
Сальтаторное проведение
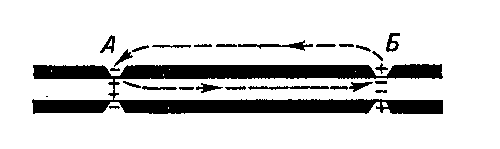
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
Смотреть что такое «Сальтаторное проведение» в других словарях:
САЛЬТАТОРНОЕ ПРОВЕДЕНИЕ — (лат. saltatorius, от salto скачу, прыгаю), скачкообразное проведение нервного импульса от одного перехвата Ранвье к другому вдоль мякотного (миелинизированного) аксона. Для С. п. характерно сочетание электротонич. распространения по… … Биологический энциклопедический словарь
Проведение нервного импульса — передача сигнала в виде волны возбуждения (См. Возбуждение) в пределах одного Нейрона и от одной клетки к другой. П. н. и. по нервным проводникам происходит с помощью электротонических потенциалов и потенциалов действия, которые… … Большая советская энциклопедия
проведение возбуждения сальтаторное — (лат. saltatorius танцевальный, плясовой) бездекрементное П. в. в мякотном нервном волокне скачками от одного перехвата Ранвье к другому … Большой медицинский словарь
Потенциал действия — Потенциал действия волна возбуждения, перемещающаяся по мембране живой клетки в процессе передачи нервного сигнала. По сути своей представляет электрический разряд быстрое кратковременное изменение потенциала на небольшом участке… … Википедия
Нервные волокна — отростки нервных клеток (Аксоны) вместе с их оболочками, проводящие нервные импульсы. Н. в. обычно имеют толщину 0,5 30 мкм; некоторые Н. в. у низших позвоночных и у беспозвоночных бывают значительно толще (у кольчатого червя Myxicola до… … Большая советская энциклопедия
Потенциал действия (action potential) — П. д. это самораспространяющаяся волна изменения мембранного потенциала, к рая последовательно проводится но аксону нейрона, перенося информ. от клеточного тела нейрона до самого конца его аксона. При нормальной передаче информ. в нервных сетях П … Психологическая энциклопедия
Не́рвы — (nervi) анатомические образования в виде тяжей, построенные преимущественно из нервных волокон и обеспечивающие связь центральной нервной системы с иннервируемыми органами, сосудами и кожным покровом тела. Нервы отходят парами (левый и правый) от … Медицинская энциклопедия
Источник
Сальтаторный способ передачи возбуждения
Миелинизированные и немиелинизированные нервные волокна. На рисунке показан поперечный срез типичного небольшого нерва, на котором видно много крупных нервных волокон, составляющих большую часть среза. Однако при более внимательном рассмотрении между крупными волокнами можно обнаружить множество очень мелких волокон. Крупные волокна миелинизированы, мелкие — немиелинизированы. В среднем нервный ствол содержит вдвое больше не-миелинизированных (безмякотных) нервных волокон, чем миелинизированных (мякотных).
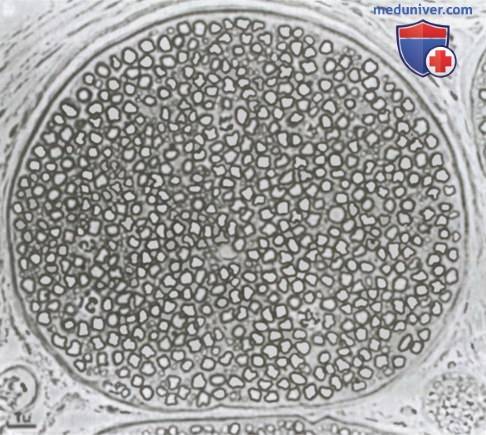 Поперечное сечение небольшого нервного ствола, содержащего миелинизированные и немиелинизированные волокна.
