
Пурпурные бактерии




![]()
![]()
Группа пурпурных бактерий, насчитывающая более 50 видов, представлена одноклеточными организмами разной морфологии (рис. 78). Длина их колеблется от 1 до 20 мкм, ширина — от 0,3 до 6 мкм. Некоторые виды образуют выросты. Среди пурпурных бактерий есть неподвижные и подвижные формы.
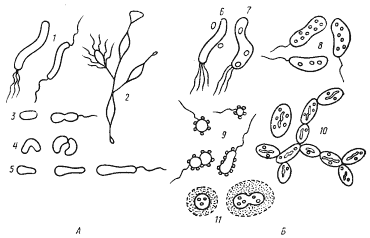 |
| Рис. 78. Основные морфологические типы пурпурных бактерий: А — несерные пурпурные бактерии: 1 — Rhodospirillum; 2 — Rhodomicrobium; 3 — Rhodobacter sphaeroides; 4 — Rhodocyclus; 5 — Rhodopseudomonas palustris; Б — пурпурные серобактерии: 6 — Ectothiorhodospira; 7 — Thiospirillum; 8 — Chromatium; 9 — Thiocystis; 10 — Thiodictyon; 11 — Thiocapsa (по Горленко, Дубининой, Кузнецову, 1977) |
Движение осуществляется с помощью одного или пучка жгутиков, расположенных обычно полярно. Большинство пурпурных бактерий размножаются бинарным делением, некоторые виды — почкованием. Клетки неподвижных форм, размножающихся поперечным делением в разных плоскостях, имеют тенденцию формировать агрегаты правильной геометрической формы.
Все пурпурные бактерии окрашиваются отрицательно по Граму и, следовательно, имеют сложное строение клеточной стенки. Для клеток характерна хорошо развитая система внутрицитоплазматических фотосинтетических мембран, являющихся производными ЦПМ и сохранивших с ней отчетливо наблюдаемую связь. Мембраны имеют вид отдельных пузырьков, трубок или пластинок (ламелл), располагающихся по периферии клетки (см. рис. 4), и представляют вместе с ЦПМ единую систему. Подобно многим обитающим в толще воды эубактериям в клетках некоторых неподвижных пурпурных бактерий содержатся газовые вакуоли. В качестве запасных веществ обнаружены углевод типа гликогена и поли--оксимасляная кислота. Группа пурпурных бактерий довольно гетерогенна в отношении нуклеотидного состава ДНК. Молярное содержание ГЦ-оснований колеблется от 45 до 73%, хотя у большинства представителей оно находится в пределах 61 — 73%.
Все пурпурные бактерии характеризуются сходным строением и функционированием фотосинтетического аппарата. Они могут расти на свету в анаэробных условиях, осуществляя фотосинтез бескислородного типа. Однако по целому ряду физиологических особенностей, в том числе и по использованию разных соединений в качестве донора электронов при фотосинтезе, между представителями пурпурных бактерий обнаружены значительные различия. Поэтому на основании ряда физиологических признаков группу подразделяют на пурпурные серные и несерные бактерии.
Для пурпурных серобактерий основной способ существования — фотолитоавтотрофия. Все представители этой группы могут расти при освещении в анаэробных условиях на среде с CO2 в качестве единственного источника углерода, используя как донор электронов сульфид (H2S). Многие виды могут использовать для этой цели молекулярную серу (S 0 ), сульфит (SO3 2 – ), тиосульфат (S2O3 2– ), молекулярный водород. Сульфид окисляется последовательно до молекулярной серы или сульфата, при этом глобулы серы откладываются в периплазматическом пространстве и впячиваниях (инвагинатах) ЦПМ, которые также являются частью этого пространства. Исключение составляют виды рода Ectothiorhodospira, окисляющие сульфид и тиосульфат до молекулярной серы, но не накапливающие последнюю в пределах клетки. Представители этого рода выделяют серу в среду, а затем поглощают ее и окисляют до SO4 2 – . Ферменты, катализирующие окисление восстановленных соединений серы, локализованы в периплазматическом пространстве и на наружной поверхности ЦПМ.
Пурпурные серобактерии обнаруживают весьма ограниченную способность использовать органические соединения. В большинстве случаев последние служат дополнительными источниками углерода и редко — донорами электронов. Все виды могут фотометаболизировать ацетат и пируват, только некоторые способны существовать полностью фотоорганогетеротрофно.
