разном количестве бластомеров: у млекопитающих и круглых червей практически с самого начала развития, у иглокожих — со стадии 32 бластомеров, у амфибий — со стадии бластулы.
Пространственная организация зародыша во время дробления. Борозды дробления
Закономерности, связанные с наличием и распределением желтка в яйцеклетке и направлением хода борозд дробления, определяются двумя правилами Гертвига — Сакса:
- клеточное ядро стремится расположиться в центре чистой, свободной от желтка цитоплазмы; веретено клеточного деления стремится расположиться по направлению наибольшего протяжения свободной от желтка цитоплазмы.
Наиболее общей закономерностью голобластического (см. ниже) дробления мезо — и олиголецитальных яиц является взаимная перпендикулярность (ортогональность) первых трех борозд, причем две проходят по меридианам яйца, а третья — по экватору.
Борозды дробления (рис. 22):

Рис. 22. Ход борозд дробления (по Гилберту, 1993): А — первая меридиональная борозда; Б — вторая меридиональная борозда; В — широтная борозда
- меридиональная — проходит от анимального полюса зиготы к вегетативному; экваториальная (ее аналогом является широтная борозда, смещенная по отношению к экваториальной в направлении к анимальному полюсу из-за перегруженности желтком вегетативного полюса) — проходит по экватору зиготы; тангенциальная — проходит параллельно поверхности зиготы, в результате чего образуется многослойный зародыш.
Следует заметить, что скорость прохождения борозд дробления всегда обратно пропорциональна количеству желтка в клетке (иногда данное положение называют третьим правилом дробления). Дробление у разных многоклеточных протекает различно. Это зависит от особенностей строения яйцеклеток, прежде всего от количества и расположения в них желтка, что и лежит в основе классификации типов дробления.
Классификация типов дробления
Существует несколько типов классификации процесса дробления.
По характеру образования и расположению бластомеров:
- полное (голобластическое) — характерно для зигот, содержащих мало желтка (мезо — и изолецитальные яйца), при этом борозды дробления проходят через все яйцо, а имеющийся у них желток включается в вегетативные бластомеры; неполное (меробластическое) — характерно для зигот, содержащих большие запасы белков желтка (полилецитальные яйца), при этом борозды дробления не проникают в богатую желтком область цитоплазмы.
В зависимости от размеров образовавшихся бластомеров:
- равномерное — бластомеры на анимальном и вегетативном полюсе имеют одинаковые размеры; неравномерное — на анимальном полюсе сосредоточены более мелкие бластомеры, чем на вегетативном.
По скорости формирования бластомеров:
- синхронное — при одинаковой скорости образования бластомеров на обоих полюсах зиготы; асинхронное — на анимальном полюсе скорость образования бластомеров выше, чем на вегетативном.
Выделяют четыре основных типа голобластического дробления. Данная классификация основана на взаимном пространственном расположении бластомеров:
- радиальное; спиральное; билатерально-симметричное; неправильное (анархическое).
Радиальный тип дробления присущ голобластическим хордовым (ланцетник, круглоротые, осетровые рыбы, амфибии), иглокожим и некоторым другим группам.
При этом типе дробления бластомеры разных широтных ярусов располагаются, по крайней мере на ранних стадиях, довольно точно один над другим, так что полярная ось яйца служит осью поворотной симметрии.
Радиальный равномерный тип дробления характерен для яиц иглокожих (рис. 23).
У яйца лягушки наблюдается радиальный неравномерный тип дробления. Борозда первого деления дробления еще не завершила разделения богатой желтком цитоплазмы вегетативного полушария, а борозды второго деления уже закладываются вблизи от анимального полюса. Из-за большой концентрации желтка в вегетативной области борозды третьего деления дробления располагаются значительно ближе к анимальному полюсу (рис. 24).
В результате возникают область быстро делящихся бластомеров вблизи анимального полюса и область более медленно делящихся бластомеров вегетативного полюса.

