
РОСТ И КУЛЬТИВИРОВАНИЕ БИООБЪЕКТОВ — В. М. Самыгин — 2016
ГЛАВА 4. ОСНОВНЫЕ МЕХАНИЗМЫ ОБМЕНА ВЕЩЕСТВ И ПРЕОБРАЗОВАНИЯ ЭНЕРГИИ У ПРОКАРИОТ
В клетках микроорганизмов постоянно происходит множество биохимических реакций, которые составляют в совокупности метаболизм (обмен веществ), направленный на сохранение, воспроизведение жизни и обеспечивающийся за счет процессов питания и дыхания. Вся совокупность процессов метаболизма подчиняется «принципу биохимического единства» (Клювер, 1926), согласно которому все живые существа на Земле в биохимическом отношении сходны: у них обнаружено сходство строительных блоков, единый энергетический аккумулятор — АТФ, универсальный генетический код и идентичные в своей основе главные метаболические пути.
Особенностью метаболизма микроорганизмов являются:
— голофитный способ питания, при котором питательные вещества должны быть растворены в жидкости;
— поступление питательных веществ по всей поверхности клетки, которая очень велика по сравнению с величиной самого микроорганизма;
— необычайная интенсивность процессов обмена веществ (за сутки при благоприятных условиях одна клетка потребляет пищи в 30-40 раз больше массы своего тела);
— многообразие источников энергии и пластических материалов;
— высокая адаптация к изменяющимся условиям окружающей среды.
Поступающие в клетку питательные вещества используются по двум основным направлениям. Часть их используется в реакциях расщепления и окисления с получением энергии и запасанием ее в форме АТФ или других энергоемких соединений — энергетический обмен (катаболизм). Другая часть расходуется по путям конструктивного, биосинтетического обмена (анаболизма). Эти процессы являются сопряженными и лишь отражают разные стороны метаболизма. Конечные продукты этих реакций называются метаболитами. На определенных этапах метаболизма протекают промежуточные реакции перестройки одних веществ в другие. Они получили название реакций амфиболизма, а их продукты — амфиболиты — могут использоваться как в процессах анаболизма, так и катаболизма.
Поступающие в клетку питательные вещества сначала расщепляются на небольшие фрагменты, а затем в ходе реакций амфиболизма превращаются в ряд органических кислот и фосфорных эфиров. Эти два пути переходят незаметно один в другой. Многообразные низкомолекулярные соединения — это тот субстрат, из которого синтезируются «строительные блоки», то есть аминокислоты, пуриновые и пиримидиновые основания, фосфорилированные сахара, органические кислоты и другие метаболиты — конечные продукты цепей биосинтеза. Из них строятся полимерные макромолекулы — нуклеиновые кислоты, белки, резервные вещества, компоненты клеточной стенки и тому подобные, из которых состоит клетка. Эти два этапа биосинтеза клеточных веществ — синтез строительных блоков и синтез полимеров — составляют синтетическую ветвь метаболизма, или анаболизм.
Окисление глюкозы, как основного амфиболита, осуществляется, как правило, по трем путям:
— гликолиз, или путь Эмбдена-Мейергофа-Парнаса, характеризуется образованием из одной молекулы глюкозы двух молекул пировиноградной кислоты (пирувата), синтезом трех молекул пиридиннуклеотидов и двух молекул АТФ;
— пентозофосфатный путь (гексозомонофосфатный, или путь Варбурга-Диккенса-Хорекера) у микробов играет вспомогательную роль. В результате его реакций образуются пентозы для последующих синтезов, пируват, три молекулы пиридиннуклеотида и две молекулы АТФ;
— кетодезоксифосфоглюконатный (КДФГ, или путь Энтнера- Дудорова) присущ исключительно микроорганизмам и приводит к образованию из одной молекулы глюкозы двух молекул пирувата, одной АТФ и одной НАДФ.