Поперечное сечение небольшого нервного ствола, содержащего миелинизированные и немиелинизированные волокна. 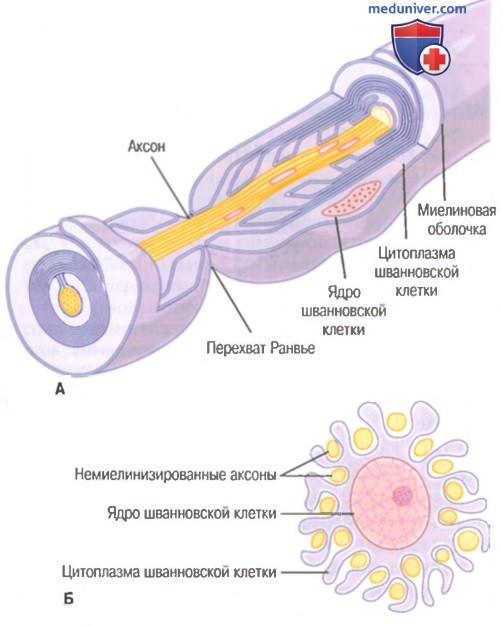 Значение шванновских клеток для изоляции нервных волокон.
Значение шванновских клеток для изоляции нервных волокон.
А. Накручивание мембраны шванновской клетки на крупный аксон формирует миелиновую оболочку миелинизированного нервного волокна.
Б. Частичная закрутка мембраны и цитоплазмы шванновской клетки вокруг множества немиелинизированных нервных волокон (показано в поперечном сечении).
На рисунке показано типичное миелинизированное волокно. Центральной его частью является аксон, по мембране которого проводится потенциал действия. Аксон заполнен аксоплазмой — вязкой внутриклеточной жидкостью. Аксон окружен миелиновой оболочкой, которая часто много толще, чем сам аксон. Примерно через каждые 1-3 мм вдоль миелиновой оболочки имеется перехват Ранвье.
Миелиновая оболочка формируется вокруг аксона шванновскими клетками. Мембрана шванновской клетки сначала охватывает аксон, затем шванновская клетка многократно вращается вокруг аксона, образуя многочисленные мембранные слои, содержащие липидное вещество сфингомиелин. Это вещество является отличным изолятором и снижает ионный ток через мембрану аксона примерно в 5000 раз. Между каждыми двумя последовательно расположенными шванновскими клетками по ходу аксона остается маленькая неизолированная область длиной всего 2-3 мкм, где ионы могут свободно переходить через мембрану аксона из внеклеточной жидкости во внутриклеточную и обратно. Эту область называют перехватом Ранвье.
Сальтаторное проведение в миелиновых волокнах от перехвата к перехвату. Ионы практически не могут проходить через толстую миелиновую оболочку мякотных волокон, однако они легко диффундируют через перехваты Ранвье. Следовательно, потенциалы действия возникают только в перехватах и проводятся от перехвата к перехвату; это называют салътаторным (скачкообразным) проведением. В этом случае электрический ток течет через внеклеточную жидкость снаружи от миелиновой оболочки, а также через аксоплазму внутри аксона от перехвата к перехвату, последовательно возбуждая один перехват за другим. Таким образом, нервный импульс как будто прыгает по волокну, на основании этого и появился термин «салътаторное проведение».
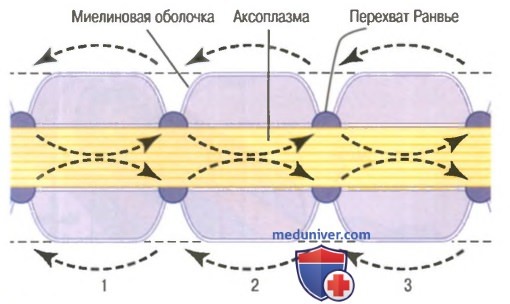 Сальтаторное проведение по миелинизированному аксону. Стрелки указывают направление движения электрического тока от перехвата к перехвату.
Сальтаторное проведение по миелинизированному аксону. Стрелки указывают направление движения электрического тока от перехвата к перехвату.
Сальтаторное проведение имеет два преимущества. Во-первых, заставляя процесс деполяризации «прыгать» через большие промежутки вдоль аксона, этот механизм повышает скорость проведения в миелинизированных волокнах в 5-50 раз. Во-вторых, сальтаторное проведение сохраняет энергию для аксона, поскольку деполяризуются только перехваты, что позволяет приблизительно в 100 раз снизить потерю ионов по сравнению с возможными потерями в других условиях. В связи с этим снижаются траты энергии, необходимые для восстановления трансмембранной разности концентраций ионов натрия и калия после серии нервных импульсов.