В течение длительного времени пурпурные серобактерии считали строгими анаэробами и облигатными фототрофами. Недавно было показано, что спектр отношения к молекулярному кислороду в этой группе достаточно широк. В большинстве пурпурные серобактерии высоко чувствительны к O2, однако и среди них есть виды, растущие в темноте в аэробных условиях на минеральной среде или с использованием органических соединений. Хемолитоавтотрофный рост при низком содержании O2 обнаружен у ряда пурпурных серобактерий, ассимилирующих CO2 в восстановительном пентозофосфатном цикле, а энергию получающих в процессе дыхания в результате окисления сульфида, тиосульфата, молекулярной серы или H2. Для некоторых представителей родов Ectothiorhodospira и Thiocapsa показан аэробный хемоорганогетеротрофный рост. Органические вещества в этом случае используются как источники углерода и энергии. Последняя запасается в процессе дыхания. Некоторые пурпурные серобактерии оказались также способными расти в темноте в анаэробных условиях (в атмосфере аргона или молекулярного водорода), сбраживая некоторые сахара или органические кислоты.

Пурпурные несерные бактерии имеют склонность к фотоорганогетеротрофному образу жизни, предпочитая в качестве доноров электронов и источников углерода в процессе фотосинтеза простые органические соединения: жирные кислоты, спирты, сахара, аминокислоты. Многие виды способны расти фотолитоавтотрофно, используя молекулярный водород в качестве донора электронов для восстановления CO2.
Некоторые типичные несерные пурпурные бактерии растут при освещении на минеральной среде, используя в качестве донора электронов H2S, тиосульфат или молекулярную серу. В большинстве случаев сульфид окисляется только до молекулярной серы, никогда не откладывающейся в клетке, но в отдельных случаях возможно последующее окисление S 0 до сульфата.
В группе несерных бактерий обнаружено большое разнообразие метаболических путей, связанных с получением энергии. Многие представители этой группы способны расти в темноте в микроаэробных или аэробных условиях, получая энергию в процессе дыхания. У них активно функционирует замкнутый ЦТК, гликолитический путь и другие пути катаболизма органических соединений.
Представители рода Rhodobacter способны к хемоавтотрофии. Они растут на минеральной среде в темноте при пониженной концентрации O2, используя энергию, получаемую при окислении молекулярного водорода, для ассимиляции CO2.
У несерных пурпурных бактерий развиты контакты с молекулярным кислородом. У них имеются ферментные системы защиты от O2. Все несерные пурпурные бактерии способны расти хемотрофно в микроаэробных условиях, хотя не все из них могут переносить атмосферное содержание O2. При концентрации O2 от 0,5 до 5% фотосинтез и окислительный метаболизм могут функционировать одновременно. Молекулярный кислород у несерных пурпурных бактерий (как и у всех эубактерий, осуществляющих бескислородный фотосинтез) выступает как мощный фактор, регулирующий их метаболизм. Уже в достаточно низких концентрациях O2 ингибируют синтез бактериохлорофиллов, внутрицитоплазматических мембран и рибулозодифосфаткарбоксилазы. В то же время в присутствии O2 наблюдается увеличение активности ферментов ЦТК.
Среди представителей рода Rhodobacter обнаружена способность расти в анаэробных условиях за счет окисления органических соединений, сопряженного с транспортом электронов на нитраты (анаэробное дыхание). Наконец, в последние годы для ряда несерных пурпурных бактерий показана способность расти анаэробно в темноте, осуществляя сбраживание органических субстратов, таких как сахара, пируват.
Таким образом, в этой группе обнаружены разные биосинтетические способности, сочетающиеся с разнообразными способами получения энергии. Источниками углерода могут быть CO2 или органические соединения, а источниками энергии — фотосинтез, аэробное и анаэробное дыхание, брожение.
Основные физиолого-биохимические различия между серными и несерными пурпурными бактериями представлены в табл. 25. До настоящего времени не найдено четкого признака или нескольких признаков, которые могли бы быть положены в основу деления пурпурных бактерий на рассмотренные физиологические группы. Как правило, признак, характерный для организмов одной группы, можно найти у представителей другой. Тем не менее можно выявить совокупность свойств, типичных либо для серных, либо для несерных пурпурных бактерий.
Недавно обнаружены эубактерий, выделенные в новый род Erythrobacter, образующие подобно многим пурпурным бактериям бактериохлорофилл a, но растущие только в аэробных условиях. Это грамотрицательные палочки, перемещающиеся с помощью жгутиков; размножаются бинарным делением. Молярное содержание ГЦ-оснований в ДНК — 60 — 64%. Система внутрицитоплазматических мембран везикулярного типа сходна с таковой несерных пурпурных бактерий. Помимо бактериохлорофилла a эритробактеры содержат в значительном количестве каротиноиды, определяющие их розовую или оранжевую окраску.