Глюкоза после ряда предварительных этапов расщепляется «пополам», а продукты расщепления превращаются в пируват, который занимает ключевое положение в промежуточном метаболизме, так как служит исходным соединением во многих процессах синтеза и распада. В результате декарбоксилирования пирувата образуются С2-соединения, которые связываются сначала с акцепторной молекулой (оксалоацетатом), а затем в цикле трикарбоновых кислот, называемом также циклом лимонной кислоты, постепенно окисляются до СО2 и Н2О. Оксалоацетат (щавелевоуксусная кислота) в этом циклическом процессе регенерируется. Атомы водорода (или восстановительные элементы), отщепившись на разных этапах окисления органических веществ, поступают в АТФ-регенирирующую систему дыхательной цепи (окислительное фосфорилирование). При каждом обороте цикла трикарбоновых кислот из одного С2-соединения (ацетилкоэнзим А — активированная уксусная кислота) образуются две молекулы СО2 и восемь протонов водорода. Эти реакции выравнивают баланс цикла трикарбоновых кислот.
К промежуточным соединениям цикла трикарбоновых кислот относятся органические кислоты, поставляемые для процессов биосинтеза (2-оксоглутарат, сукцинат, оксалоацетат). Цикл трикарбоновых кислот не только участвует в конечном окислении питательных веществ, но и служит «распределителем», поставляющим исходные соединения для синтеза основных структурных единиц («строительных блоков») клетки.
Если бы указанные кислоты постоянно выводились из цикла, то регенерации молекулы-акцептора не происходило бы, и цикл был бы нарушен. Однако анаплеротические последовательности реакций обеспечивают поступление в цикл трикарбоновых кислот все новых количеств промежуточных соединений, взамен израсходованных для биосинтеза. Эти последовательности имеют особо важное значение для организмов, растущих за счет простых одно- или двухуглеродных соединений.
Все процессы жизнедеятельности микробной клетки сопряжены с тратой энергии и требуют ее возобновления, которое осуществляется за счет экзотермических биохимических реакций окисления различных органических и неорганических веществ. Эти процессы энергетического обмена (катаболизма), восстанавливающие энергию клетки, обусловлены дыханием. При этом потребляются те же соединения, которые идут на построение структурных компонентов клетки, — сахара, спирты, органические кислоты, жиры и др.
Окисление веществ может происходить разными путями:
— прямым, когда к веществу присоединяется кислород. В этом случае при помощи оксидаз происходит непосредственное окисление атмосферным кислородом неорганического субстрата — молекулярного водорода, окиси углерода или серы. Прямое окисление регистрируется у большинства сапрофитов;
— непрямым — отделением от субстрата двух атомов водорода или двух электронов (е — ). Этот процесс называется дегидрированием. Отнятый от окисляемого субстрата-донора водород (е — ) переносится на другое вещество, которое при этом восстанавливается. Вещество, присоединяющее водород (е — ), называют акцептором. Процесс переноса водорода и электронов эквивалентен. Он сопровождается высвобождением энергии, которая утилизируется клеткой и аккумулируется в макроэргических соединениях (преимущественно АТФ). Нередко термин «донор водорода» и «донор электронов» употребляются как синонимы; равнозначны и такие термины, как «акцептор водорода» и «акцептор электронов»; «окисление» и «дегидрирование», «восстановление» и «гидрирование».
Ферментные белки, отщепляющие от субстратов атомы водорода, называются дегидрогеназами. У бактерий в этом процессе участвуют не
сколько дегидрогеназ, и их называют по донору водорода (например, лактатдегидрогеназа, малатдегидрогеназа). Они переносят водород на один из двух коферментов — никотинамидадениндинуклеотид (НАД + ) или никотинамидадениндинуклеотидфосфат (НАДФ + ). Эти коферменты легко отделяются от белка-дегидрогеназы и переносят водород после связывания с другой дегидрогеназой на другой акцептор. Поэтому их называют также переносчиками водорода.
Процессы, требующие затраты энергии, осуществляются благодаря АТФ. АТФ (аденозинтрифосфат) является основным носителем биологически используемой энергии («энергетическая валюта» клетки), то есть универсальным переносчиком химической энергии между реакциями, доставляющими и потребляющими энергию. Все энергетические процессы в живых клетках прямо или косвенно сопряжены с превращением АТФ в АДФ и неорганический фосфат (Р). При разрыве фосфатных связей в АТФ выделяется большое количество энергии, которая и запасается в продуктах реакции.