Существует другая особенность сальтаторного проведения в крупных миелинизированных волокнах: отличная изоляция, обеспечиваемая миелиновой оболочкой, и 50-кратное снижение мембранной емкости позволяют осуществлять реполяризацию путем перемещения очень незначительного числа ионов.
Скорость проведения в нервных волокнах. Скорость проведения в нервных волокнах колеблется от 0,25 м/сек в очень тонких немиелинизирован-ных волокнах до 100 м (длина футбольного поля) в 1 сек в очень толстых миелинизированных волокнах.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
6.3.2. Сальтаторное проведение
Второй способ увеличить скорость проведения нервных импульсов, реализовавшийся только у позвоночных животных, состоит в изоляции участков аксона с помощью миелиновой оболочки. При этом постоянная длины соответствующих участков значительно увеличивается, и тем самым существенно облегчается проведение тока в продольном направлении. По мере развития животного миелин откладывается вокруг периферических и центральных аксонов глиальными клетками, расположенными вблизи этих аксонов. В результате вокруг волокон образуется плотная многослойная оболочка из клеточных мембран. К клеткам, синтезирующим миелин, относятся шванновские клетки (в области периферических нервов) и олигодендроциты (рис. 6–10) (в ЦНС). На поперечных срезах миелиновой оболочки видны периодически повторяющиеся промежутки в 12 нм, образующиеся в результате наслоения мембран глиальных клеток. С образованием каждого нового слоя поперечное сопротивление оболочки увеличивается. Поскольку слоев в этой оболочке много, ее емкость гораздо ниже, чем у одиночной мембраны. Многослойная миелиновая оболочка периодически прерывается (так называемые перехваты Ранвье>, и на этих небольших участках возбудимая мембрана аксона контактирует с внеклеточной средой. Между перехватами Ранвье миелиновая оболочка тесно прилегает к мембране аксона, практически вытесняя внеклеточную среду. Кроме того, участки мембраны аксона между перехватами Ранвье, по–видимому, не содержат натриевых каналов.

Рис. 6.10. Перехват Ранвье. Показан короткий «голый» участок аксона, расположенный между двумя миелинизированными участками. Именно этот участок возбуждается при сальтаторном проведении. На рис. 4–12 приведена электронная микрофотография, на которой видна многослойная миелиновая оболочка, образуемая мембранами глиальных клеток. (Bunge et al., 1961.)
Благодаря изолирующим свойствам миелиновой оболочки постоянная длины аксона резко возрастает: наличие этой оболочки оказывает такой же эффект, как и увеличение rм [уравнение (6–2)]. Из–за высокого сопротивления миелиновой оболочки местные токи, текущие впереди от волны возбуждения, выходят из аксона почти исключительно в области перехватов Ранвье. Кроме того, поскольку емкость толстой миелиновой оболочки мала, на перезарядку этой емкости в участках между перехватами расходуется лишь очень небольшой ток. Благодаря этим особенностям ПД, возникающий в каком–либо перехвате, электротонически деполяризует лишь мембрану, расположенную в области следующего перехвата, и поэтому импульсы в таких аксонах не распространяются по всей их длине, как в немиелинизированных нервных волокнах (например, в аксоне кальмара). Они возникают лишь в небольших участках мембраны –перехватах Ранвье. Все это обусловливает сальтаторное (скачкообразное) проведение, при котором импульсы распространяются прерывисто от перехвата к перехвату (рис. 6–11). Скорость распространения. ПД при этом резко увеличивается, поскольку электротоническое проведение местных токов между перехватами осуществляется очень быстро. Таким образом, у позвоночных животных Природа решила проблему быстрого распространения нервных импульсов, не прибегая к созданию таких громоздких структур, как гигантские аксоны.