Таблица 25. Основные физиолого-биохимические различия между серными и несерными пурпурными бактериями
| Признак | Пурпурные бактерии | |
| несерные | серные | |
| Доноры электронов при фотосинтезе | ||
| H2S |  * * |  |
| S 0 | |  |
| H2 | | |
| Источник углерода | ||
| CO2 | | |
| Органические соединения | | ** |
| Рост в темноте на органических средах за счет | ||
| аэробного дыхания | | * |
| анаэробного дыхания | |  |
| брожения | | |
| Способность к хемоавтотрофии | | |
| Отношение к O2 воздуха | факультативные анаэробы | в большинстве — строгие анаэробы; отдельные виды — факультативные анаэробы |
| Способность к азотфиксации | | |
* Признак обнаружен у всех ( ), большинства ( ), некоторых ( ) представителей группы; ( ) признак отсутствует.
** Количество фотоассимилируемых органических соединений и степень их использования невелики.
В отличие от пурпурных бактерий выделенные до сих пор представители рода Erythrobacter — облигатно аэробные хемоорганогетеротрофы: не могут расти анаэробно ни на свету 59 , ни в темноте; не растут на свету в аэробных условиях за счет неорганических субстратов, но хорошо растут в тех же условиях в присутствии разнообразных органических соединений (сахаров, органических кислот, некоторых одноуглеродных соединений). Многие штаммы нуждаются в витаминах.
59 Описан штамм Erythrobacter, способный расти на свету в анаэробных условиях, если молекулярный кислород заменить нитратами в качестве конечного акцептора электронов.
В то же время получены экспериментальные доказательства использования эритробактерами энергии света: установлено обратимое фотоокисление бактериохлорофилла a реакционного центра, показано светозависимое включение CO2 и повышение уровня АТФ в клетке; установлена способность мембранных препаратов к фотофосфорилированию. Однако фотосинтетический аппарат, имеющийся в клетках Erythrobacter, не может обеспечить их рост. Облигатная зависимость от молекулярного кислорода связана с тем, что для эритробактеров основным источником энергии служит O2-зависимое дыхание. Фотосинтетическая активность может иметь значение для поддержания жизнеспособности клеток в отсутствие в среде субстратов, обеспечивающих рост.
Бактерии рода Erythrobacter интересны тем, что необходимую для роста энергию получают в результате аэробного дыхания, но не утратили при этом бактериохлорофилла a и других компонентов фотосинтетического аппарата.
Источник
Пурпурные бактерии — описание, особенности и интересные факты
Что такое пурпурные бактерии? Эти микроорганизмы пигментированы бактериохлорофиллом a или b вместе с различными каротиноидами, которые придают им цвета в диапазоне от фиолетового, красного, коричневого и оранжевого. Это довольно разнообразная группа. Их можно разделить на две группы: пурпурные серные бактерии и простые фиолетовые бактерии (Rhodospirillaceae). В документе 2018 Frontiers in Energy Research было предложено использовать их в качестве биоресурсов.
Биология
 Вам будет интересно: Площадь боковой поверхности и объем усеченной пирамиды: формулы и пример решения типовой задачи
Вам будет интересно: Площадь боковой поверхности и объем усеченной пирамиды: формулы и пример решения типовой задачи
Пурпурные бактерии в основном являются фотоавтотрофными, но также известны химиоавтотрофные и фотогетеротропные виды. Они могут быть миксотрофами, способными к аэробному дыханию и ферментации.
Фотосинтез пурпурных бактерий происходит в реакционных центрах на клеточной мембране, где фотосинтетические пигменты (то есть бактериохлорофилл, каротиноиды) и связывающие пигмент белки вводятся в инвагинацию с образованием особых пузырьков, трубочек или однопарных или штабелированных пластинчатых листов. Это называется внутрицитоплазматической мембраной (ICM), которая имеет увеличенную площадь поверхности, чтобы максимизировать поглощение света.
Физика и химия
 Вам будет интересно: Профиль крыла самолета: виды, технические и аэродинамические характеристики, метод расчета и наибольшая подъемная сила
Вам будет интересно: Профиль крыла самолета: виды, технические и аэродинамические характеристики, метод расчета и наибольшая подъемная сила
Пурпурные бактерии используют циклический перенос электронов, вызванный серией окислительно-восстановительных реакций. Легкоуборочные комплексы, окружающие реакционный центр (RC), собирают фотоны в виде резонансной энергии, захватывающие пигменты хлорофилла P870 или P960, расположенные в RC. Возбужденные электроны циклируются от P870 до хинонов QA и QB, затем переходят в цитохром bc1, цитохром c2 и обратно в P870. Восстановленный хинон QB притягивает два цитоплазматических протона и становится QH2, в конечном счете окисляясь и высвобождая протоны, подлежащие закачке в периплазму комплексом цитохрома bc1. Результирующее разделение заряда между цитоплазмой и периплазмой создает протонную движущую силу, используемую АТФ-синтазой для получения энергии АТФ.