Существует два основных механизма образования АТФ в клетках:
— мембранное окислительное фосфорилирование, осуществляющееся путем отнятия электронов от органического или неорганического субстрата (его окисление) и передачи их конечным акцепторам, которые при этом восстанавливаются. Причем конечными акцепторами являются неорганические соединения — кислород (у аэробов), сульфаты, нитраты, карбонаты и др. (у анаэробов). Эта передача электронов осуществляется в мезосомах — мембранных структурах, где локализованы ферменты дыхания, и происходит путем переноса электронов в электронно-транспортной (дыхательной) цепи (ЭТЦ). Классическим примером такого механизма образования АТФ является цикл Кребса. За один оборот такого цикла, как отмечалось, выделяется две молекулы углекислоты, восемь восстановительных эквивалентов и одна молекула АТФ;
— субстратное фосфорилирование (брожение). При этом пути донорами и акцепторами электронов являются органические соединения (чаще всего гексозы), окисление которых происходит в циклических процессах (цикле трикарбоновых кислот и его вариантах, пентозофосфатном цикле и др.).
У бактерий наиболее частым путем окисления является дегидрирование. При этом перенос водорода (е — ) осуществляется окислительно-восстановительными ферментами (оксидоредуктазами), а конечным акцептором Н2 (е — ) может быть кислород или другое вещество, способное восстанавливаться:

В соответствии с путями биологического окисления бактерии объединяют в несколько групп:
— облигатные аэробы — бактерии, у которых конечным акцептором водорода (е — ) является свободный молекулярный кислород воздуха;
— микроаэрофилы — бактерии-аэробы, у которых рост и размножение оптимальны при сниженном парциальном давлении кислорода. К ним относятся лептоспиры, актиномицеты и др. Некоторые из бактерий — микроаэрофилов (например, нейссерии, бруцеллы) лучше растут при повышенном содержании СО2;
— облигатные анаэробы — бактерии, которые получают энергию при окислении органических или неорганических (водород, сера, железо, сероводород и др.) веществ без доступа свободного кислорода воздуха или анаэробным фотосинтезом;
— факультативные анаэробы, способные извлекать энергию из субстратов аэробным и анаэробным путями биологического окисления в зависимости от источника кислорода.
В энергетическом отношении аэробный тип дыхания является наиболее эффективным. В аэробных условиях происходит окисление субстрата до СО2 и воды или перекиси водорода. Процесс этот многоэтапный, протекает при участии ряда ферментов (цитохромоксидазы и системы геминовых ферментов цитохромов) с образованием различных промежуточных продуктов. Отнятые при этом электроны передаются по дыхательной цепи через ряд окислительных стадий до активированного кислорода. Свободный кислород выполняет функцию конечного акцептора отнятых электронов. Этот процесс идет с выделением энергии и сопровождается образованием АТФ. К аэробным микроорганизмам относят микобактерии, вибрионы, бациллы и др. Для их культивирования используют питательные среды с высокой аэрацией, а при идентификации бактерий определяют наличие ферментов дыхания: оксидаз, каталазы, цитохромов и др.
Под анаэробным дыханием понимают такие пути биологического окисления, при которых конечным акцептором электронов являются органические (фумарат) или неорганические вещества (нитраты, сульфаты, сера, железо, карбонаты), но не свободный кислород воздуха. Все виды анаэробного дыхания подразделяются по используемому конечному акцептору электронов (табл. 2).
Таблица 2. Виды анаэробного дыхания
Источник
Типы биологического окисления
Энергетический метаболизм
Различных способов получения энергии у гетеротрофов
Сравнительная эффективность
| Пути катаболизма | Выход АТФ |
| Гликолиз, использует большинство микроорганизмов, основной способ у анаэробов | 2 молекулы |
| Лимонно-кислый (цикл трикарбоновых кислот), широко используется | 34 молекулы |
| Фосфоглюконатный | 1 молекула |
| Петозофосфатный | нет прямой продукции АТФ |
| Ферментация, используется анаэробами | 2 или 3 молекулы |
| Кетодезоксифосфоглюконатный | 1 молекула |
Процесс получения энергииу прокариотов—дыхание (биологическое окисление), в результате которого синтезируются молекулы АТФ. У некоторых микроорганизмов аккумулятором энергии могут быть и другие фосфаты, иногда даже неорганические.