Рис. 6.11. Сальтаторное проведение в миелинизированном аксоне. Приведены кривые тока, как бы «зафиксированные» в определенный момент времени. А. ПД возникает только в перехватах Ранвье и перескакивает от одного перехвата к другому. Местный ток распространяется между перехватами в продольном направлении. Крупными стрелками изображен вход Na + через активированные натриевые каналы в области перехватов. Б. Внутриклеточный потенциал в каждом перехвате в момент времени (кружки), представленный в верхней части рисунка.
Источник
6.3.2. Сальтаторное проведение
Второй способ увеличить скорость проведения нервных импульсов, реализовавшийся только у позвоночных животных, состоит в изоляции участков аксона с помощью миелиновой оболочки. При этом постоянная длины соответствующих участков значительно увеличивается, и тем самым существенно облегчается проведение тока в продольном направлении. По мере развития животного миелин откладывается вокруг периферических и центральных аксонов глиальными клетками, расположенными вблизи этих аксонов. В результате вокруг волокон образуется плотная многослойная оболочка из клеточных мембран. К клеткам, синтезирующим миелин, относятся шванновские клетки (в области периферических нервов) и олигодендроциты (рис. 6–10) (в ЦНС). На поперечных срезах миелиновой оболочки видны периодически повторяющиеся промежутки в 12 нм, образующиеся в результате наслоения мембран глиальных клеток. С образованием каждого нового слоя поперечное сопротивление оболочки увеличивается. Поскольку слоев в этой оболочке много, ее емкость гораздо ниже, чем у одиночной мембраны. Многослойная миелиновая оболочка периодически прерывается (так называемые перехваты Ранвье>, и на этих небольших участках возбудимая мембрана аксона контактирует с внеклеточной средой. Между перехватами Ранвье миелиновая оболочка тесно прилегает к мембране аксона, практически вытесняя внеклеточную среду. Кроме того, участки мембраны аксона между перехватами Ранвье, по–видимому, не содержат натриевых каналов.
Рис. 6.10. Перехват Ранвье. Показан короткий «голый» участок аксона, расположенный между двумя миелинизированными участками. Именно этот участок возбуждается при сальтаторном проведении. На рис. 4–12 приведена электронная микрофотография, на которой видна многослойная миелиновая оболочка, образуемая мембранами глиальных клеток. (Bunge et al., 1961.)
Благодаря изолирующим свойствам миелиновой оболочки постоянная длины аксона резко возрастает: наличие этой оболочки оказывает такой же эффект, как и увеличение rм [уравнение (6–2)]. Из–за высокого сопротивления миелиновой оболочки местные токи, текущие впереди от волны возбуждения, выходят из аксона почти исключительно в области перехватов Ранвье. Кроме того, поскольку емкость толстой миелиновой оболочки мала, на перезарядку этой емкости в участках между перехватами расходуется лишь очень небольшой ток. Благодаря этим особенностям ПД, возникающий в каком–либо перехвате, электротонически деполяризует лишь мембрану, расположенную в области следующего перехвата, и поэтому импульсы в таких аксонах не распространяются по всей их длине, как в немиелинизированных нервных волокнах (например, в аксоне кальмара). Они возникают лишь в небольших участках мембраны –перехватах Ранвье. Все это обусловливает сальтаторное (скачкообразное) проведение, при котором импульсы распространяются прерывисто от перехвата к перехвату (рис. 6–11). Скорость распространения. ПД при этом резко увеличивается, поскольку электротоническое проведение местных токов между перехватами осуществляется очень быстро. Таким образом, у позвоночных животных Природа решила проблему быстрого распространения нервных импульсов, не прибегая к созданию таких громоздких структур, как гигантские аксоны.
Рис. 6.11. Сальтаторное проведение в миелинизированном аксоне. Приведены кривые тока, как бы «зафиксированные» в определенный момент времени. А. ПД возникает только в перехватах Ранвье и перескакивает от одного перехвата к другому. Местный ток распространяется между перехватами в продольном направлении. Крупными стрелками изображен вход Na + через активированные натриевые каналы в области перехватов. Б. Внутриклеточный потенциал в каждом перехвате в момент времени (кружки), представленный в верхней части рисунка.
Источник