Рис. 23. Голобластическое дробление иглокожих (по Гилберту, 1993): А, Б, В — последовательные стадии процесса

Рис. 24. Дробление яйца лягушки (по Гилберту, 1993): А — первое деление; Б — второе деление; В — четвертое деление

Рис. 25. Схема начальных стадий спирального дробления
(по Мануйловой, 1973): А — переход от 4 к 8 бластомерам (в клетках видны веретена деления); Б — стадия 8 бластомеров; В — переход от 8 к 16 бластомерам; Г — стадия 16 бластомеров
Спиральный тип дробления характеризуется утерей элементов симметрии уже на стадии четырех, а иногда и двух бластомеров и присущ беспозвоночным (моллюски, кольчатые и ресничные черви), объединяемым в группу Spiralia.
Свое название этот тип дробления получил из-за того, что при взгляде с анимального полюса последовательно отделяющиеся четверки (квартеты) бластомеров поворачиваются относительно анимально-вегетативной оси то в правую, то в левую сторону, как бы образуя при наложении друг на друга спираль (рис. 25).
Знак спирального дробления, его дексио-(право-) или лео-(лево-) тропность, т. е. «закрученность», определяется геномом матери данной особи. Оно во многом отличается от радиального типа дробления.
Во-первых, яйца не делятся параллельно или перпендикулярно анимально-вегетативной оси. Плоскости делений дробления ориентированы наклонно, что приводит к спиральному расположению дочерних бластомеров.
Во-вторых, число контактов между клетками больше, чем при радиальном дроблении. В-третьих, зародыши со спиральным типом дробления проходят меньше делений до начала гаструляции. Возникающие таким образом бластулы обычно не имеют бластоцели (стерробластула).
Билатеральный тип дробления (круглые черви, оболочники) характеризуется наличием одной плоскости симметрии. Наиболее примечательная особенность этого типа дробления заключается в том, что плоскость первого деления устанавливает единственную плоскость симметрии зародыша (рис. 26).
Каждое последующее деление ориентируется по отношению к этой плоскости симметрии так, что половина зародыша по одну сторону от первой борозды представляет собой зеркальное отражение половины зародыша по другую ее сторону.

Рис. 26. Дробление яйца аскариды (по Мануиловой, 1973): А — стадия двух бластомеров (в клетках видны веретена следующих делений); Б — стадия четырех бластомеров до поворота вегетативной пары; В — начало поворота вегетативной пары бластомеров; Г—ромбическая фигура из четырех бластомеров после завершения поворота (последовательные поколения предков половых клеток — Рп, где п — номер поколения)
рис. 27. Анархическое дробление (по Токину, 1987)
При билатеральном типе дробления формируется одна плоскость симметрии: первая борозда проходит экваториально, далее анимальный бластомер делится меридиональной бороздой, а вегетативный — широтной. В результате получается Т-образная фигура из четырех бластомеров, не обладающая поворотной симметрией.
Путем поворота вегетативной пары бластомеров Т-образная фигура преобразуется в ромбическую. Этот поворот происходит в промежутке между делениями, в интерфазе.
Анархический тип дробления присущ кишечнополостным и паразитическим плоским червям. Он характеризуется тем, что бластомеры слабо связаны между собой и располагаются неправильными цепочками (рис. 27).
При этом они могут распадаться, например под ударами волн, но из отдельных участков образуются полноценные зародыши. В результате плотного объединения бластомеров друг с другом в конце дробления образуется морула.
Основными типами меробластического дробления являются:
При поверхностном дроблении после слияния пронуклеусов ядро зиготы делится на много ядер, которые с небольшим количеством цитоплазмы по цитоплазматическим мостикам переходят во внешний слой свободной от желтка цитоплазмы (периплазму) и равномерно там распределяются
(речь идет о центролецитальных яйцеклетках). Здесь ядра еще несколько раз синхронно делятся, располагаясь довольно близко друг к другу (рис. 28).
На этой стадии, еще до возникновения клеточных перегородок (так называемой синцитиальной бластодермы), ядра окружаются особыми структурами из микротрубочек, затем деление ядер становится асинхронным, между ними формируются клеточные перегородки и образуется базальная мембрана, отделяющая периплазму от центральной массы желтка. Борозды дробления появляются, но они не заходят глубоко в яйцо. Возникший поверхностный слой клеток называется клеточной бластодермой. Этот тип дробления характерен для большинства насекомых.