Дыхательный аппарат бактерийсостоит из мезосом — инвагинаций ЦПМ, где локализованы ферменты-оксидоредуктазы, с помощью которых происходит биологическое окисление:
– пиридиновые дегидрогеназы есть у всех микроорганизмов, отнимают от субстрата водород и таким образом происходит окисление. У облигатных анаэробов есть только эти ферменты;
– флавиновые ферменты широко распространены и делятся на 2 группы: флавинодегидрогеназы (окисляют субстрат, отнимая у него водород) и ферменты переноса (только передают водород, забирая его у пиридиновых дегидрогеназ);
– железосодержащие ферменты (цитохромы) — цитохромоксидаза, пероксидаза, каталаза — сами субстрат не дегидрогенируют, а выполняют роль переносчика водорода на кислород;
– медьсодержащих ферментов мало (фенолоксидаза). Они сами дегидрогенируют субстрат и переносят водород на кислород.
Кроме ферментов, локализованых в мезосомах у микроорганизмов есть свободные переносчики водорода (убихиноны, аскорбиновая кислота). Они вступают в действие, когда мембрана растягивается и наблюдается разрыв между локализованными ферментами.
В зависимости от того, что является конечным акцептором электронов, выделяют три типа образования энергии у бактерий: аэробное дыхание, анаэробное дыхание и ферментацию (брожение)(рис. 57, 58).
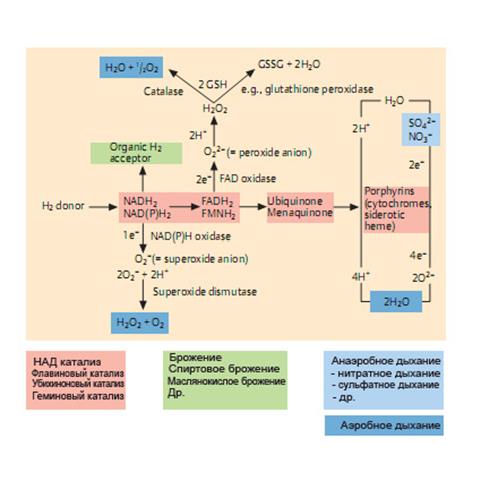
Рис. 57. Типы биологического окисления
1. Аэробный(от греч. аer — воздух и bios — жизнь) тип— окислительное фосфорилирование— совокупность экзотермических процессов ферментативного распада органических веществ (реже — неорганических), происходящих с участием свободного кислорода.
Процесс дыхания у аэробов протекает по типу окислительной реакции путем отщепления от субстратов водорода (или электронов). Чаще наблюдается гликолиз — процесс ферментативного расщепления углеводов. Пировиноградная кислота, образовавшаяся в результате гликолиза, окисляется в цикле трикарбоновых кислот, который снабжает аэробов предшественниками для реакций биосинтеза.
Водородные атомы передаются вдоль цепи молекул-переносчиков, которые образуют цепь переноса электронов, или «дыхательную» цепь. Водород и электроны отщепляются от субстратов с помощью пиридиновых дегидрогеназ,связанных с мембранами. Далее поток электронов направляется от системы с более низким (более отрицательным) потенциалом к системе с более высоким (более положительным) потенциалом: флавопротеидные ферменты — цитохромные ферменты — цитохромоксидаза. На последней ступени этой реакции, катализируемой цитохромоксидазой, ионы водорода связываются с молекулярным кислородом — конечным акцептором электронов. При этом образуется перекисный радикал — очень сильный окислитель. Фермент супероксиддисмуатаза переводит перекисный радикал в перекись водорода, которая ферментами каталазойили пероксидазойпреобразуется в безвердные продукты — воду и кислород. Таким образом, специфические ферменты защищают микробную клетку от токсических продуктов окисления.
Аэробное дыхание энергетически более эффективно. Перенос электронов вдоль дыхательной цепи сопряжен с образованием богатых энергией фосфатных связей в молекулах АТФ. Образуется также много промежуточных и конечных продуктов метаболизма, которые используются для синтеза белков, углеводов, витаминов. Филогенетически аэробное дыхание возникло позже.
Облигатным (строгим) аэробамдля дыхания необходим молекулярный (атмосферный) кислород. Они не могут жить и размножаться в отсутствие молекулярного кислорода, используют только аэробный путь окисления (некоторые виды псевдомонад, холерный вибрион, туберкулезная палочка).