 Вам будет интересно: Единицы измерения объема информации
Вам будет интересно: Единицы измерения объема информации
Пурпурные бактерии также переносят электроны от внешних доноров непосредственно на цитохром bc1, чтобы генерировать NADH или NADPH, используемые для анаболизма. Они являются однокристами, потому что они не используют воду в качестве донора электронов для получения кислорода. Один тип фиолетовых бактерий, называемых фиолетовыми серными бактериями (PSB), использует сульфид или серу в качестве доноров электронов. Другой тип, называемый фиолетовыми несернистыми бактериями, обычно использует водород в качестве донора электронов, но также может использовать сульфидные или органические соединения при более низких концентрациях по сравнению с PSB.
Фиолетовым бактериям не хватает внешних носителей электронов, чтобы спонтанно уменьшить NAD (P) + до NAD (P) H, поэтому они должны использовать их уменьшенные хиноны для энангорного уменьшения NAD (P) +. Этот процесс обусловлен движущей силой протона и называется обратным потоком электронов.
Сера вместо кислорода
Пурпурные несерные бактерии были первыми бактериями, у которых был обнаружен фотосинтез без побочного продукта в виде кислорода. Вместо этого их побочным продуктом является сера. Это было доказано, когда впервые были установлены реакции бактерий на различные концентрации кислорода. Обнаружено, что бактерии быстро отходят от малейшего следа кислорода. Затем провели эксперимент где использовалось блюдо с бактериями, и свет был сфокусирован на одной его части, а другая оставлена в затемнении. Поскольку бактерии не могут выжить без света, они перемещаются в круг света. Если бы побочный продукт их жизнедеятельности был кислородом, расстояния между отдельными особями становилось бы все больше по мере увеличения количества кислорода. Но из-за поведения пурпурных и зеленых бактерий в сфокусированном свете было сделано заключение, что побочный продукт фотосинтеза бактерий не может быть кислородом.
Исследователи предположили, что некоторые фиолетовые бактерии сегодня связаны с митохондриями, симбиотическими бактериями в клетках растений и животных, которые действуют как органеллы. Сравнение их белкового строения показывает, что существует общий предок этих структур. Пурпурные зеленые бактерии и гелиобактерии также имеют схожее строение.
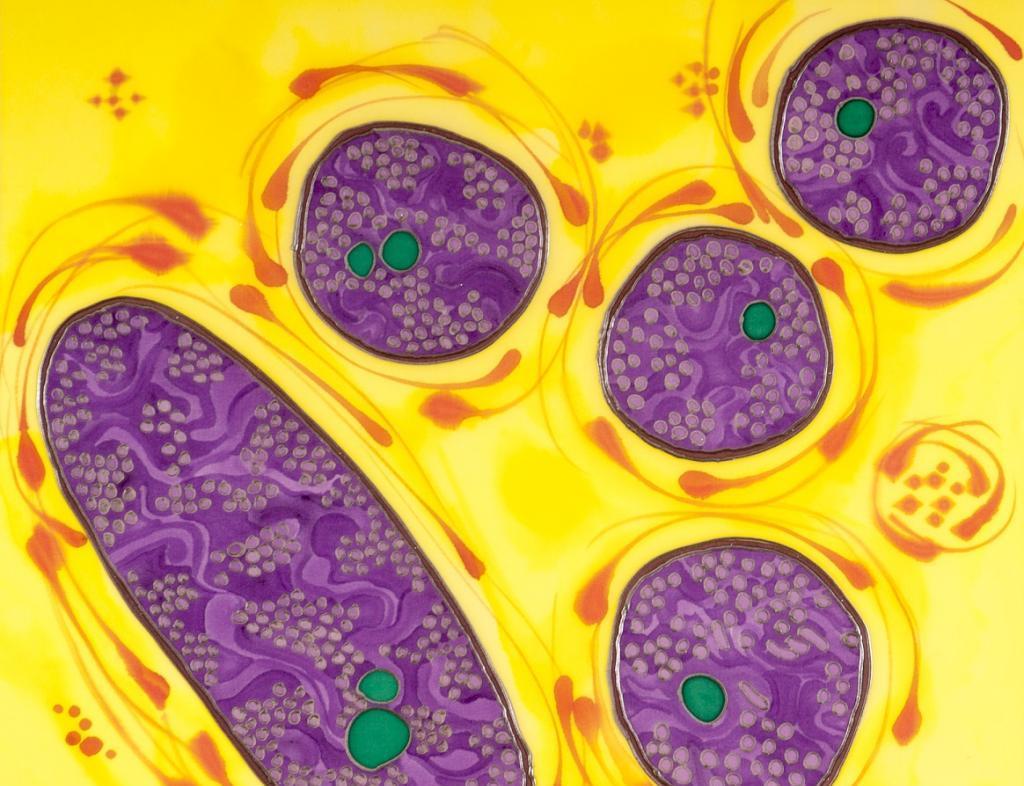
Серобактерии (серные бактерии)
Фиолетовые серные бактерии (PSB) являются частью группы Proteobacteria, способной к фотосинтезу, в совокупности, называемой пурпурными бактериями. Они являются анаэробными или микроаэрофильными и часто встречаются в стратифицированных водных средах, включая горячие источники, застойные водоемы, а также микробные скопления в зонах прилива. В отличие от растений, водорослей и цианобактерий, пурпурные серные бактерии не используют воду в качестве восстанавливающего агента и поэтому не производят кислород. Вместо этого они могут использовать серу в форме сульфида или тиосульфата (а также некоторые виды могут использовать H2, Fe2 + или NO2-) в качестве донора электронов на своих путях фотосинтеза. Сера окисляется для получения гранул элементарной серы. Она, в свою очередь, может быть окислена с образованием серной кислоты.
Классификация