Дискоидальный тип дробления присущ оплодотворенным полилецитальным и телолецитальным яйцеклеткам рыб, рептилий и птиц (рис. 29).

Рис. 28. Стадии поверхностного дробления (по Белоусову, 1993): А, Б—ядра дробления постепенно переходят на поверхность клетки; В, Г — образование перибласта

Рис. 29. Дискоидальное дробление куриного яйца (по Гилберту, 1993). Представлен вид со стороны анимального полюса. Борозды дробления не распространяются на желток, а возникающая бластодерма состоит из одного слоя клеток
Первые две борозды проходят перпендикулярно друг другу, но далее строгий порядок прохождения борозд нарушается. При этом на бластомеры делится лишь тонкий диск цитоплазмы (бластодиск), расположенный на анимальном полюсе.
У многих яйцеклеток еще на ранних стадиях дробления внутренние концы бластомеров расходятся и между ними возникает небольшая, постепенно увеличивающаяся полость дробления (бластоцель). У некоторых типов яйцеклеток бластоцель может достигать значительных размеров. Зародыш на этой стадии развития называется бластулой. В ходе дальнейшего развития бластоцель превращается в первичную полость тела, которая является основной полостью тела у низших беспозвоночных. У высших беспозвоночных и позвоночных она почти полностью вытесняется возникающей позже вторичной полостью тела (целомом).
Бластоцель выполняет две функции:
- дает возможность клеткам при гаструляции мигрировать внутрь зародыша; существование бластоцели предотвращает взаимодействие между клетками, которые находятся выше и ниже ее.
Бластоцель — первый возникающий по ходу развития отсек внутренней среды организма, отличающийся по ионному составу от наружной среды. Клетки стенок бластоцеля, отгораживающие его от наружной среды, образуют между собой плотные контакты. В обращенной к внешней среде мембране клеток расположены ионные каналы для натрия, хлора и других ионов, обеспечивающие их перенос по градиентам концентрации, а в мембране, обращенной в бластоцель, — ионные насосы, работающие против градиента. В результате Na и Cl — перекачиваются в бластоцель, где возникает их избыток, что создает в бластоцеле повышенное осмотическое давление и обеспечивает перенос воды, повышая в ней тургорное давление. Это давление растягивает поверхность зародыша, что важно для его последующего развития. Избыток в бластоцеле может оказывать влияние на скорость клеточных циклов и стимулировать экспрессию генов в клетках бластулы.
Имеется 5 основных типов бластул:
- целобластула; бластоциста; амфибластула; дискобластула; перибластула.
Целобластула (от греч. koilos — пустой) характерна для иглокожих и ланцетника (рис. 30, А). Этот тип бластул отличается тем, что бластомеры на анимальном и вегетативном полюсе почти одинаковы по размеру, тонкую бластодерму формирует один слой клеток

Рис. 30. Типы бластул (по Мануйловой, 1973; Токину, 1987; Белоусову, 1989): А — целобластула, поперечный разрез (морской огурец); Б — стерробластула, поперечный разрез (ставромедуза); В — стомобластула, поперечный разрез (известковые губки)
Полость внутри — бластоцель. Выделяют в целобластуле следующие зоны: крыша на анимальном полюсе, дно — на вегетативном, между ними находится краевая, или промежуточная, зона.
Бластула со стенкой равномерной толщины и очень маленьким центрально расположенным бластоцелем (рис. 30, Б), встречаемая у некоторых кишечнополостных, моллюсков и червей, называется стерробластулой (от греч. sterros — твердый, плотный).
Зародышевое развитие некоторых червей, асцидий приводит к образованию плакулы. Она имеет вид двухслойной пластинки, образованной однородными клетками. Между слоями располагается полость дробления (бластоцель).
У некоторых кишечнополостных, а также у млекопитающих на ранних стадиях развития бластоцель вообще не возникает, и дробление приводит к образованию плотного скопления клеток — морулы (от лат. morum — тутовая ягода) (рис. 31, А).