Для аэробов, растущих на агаре или в тонких слоях жидкости в присутствии воздуха, кислорода вполне достаточно. В жидких средах при большом объеме жидкости аэробные бактерии могут расти только на поверхности, так как в более глубоких слоях по мере удаления от поверхности условия приближаются к анаэробным. Для нормального роста аэробных микроорганизмов в глубоких слоях жидкой культуры требуется аэрация.
Микроаэрофилы (факультативные аэробы) нуждаются в уменьшенной концентрации (низком парциальном давлении) свободного кислорода (от 0,01 до 0,03 бар). При полном доступе кислорода (в воздухе парциальное давление О2 составляет 0,2 бар) рост микроорганизмов прекращается или резко замедляется. Микроэрофилами является большинство аэробных бактерий.
Капнофилы нуждаются в наличии в среде двуокиси углерода (до 10 %), так как приспособлены к более высокому, чем в воздухе, содержанию СО2. Для этого кпитательным средам добавляют бикарбонат натрия и инкубируют культуры в закрытых сосудах в атмосфере, содержащей СО3 -2 ; можно также продувать обычный или обогащенный СО2 воздух. Капнофилами являются многие патогенные бактерии.
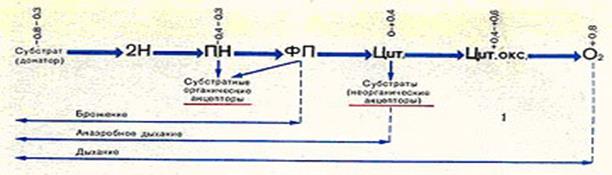
Рис. 58.Транспорт электронов при различных типах биологического окисления
ПН — пиридиннуклеотидных ферменты; ФП — флавопротеидные ферменты;
Цит. —цитохромные ферменты; Цит. окс. — цитохромоксидаза
2. Анаэробный(от греч. an — отрицание, аer — воздух и жизнь) тип—совокупность экзотермических процессов ферментативного распада органических веществ, происходящих без участия кислорода. Анаэробы получают энергию в форме АТФ путем ускоренного, но не полного окисления(субстратного фосфорилирования) углеводов, белков, липидов.

Различия между облигатными аэробами и анаэробами касаются ферментативного обеспечения конечных этапов окисления. У анаэробов дегидрогеназы не связаны с мембранами и находятся в ЦП только в растворимой форме. Для переноса водорода анаэробы используют флавиновые ферменты, самоокисляющиеся кислородом воздуха. Реакция флавиновых ферментов с кислородом направлена на детоксикацию молекулярного кислорода. Поэтому содержание флавиновых дегидрогеназ в клетках анаэробов значительно выше, чем у аэробов.
Из-за отсутствия ряда ферментов цепи переноса электронов (цитохромов) транспорт электронов осуществляется просто и примитивно. Органические субстраты окисляются только за счет отщепления водорода. Освобождающийся водород либо присоединяется к продуктам распада того же самого органического вещества, либо выделяется в газообразном состоянии. Если у анаэробов в качестве акцепторов водорода в оксилительно-восстановительных реакциях выступают неорганические вещества, происходит «анаэробное дыхание». Конечным акцептором водорода у анаэробов является связанный кислород в виде нитратов («нитратное дыхание») или сульфатов («сульфатное дыхание»).
Анаэробный путь филогенетически более ранний, чем аэробный, но он экономически невыгоден, так как выделяется небольшое количество энергии и образуется мало метаболитов. Так как энергии выделяется мало, микроорганизмам надо использовать большое количество субстратов (сахаров, аминокислот, пуринов и пиримидинов). Многие анаэробы являются строго протеолитическими организмами, неспособными сбраживать углеводы. Протеолитическая активность у некоторых анаэробов (C. histolyticum) настолько высока, при инфекциях, вызванных этими микроорганизмами, наблюдается расплавление мышц. Другие анаэробы обладают слабовыраженной сахаролитической активностью.
3. Если и донорами и акцепторами водорода выступают органические соединения, происходит брожение— процесс ферментативного расщепления органических соединений, преимущественно углеводов, с образованием спиртов (этилового, пропилового, глицерина) и органических кислот (молочной, муравьиной, уксусной, масляной). Типы брожения именуются по продуктам, которые образуются: молочнокислое, уксуснокислое, спиртовое, муравьинокислое, маслянокислое, ацетонобутиловое, пропионовокислое, сукцинатное, смешанное (рис. 59).