Группа пурпурных бактерий делится на два семейства: Chromatiaceae и Ectothiorhodospiraceae, которые производят внутренние и внешние гранулы серы соответственно и показывают различия в структуре их внутренних мембран. Они составляют часть порядка Chromatiales, включенного в гамма-подразделение Proteobacteria. Род Halothiobacillus также включен в Chromatiales в собственной семье, но он не является фотосинтетическим.
Места обитания
Фиолетовые серные бактерии, как правило, обнаруживаются в освещенных аноксических зонах озер и других водных ареалах, где накапливается сероводород, а также в «серных источниках», где геохимически или биологически производимый сероводород может вызвать образование цветков фиолетовых серобактерий. Для фотосинтеза необходимы аноксические условия; эти бактерии не могут процветать в кислородосодержащих средах.

Наиболее благоприятными для развития фиолетовых серобактерий являются меромиктические (постоянно стратифицированные) озера. Они стратифицируют, потому что имеют более плотную (обычно физиологическую) воду внизу и менее плотную (обычно пресную воду) ближе к поверхности. Рост фиолетовых серобактерий также поддерживается расслоением в голомиктических озерах. Они термически стратифицированы: в весеннее и летнее время вода на поверхности нагревается, делая верхнюю воду менее плотной, чем нижняя, которая обеспечивает довольно стабильную стратификацию для роста пурпурных серных бактерий. Если присутствует достаточное количество сульфата для поддержки сульфатирования, сульфид, образующийся в отложениях, диффундирует вверх в бескислородные придонные воды, где фиолетовые серные бактерии могут образовывать плотные клеточные массы.

Скопления
Фиолетовые серные бактерии также могут быть найдены и являются заметным компонентом в промежуточных микробных скоплениях. Скопления такие, как микробный коврик Sippewissett, имеют динамическую среду из-за потока приливов и поступающей свежей воды, приводящей к аналогичным стратифицированным средам как меромиктические озера. Рост фиолетовых серобактерий активируется, поскольку сера поставляется из-за смерти и разложения микроорганизмов, расположенных над ними. Стратификация и источник серы позволяют PSB расти в этих приливных бассейнах, где возникают скопления. PSB может помочь стабилизировать осадки микробного скопления с помощью секреции внеклеточных полимерных веществ, которые могут связывать осадки в бассейнах водоемов.
Экология
Фиолетовые серные бактерии способны влиять на окружающую среду, способствуя циклированию питательных веществ, используя свой метаболизм для изменения окружающей среды. Они могут играть значительную роль в первичной продукции, влияя на углеродный цикл посредством фиксации углерода. Фиолетовые серные бактерии также способствуют производству фосфора в своей среде обитания. Через жизнедеятельность этих организмов фосфор, ограничивающий питательное вещество в оксическом слое озер, рециркулируется и предоставляется гетеротрофным бактериям для использования. Это указывает на то, что хотя фиолетовые серные бактерии находятся в аноксическом слое своей среды обитания, они способны стимулировать рост многих гетеротрофных организмов, поставляя неорганические питательные вещества в вышеупомянутый оксидный слой.
Источник