Рис. 31. Типы бластул (по Токину, 1987 и Белоусову, 1993): А — морула, внешний вид (мышь); Б — бластоциста, поперечный разрез (летучая мышь)
Своеобразным типом бластулы является стомобластула (от греч. stoma — рот), характерная для зародышевого развития известковых губок (рис. 30, В). Она имеет полость в центре и отверстие (фиалопор) на анимальном полюсе. Жгутикообразующие полюсы клеток обращены внутрь. По окончании дробления стомобластула выворачивается наизнанку через фиалопор (экскурвация), в результате чего образуется покрытая жгутиками амфибластула.
Бластоциста характерна для млекопитающих (рис. 31, Б). Образуется после прохождения стадии морулы.

Она представлена в виде пузырька (от греч. kystis — пузырь). Причем в составе этого пузырька имеются две части. Стенку формирует один слой клеток — трофобласт. Клетки эти очень светлые, другая часть — темноокрашенные клетки эмбриобласта. Трофобласт обеспечивает питание, а эмбриобласт идет на построение тела зародыша.
Амфибластула характерна для амфибий и некоторых представителей известковых губок. В отличие от целобластулы бластоцель в амфибластуле смещена к анимальному полюсу вследствие большего количества желтка на вегетативном полюсе. На анимальном полюсе находятся меньшие по размеру клетки — микромеры. В области вегетативного полюса располагаются макромеры. Так же как и в целобластуле, в амфибластуле выделяют крышу, дно и промежуточную (краевую) зону (рис. 32).
Дискобластула характерна для костистых рыб, рептилий и птиц. В результате дискоидального дробления образуется однослойная бластодерма (бластодиск), из которого затем возникает трех — или четырехклеточный пласт.

Рис. 33. Дискобластула — образование гипобласта в яйце птицы
(по Гилберту, 1993): А — бластодерма, преобразованная в трех — или четырехслойный клеточный пласт; Б—начало выселения клеток бластодермы в подзародышевую полость и формирование гипобласта; В — завершение образования дис кобластулы: формирование эпи — и гипобласта, бластоцели
Бластодерма дискобластулы несколько выгибается над желтком, и между ними формируется подзародышевая полость. На этой стадии четко различимы две области бластодиска: светлое поле (areapellucida) соответствует той его части, которая располагается над полостью; темное поле (area opaca) состоит из клеток, расположенных по краю бластодиска и прилежащих к желтку (рис. 33, А).
К моменту откладки яйца некоторые из клеток бластодермы выселяются в подзародышевую полость, где образуют второй слой (рис. 33, Б). Таким образом, на данной стадии зародыш состоит из эпибласта (наружный слой клеток) и лежащего под ним гипобласта. Между ними находится бластоцель (рис. 33, В).
Перибластула характерна для насекомых и представляет собой заключительную стадию поверхностного дробления. Энергиды (ядра с окружающими их островками цитоплазмы) мигрируют на поверхность яйца, где они окружаются новыми плазматическими мембранами. Первыми претерпевают изменения ядра, мигрировавшие к заднему полюсу яйца и образующие полярные клетки зародыша, которые дают начало половым клеткам взрослого организма. Клетки образующейся бластодермы формируют скопление на вентральной поверхности, обозначая место, где возникнут зародышевые листки (зародышевая полоска). Эта область дает начало всем клеткам насекомого. Остальные клетки бластодермы образуют внезародышевые оболочки. Впоследствии именно клеточная организация бластулы определит характер дальнейшего развития зародыша.
Источник