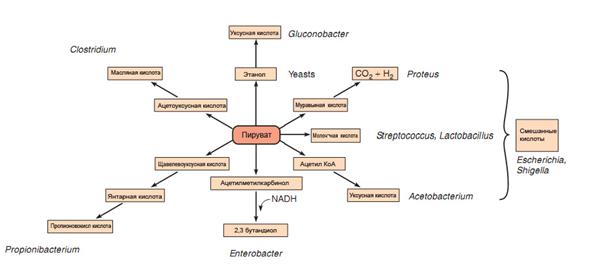
Рис. 59. Различные продукты ферментации пирувата
Образующиеся в процессе брожения продукты и энергия расходуются микроорганизмами для различных биосинтетических целей. Многие виды брожения используются в пищевой и микробиологической промышленности для получения спиртов и органических кислот.
Анаэробы осуществляют обмен веществ и размножаются в условиях отсутствия кислорода в среде обитания. Различают облигатных, аэротолерантных и факультативных анаэробов.
Облигатные (строгие) анаэробы (палочка маслянокислого брожения, возбудители столбняка, ботулизма) используют только анаэробный путьбиологического окисления и размножаются только в анаэробных условиях. Они лишены ферментов супероксиддисмутазы и каталазы,защищающих микробную клетку от токсических продуктов окисления. Поэтому вегетативные формы облигатных анаэробов погибают от ядовитых концентраций перекиси водорода даже при кратковременном контакте с воздухом. Спорам облигатных анаэробов свойствен крайне выраженный анабиоз, поэтому они могут сохраняться в присутствии кислорода.
Аэротолерантные микроорганизмы (молочнокислые бактерии, C. histolyticum) обладают супероксиддисмутазной активностью, но лишены каталазы и пероксидазы;они могут расти в присутствии атмосферного кислорода, но не способны его использовать; энергию получают исключительно с помощью брожения.
Факультативные анаэробы(большинство патогенных бактерий, например, Enterobacteriaceae (кишечная палочка, сальмонеллы), многие дрожжи) обладают супероксиддисмутазной и каталазной активностью.Факультативные анаэробы образуют АТФ при окислительном и субстратном фосфорилировании. Они могут изменять свой путь биологического окисления в зависимости от наличия или отсутствия кислорода. В кислородных условиях в качестве конечного акцептора водорода они используют атмосферный кислород. В бескислородных условиях в качестве конечного акцептора водорода они используют нитраты или сульфаты.
Классификация бактерий по отношению к кислороду воздуха представлена в табл. 22 и на рис. 60.
Классификация бактерий по отношению к кислороду воздуха
| Группа бактерий | Наличие ферментов, нейтрализующих токсические кислородные продукты | При доступе кислорода воздуха | Без доступа кислорода воздуха | ||
| Супероксид-дисмутаза (О • →Н2О2) | Каталаза (Н2О2→ Н2О+ О2) | ||||
| Облигатные аэробы | + | + | Растут | Не растут | |
| Микроаэрофилы | + | ± | ↓О2 – растут | ||
| Капнофилы | + | ± | ↑СО2 — растут | ||
| Анаэробы | Облигатные | – | – | Погибают | Растут |
| Аэротолерантные | + | – | Не растут, но не погибают | ||
| Факультативные | + | + | Растут |
Анаэробы очень широко распространены. Они являются возбудителями ряда опасных инфекционных заболеваний человека, используются в промышленности как продуценты ценных для народного хозяйства соединений.
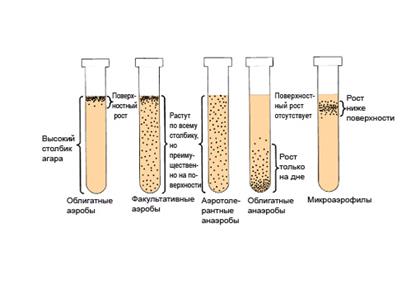
Рис. 60. Выявление типа дыхания микроорганизмов
при культивировании в высоком столбике агара
Источник



